
1. INTRODUCTION
Lycopene is a carotenoid compound (a lipophilic hydrocarbon pigment) that induces red, pink, and orange coloring in plants. It is synthesized in the chloroplast from phytoene and accumulates in its trans- form differentially in fruits and vegetables, mainly in tomatoes and chili peppers [1-4], but its content is affected by variety, crop maturity, plant–environment interaction [5], and agricultural plant management [6-11]. Thus, crops with high levels of this carotenoid represent a window of opportunity in the agri-food sector due to their impact on human health.
Crops rich in lycopene can be easily included in the diet and, thanks to their absorption [12], can improve endothelial function [13,14], minimize damage to immune cells [15], regulate the endocrine system, participate in the prevention of certain types of cancer [16], protect the gastrointestinal tract [17], induce neuroprotective effects [18], mitigate damage to the respiratory tract [19], and reduce cardiovascular disease [13]. These benefits are due to the inhibition of inflammatory mediators [20], removal and neutralization of reactive oxygen species (ROS) and reactive nitrogen species (RNS) [21], reduction of lipid peroxidation [22], and differential regulation of antioxidant genes [23]. These responses are conditioned both by intake and by individual metabolism. The average daily intake of lycopene ranges from 0.1 to 146.3 mg per day [24] depending on the bioavailability of the raw material [25]. However, studies show inconsistencies depending on sex, age, alcohol and tobacco consumption, food matrix, interaction with other biomolecules, and source processing [26-28].
Likewise, lycopene plays an important role in the industrial sector, as it is used as a food additive (coloring agent) in meat products [29], animal feed, cheese [30], yogurt [31], and in the bakery sector [32], where it minimizes oxidative damage and improves texture and flavor, enhancing the final presentation of the product [33]. This carotenoid is also marketed as a dietary supplement (capsules and tablets) and, thanks to its potential to neutralize free radicals, is used in the development of serums, creams, and sunscreens [34,35]. However, in this case, the evidence is not conclusive, and further studies and trials are needed to determine the permissible doses in humans to obtain adequate results.
Against this backdrop, various studies have focused on promoting lycopene yield in crops, improving the quality of extraction during processing, and investigating its impact on human health to identify any inconsistencies associated with this carotenoid. Considering the potential of lycopene in the agricultural sector, growing industrial demand, and its high importance in human health, this article aims to present an analysis of its biosynthesis, potential sources, content response to environmental conditions, strategies to promote content, effects on human health, and industrial applications. This information may be a point that, by showing the association between biosynthesis → environmental modulation → bioaccessibility → health outcomes → industrial uses, allows for the implementation of comprehensive studies linking the agricultural sector with the health and industrial sectors for the optimal and efficient use of lycopene.
2. LYCOPENE: STRUCTURE AND PROPERTIES
2.1. Chemical Structure
Lycopene (ψ,ψ-carotene) is a carotenoid characterized by an acyclic (possessing no closed chain) and unsaturated structure, without the presence of a β-ionone ring [36]. It is an isoprene-derived compound with the molecular formula C40H56 (89.45% carbon and 10.51% hydrogen) and a molecular weight of 536.85 g/mol, with a total of 13 carbon-carbon double bonds, equivalent to 11 conjugated and 2 unconjugated [37]. This system forms a cloud of π electrons extending along the polyene chain, which facilitates light absorption in the visible region with peaks at 444, 470, and 502 nm [3]. Similarly, the arrangement of double bonds along the chain gives lycopene the biological functionality of absorbing light in the visible range and acting as a free radical scavenger [38]. In addition, the chemical structure of lycopene is vitally important to health as it is associated with its potential to mitigate radicals, thus preventing oxidative damage in the body, while in the industrial sector, the structure provides the versatility to be used in various processes.
2.2. Properties
Lycopene has antioxidant capacity, captures or scavenges free radicals, and quenches singlet oxygen, which reduces damage caused by oxidative stress at the cellular level. From this, lies the importance of lycopene, given that several investigations have linked oxidative stress as a phenomenon that contributes to the pathogenesis and pathophysiology of many chronic health problems. The antioxidant activity of lycopene comes from its electron-rich environment, which allows it to absorb long wavelengths and neutralize singlet oxygen. According to several studies, these properties give it a greater capacity to eliminate free radicals: twice that of β-carotene and ten times that of α-tocopherol [Figure 1] [12,28,39]. However, this molecule exhibits extremely high lipophilicity (log P, partition coefficient = 15.09–17.64) [37], which limits its potential pharmacological applications. In addition to its low solubility, this molecule is unstable when exposed to light and heat, which poses a fundamental challenge as it can lead to significant degradation during processing. These limitations result in reduced lycopene yield and quality. In this regard, new studies should be conducted to minimize these losses and maximize their benefits in the food, cosmetics, and industrial sectors.
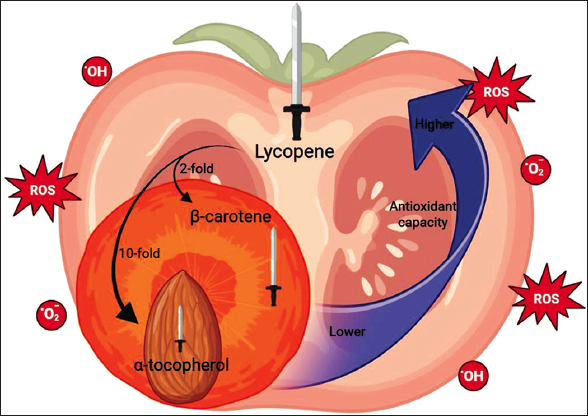 | Figure 1: Potential antioxidant capacity of lycopene. Figure our own, created using Microsoft Office PowerPoint (Microsoft Corporation, Redmond, Washington, USA). [Click here to view] |
2.3. Lycopene Isomers
Of the 2048 theoretical geometric configurations of lycopene, only 72 cis isomers (5-cis, 9-cis, 13-cis, and 15-cis) show favorable structural stability [Figure 2] [12,28]. It should be noted that during food processing, the use of organic solvents, heat treatments, and catalysts causes lycopene to undergo geometric isomerization, increasing cis-isomers. However, during food storage, this molecule undergoes retro-isomerization, meaning that the ratio of isomers in the final product is variable. This is vital because lycopene isomerization can be directed to maintain its stability, solubility, and bioaccessibility in food.
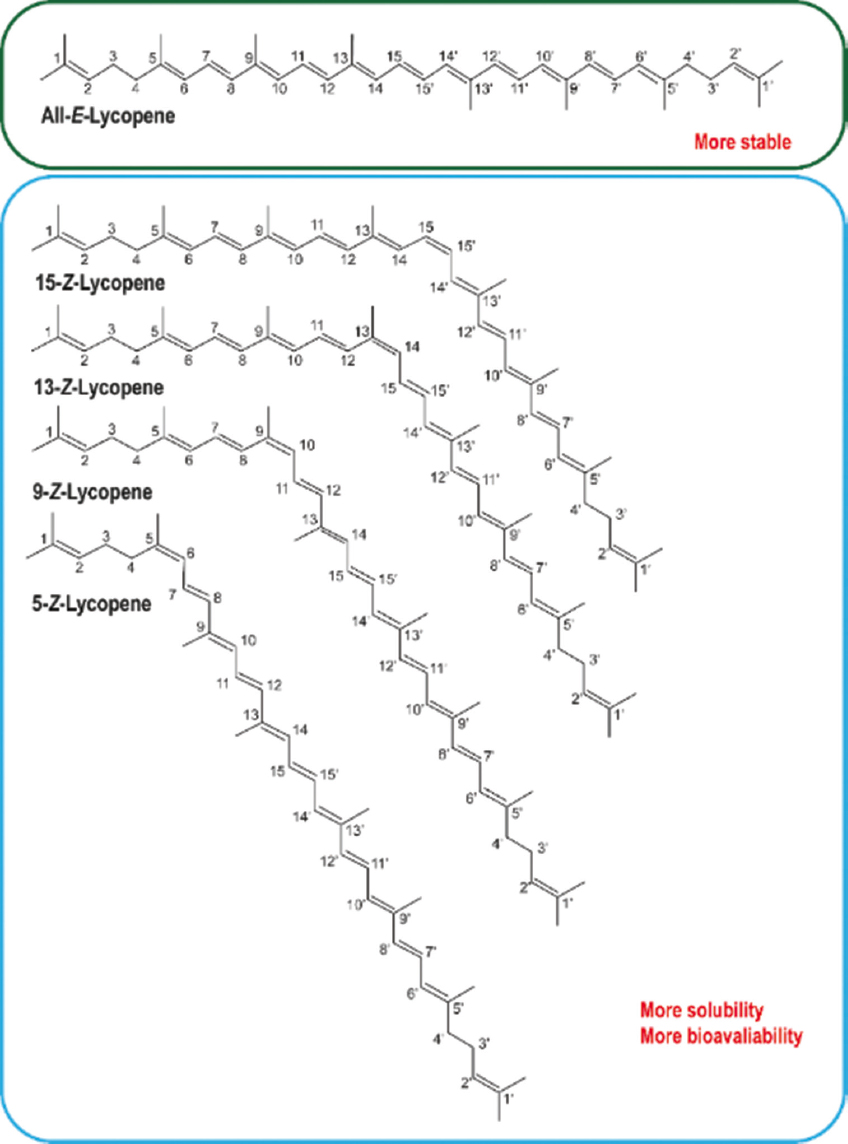 | Figure 2: Chemical structure of various lycopene isomers. Figure our own, created using Adobe Illustrator (Adobe, Inc., San Jose, CA, USA). [Click here to view] |
Lycopene is predominantly found in its trans-form because it is more thermodynamically stable than its cis-isomers. An example is the tomato, which is one of the most common sources of lycopene, where the natural trans-configuration exists, and the presence of cis-lycopene can probably be derived as a result of the storage process or treatment of the material [2,3]. The cis-isomers show higher bioavailability owing to their lower aggregation and better solubility in bile micelles, whereas the trans-isomers crystallize easily, which limits their absorption [2,3]. Both cis- and trans-isomers can be synthesized naturally through plants or by microbial fermentation [28]. In this sense, the form in which lycopene is present provides specific characteristics and properties that are potentially useful, as described below.
The bioavailability and antioxidant properties of lycopene are influenced by the presence of geometric isomers and the position of isomerization; Z-isomers have higher antioxidant activity and bioavailability than (all-E)-lycopene. The ratio of lycopene isomerization affects its biological properties. Wang et al. [40] evaluated lycopene samples containing 5%, 30%, and 55% Z-isomers, and found that the 55% total Z ratio of lycopene revealed the highest radical (2,2-diphenyl-1-picrylhydrazyl) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) scavenging activity. This indicates that the Z-isomers of lycopene, rather than the isomer (all-E), have a greater health impact; however, Z-isomers are less stable. However, as mentioned above, this response varies depending on how the raw material is processed. However, factors such as the intensity of light incident on the fruit or heat and oxidation during fruit processing can modify the isomeric pattern of lycopene, altering its final composition. It is important to note that some inconsistencies in isomerization data may lead to biases in terms of their actual impact on health, whether in terms of stability and/or antioxidant capacity. Regardless of the isomer, the main sources of lycopene are fruits, vegetables, and processed foods.
3. CROPS ARE A MAJOR SOURCE OF LYCOPENE
This carotenoid pigment tends to accumulate differentially in the plants (0.01–15,000 mg·100/g fresh weight [FW]), depending on a variety of factors. In general, lycopene is regulated by the differential expression of genes associated with carotenoid biosynthesis during the ripening process, as transcripts of the lycopene-inducing enzymes phytoene synthase (PSY) and phytoene desaturase (PDS) are increased, whereas transcripts of the β- and ε-lycopene cyclase (associated with the conversion of lycopene to β- or δ-carotene, respectively) are reduced or disappear completely during this process [41].
There are primary and secondary sources of lycopene, with tomatoes and peppers being the most important primary sources. Secondary sources include watermelon, carrot, pumpkin, grapefruit, guava, papaya, oranges, mandarin, apricot, red amaranth, olive, horse celery, pitanga, wild rose fruits, and other less common fruits such as mertajam, yellow Himalayan raspberry, nopal fruits, firethorn fruits or pyracanta, persimmon fruits, and some flowers [Table 1], gac (Momordica cochinchinensis) arils [42], species of the genus Citrus (0.35–4.39 mg lycopene per 100 g FW) [43], and Prunus (0.01–0.05 mg·100/g FW) [1].
Table 1: Average lycopene content in different crops.
| Crop | Content (mg·100/g) | Potential lycopene yields (kg/ha) | References |
|---|---|---|---|
| Purple amaranth (Amaranthus gangeticus L.) | 5.019§ | 0.018 | [44] |
| Ash gourd (Benincasa hispida Thunb). Pumpkin (Cucurbita pepo L.). Night-flowering jasmine or Harshringar (Nyctanthes arbor-tristis L.) | 0.34–4.83 of flower petals. | NS | [45] |
| Green bell pepper (Capsicum annuum L.). | 0.265–62.0§ | 1.184 | [46] |
| Red bell pepper (Capsicum annuum L.) | 1,698§ | 32.423 | [46] |
| Bell pepper “Cannon” (Capsicum annuum L.) | 22.96§ | 1.492 | [47] |
| Bell pepper “Bragi” (Capsicum annuum L.) | 12.95§ | 0.842 | [47] |
| Papaya (Carica papaya L.) | 0.02–7.56§ | 0.227 | [1,48-50] |
| Watermelon (Citrullus lanatus L.) | 1.17–13.52§ | 0.460 | [1,48,51] |
| Chinese grapefruit (Citrus maxima [Burm.] Merr) | 0.50§ | 0.024 | [43] |
| Mandarin (Citrus reticulata cv “Shatangju”). | 0.321–0.343§ | 0.005 | [52] |
| Sweet orange tree (Citrus sinensis L. Osbeck) | 0.8§ | 0.006 | [53] |
| Yellow zucchini (Cucurbita moschata L.). | 17.85–26.4 edible portion | 0.330 | [54] |
| Cucurbita sp | 0.38–0.50§ | 0.006 | [1] |
| Carrot (Daucus carota L.) | 0.65–0.78§ | 0.0031 | [1] |
| Ripe fruit of autumn olive (Elaeagnus umbellata Thunb). | 3.0–4.0§ | 0.00002 | [55] |
| Pitanga (Eugenia uniflora L.) | 0.26§ | 0.00001 | [56] |
| Mertajam (Lepisanthes rubiginosa [Roxb.] Leenh) | 158.0§ | NS | [57] |
| Apricot (Prunus armeniaca L.) | 0.0005–0.1760§ | 0.001 | [58] |
| Guava (Psidium guajava L.) | 0.769–7.64§ | 0.115 | [1,48,59,60] |
| Rosa sp. | 0.68–0.71§ | 0.0000004 | [1,59] |
| Yellow Himalayan raspberry (Rubus ellipticus Smith.) | 1.0–1.3§ | 0.0000034 | [61] |
| Celery horse (Smyrnium olusatrum L.) | 8.00–49.45§ | NS | [62] |
| Tomato (Solanum lycopersicum L.) | 0.0350–670§ | 46.304 | [1,5,59,63–69] |
| Pepper (Capsicum annuum L.) | 307.99–11,800¥ | 11.789 | [70] |
| Capsicum chinense L. | 14,200¥ | 31.11 | [70] |
| Green scotch bonnet pepper (Capsicum chinense L.) | Flesh: 7.63¥ Flesh and seed: 6.87¥ | 0.02 | [71] |
| Red scotch bonnet pepper (Capsicum chinense L.) | Flesh: 8.60¥ Flesh and seed: 8.31¥ Seed: 7.56¥ | 0.02 | [71] |
| Pomelo – Grapefruit (Citrus grandis “Majiayou”). | 0.01–0.02¥ | 0.0001 | [72] |
| Carrot (Daucus carota subsp. sativus) | 10.0–40.0¥ | 0.16 | [73] |
| Persimmon fruit (Diospyros kaki L.) | 0.0061–0.127¥ | 0.0003 | [74] |
| Tuna (Opuntia ficus indica L.) | 1.038–2.82¥ | 0.0020 | [75] |
| Wild rose bush (Rosa canina L.) | 4.32–12.22¥ | 0.000001 | [76] |
| Tomato (Solanum lycopersicum L.) | 10.0–2,939¥ | 10.698 | [77–80] |
A common and highly studied source of lycopene is tomato fruit; however, its content differs in the peel, whole berry, pulp, and seed [68], and according to the cultivar and its development environment [77]. Watermelon [81], apricot [58], and guava [60] accumulated lycopene differentially in response to genetic characteristics between varieties as well as plant development conditions. Another source of lycopene is Capsicum fruits, such as peppers [46,47,70], and green and red habaneros [70,71]. In these species, lycopene accumulation is partially caused by the expression of phytoene synthase 1 (PSY1), capsanthin-capsorubin synthase, and pseudo-response regulator 2 genes. In the case of some carrots with reddish pigmentation, low expression of the lycopene ε-cyclase gene (DcLCYE) leads to the accumulation of lycopene [73], and the same response occurs in certain pumpkin cultivars [54].
In some fruits, such as red-fleshed sweet oranges from the tropics [53] and grapefruit, lycopene accumulation occurs in the pulp during the ripening process, and in the latter case, owing to higher expression of the PSY gene associated with lower expression of CCD1 [72]. Another source of lycopene is the ripe fruit of autumn olives (Elaeagnus umbellata Thunb.) that accumulates lycopene because during ripening, PSY (EutPSY gene) is highly expressed, whereas the enzyme lycopene ε-cyclase (EutLCYe gene) only shows some activity during some periods of fruit ripening [55].
Considering a potential yield of ~31 and ~46 kg/ha of lycopene per ha, pepper and tomato cultivation are the most plausible alternatives for supplying this molecule to the agricultural sector and industry. In the case of crops with lower lycopene content (watermelon, papaya, yellow zucchini, and guava), agricultural management strategies should be implemented to promote the concentration of this carotenoid. Likewise, crops with very low lycopene content would not be a reliable source of lycopene for the sector.
4. LYCOPENE: RESPONSE TO AGRICULTURAL CONDITIONS
In plants, lycopene is associated with photosynthetic complexes, which function as a photoprotectant by participating in singlet oxygen (1O2) quenching, scavenging peroxyl radicals [41], and as a substrate for the biosynthesis of abscisic acid (ABA) or strigolactones (SLs). Lycopene biosynthesis starts in plastids from two molecules of geranylgeranyl diphosphate, forming phytoene thanks to the enzymes PSY1 and PSY2. Next, PDS gives rise to ς-carotene, which is converted to lycopene by ς-carotene desaturase (ZDS) and carotene isomerase (CrtISO). However, it is here that the environmental exposures can condition the accumulation of lycopene, since this carotenoid undergoes cyclization at its ends with the intervention of lycopene δ-cyclase (LCYE) or lycopene β-cyclase (LCYB1 and LCYB2) originating from δ-carotene or γ-carotene, respectively. These two molecules originate from α-carotene and β-carotene via LCYB [41]. In this sense, farmers could manipulate the light environment of a crop (containing lycopene) to promote and increase its synthesis.
Hydroxylation of α-carotene at C3 is mediated by the enzymes β-carotene hydroxylase (CHXB) and ε-carotene hydroxylase, which generate lutein, whereas β-carotene is converted to β-cryptoxanthin, which gives rise to zeaxanthin (thanks to the enzyme CHXB). Zeaxanthin, by the enzyme violaxanthin de-epoxidase, generates all-trans-violaxanthin, which serves as a substrate for neoxanthin synthase, and the enzyme zeaxanthin epoxidase (ZEP) can reconvert violaxanthin to zeaxanthin (xanthophyll cycle that participates as a mediator under conditions of high solar radiation). Finally, 9-cis-violaxanthin is generated, which, together with 9’-cis-neoxanthin, is oxidized and originates from cis-xanthoxin. Oxidation of xanthoxin in the cytosol by ABA-aldehyde originates from ABA, and xanthoxin can be a substrate for SLs synthesis in the cytoplasm. This pathway is altered during the ripening process, increasing its content during the transition from ripe green (50 mg·100/g) to ripe red (100 mg·100/g) [82]. Based on the above, farmers can focus on crop management during the ripening-fruiting stage to induce greater lycopene synthesis and accumulation.
4.1. Lycopene Behavior in Response to Environment
4.1.1. Light/radiation
Radiation is a determining factor in lycopene content and biosynthesis, and is modulated by phytochrome-interacting factor (PIF) transcription factors. During the development of the immature green fruit until its final mature green size, PIF transcripts remained constant, but in the orange phase, they doubled, and in the mature red fruit stage, they increased 5 times more [83]. The same authors pointed out that ripe red fruits exposed to light have higher lycopene and β-carotene content than ripe green fruits that contain higher chlorophylls.
PIF regulates carotenoid gene expression in light environments (direct and/or diffuse) and at various temperatures. Curiously, in tomatoes, as the fruits ripen, the light reaching the green fruit flesh creates a self-shading microenvironment that induces high levels of PIF1a and phytochrome Pr in its inactive form (repressing the carotenoid catalytic PSY1 gene). Upon the initiation of the maturation development program, exocarp chlorophyll is degraded, minimizing self-shading, and phytochrome Pr is converted to its active form, Pfr, by photoreversibility. This change causes the PIF1a proteins in the pulp to be degraded, allowing the synthesis of lycopene (by PSY1 expression), which is perceived as a signal that censors the pigment profile, allowing the coordination of color transition and carotenoid accumulation [84].
In this context, tomatoes exposed to intra-canopy light-emitting diode (LED) light significantly increase in lycopene content which may be due to the fact that lycopene accumulation responds to the red/far red ratio by phytochrome signaling, in that sense intra-canopy irradiance with LED may be a technique to increase lycopene by modifying the expression of PSY gene and its isoforms PSY, PSY1, and PSY3 [85].
Similarly, in Capsicum cultivars, lycopene content and accumulation respond strongly to the light environments to which the plant and fruit are exposed, altering the differential expression of genes associated with carotene synthesis [86,87]. In contrast, in open- and net-grown mandarin fruits, lycopene content was between 0.321 and 0.343 mg·100/g, respectively. These responses may be due to modifications in the PSY, carotenoid cleavage dioxygenase 4, and ZEP genes [52]. As a post-harvest treatment, irradiation of green fruit in phase 1 with red light (12 h/day for 15 days and stored in darkness for an additional 6 days or irradiated continuously with red light for 21 days) causes changes in the outer layer of the tomato fruit [88]. The lycopene content in plants is strongly altered (besides variety, cultivar origin, and nutritional management) by interaction with the environment and stressors (abiotic and biotic), such as high/low light intensity, salinity, water deficit, metalloids, and nutrient deficit [Figure 3].
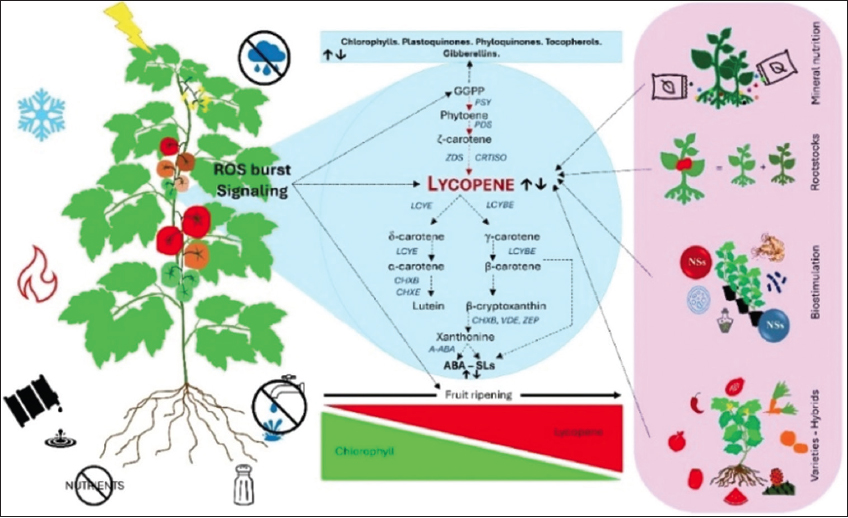 | Figure 3: Lycopene – environment interaction and strategies to promote its content in plants. Environmental conditions can cause an increase or an oxidative burst of ROS, which is accompanied by an increase in cytosolic Ca2+, inducing signaling at the cellular level that (i) alters lycopene biosynthesis in leaves and fruits by promoting an increased synthesis of chlorophylls, plastoquinones, phyloquinones, tocopherols, or gibberellins, or (ii) causes cyclization of lycopene to generate molecules such as β-carotene, lutein, zeaxanthin, abscisic acid, and strigolactones. Likewise, lycopene responds to the ripening process of the fruit; therefore, during this stage, lycopene content can be strongly affected by agronomic management conditions and environmental stresses. Alterations in lycopene content are a function of the genetic load of the plant as well as the type, timing, duration, and intensity of environmental stressors. In this situation, a series of strategies can alleviate the damage or promote a greater accumulation of lycopene in the fruit, such as mineral nutrition, rootstocks, biostimulation (proteins, chitosan, microorganisms, and nanomaterials), and the use of hybrid varieties with a greater capacity for carotene synthesis. Figure our own, created using Microsoft Office PowerPoint (Microsoft Corporation, Redmond, Washington, USA). [Click here to view] |
4.1.2. Water regime
The water regime strongly affects the lycopene content of the fruit because an adequate water supply promotes higher lycopene content, whereas a water deficit induces greater accumulation of β-carotene, reducing lycopene content [78]. A similar response was reported when supplying 100, 75, and 50 of crop evapotranspiration until the beginning of ripening, as it modifies lycopene concentration [64]. Interestingly, under drought conditions, lycopene in tomato fruit increases from ≈0.65 to 0.90 mg·100/g [65]; the same trend is seen in non-irrigated or rehydrated tomato plants after drought, where lycopene content increased significantly compared to fully irrigated tomato plants [66].
Similarly, the Italian tomato cultivar Quarantino, developed under drought conditions, increased significantly in lycopene content in the skin and pulp [89]. This trend has also been reported in tomatoes grown in sandy loam and silty loam soils under deficit irrigation and in the indigenous watermelon variety Sistani [81]. In cotton grown under drought stress, lycopene is transformed to lutein by the enzyme lycopene cyclase, as evidenced by the high expression of the lycopene ε-cyclase gene GhLCYε-3 [90]. Since water restriction generates oxidative damage, the increase in lycopene may be due to the regulation of metabolism that seeks to cope with stress by allowing the plant to acclimatize and develop by detoxifying ROS. Therefore, water resource management is a tool that can condition lycopene content in plants; however, prolonged or extreme drought conditions can be counterproductive, significantly reducing lycopene content [9].
The contrasting responses of lycopene to water stress conditions could be due to (i) the plant’s ability to cope with stress either by increasing its secondary metabolism or by promoting the methyl erythritol diphosphate pathway, (ii) the phenological stage at which stress is applied, since its nutrient reserves condition its response, (iii) the level of stress to which the plant is subjected, since carotenoids can act as antioxidants; (iv) the duration of water deprivation, as prolonged periods will reduce the plant’s response capacity.
4.1.3. Salinity and metalloids
Another environmental condition is salt stress, which has been suggested to increase lycopene content [91] as a primary mechanism counteracting the negative effects of stress by inhibiting the enzyme lycopene-β-cyclase or increasing the precursor phytoene content [63]; however, this response depends on the concentration and duration of exposure to salt stress. Regarding exposure to metalloids, it has been reported that under As stress (0.2 mg/L), lycopene content in tomato fruit increases, whereas applying 1000 mg/L of NPs-SiO2 [77]. Exposure to Cr(VI), 10 mg/L of K2Cr2O7, promote a significant increase in lycopene content with respect to the control treatment, which may be due to the increased expression of genes associated with carotenoid biosynthesis (ZDS, SlZDS; carotenoid isomerase, SlCRTISO; and LCYB, SlbLCY) [92].
4.1.4. Temperature
Another relevant factor in lycopene accumulation is growth temperature; it has been reported that Citrus Maxima [Burm] Merr presents higher lycopene content (0.45 mg·100/g) in warm seasons (20–22°C) compared to the cold or rainy season (0.3 mg·100/g). The latter response may be due to the inhibition of enzymes associated with the carotenoid pathway, which promotes rapid conversion of lycopene to β-carotene [43]. The response to temperature stress (high/low) is contrasting, as a decrease in lycopene content is reported in plants grown at high temperature [93], whereas in high temperature-tolerant hybrid tomato, the accumulation of lycopene is promoted, suggesting higher antioxidant activity and better tolerance capacity to this stress [94]. In another study, low- (4°C) or high-temperature (32°C) stress did not severely alter lycopene content relative to the control treatment (25°C) [95]. Interestingly, in tomato fruits, high microRNA (Sly-miR167a) expression promotes lycopene accumulation under cold stress [96].
In this context, the data suggest that lycopene biosynthesis appears to respond hormetically to stress conditions. Therefore, we propose the development of mathematical models that allow the projection of the most favorable “microstress” conditions for improving lycopene in crops.
However, it is necessary to establish that the lycopene content in fresh fruit does not represent the actual content; therefore, thermal handling of the fruit leads to an increase in bioaccessible lycopene owing to the release of phytochemicals from the matrix, which makes it more accessible [79].
4.1.5. Nutrient stresses
Other factors that alter lycopene content are Cu2+ deficiency, which reduces lycopene content in tomato fruits [97].
4.1.6. Biotic stresses
In the tomato yellow leaf curl virus stress, which decreases lycopene levels from 1.412 mg·100/g in uninfected plants to 0.979 mg·100/g in infected plants [98]. Evidently, a combination of several stresses can also occur; in this regard, it has been documented that in tomato fruit under salinity (60 mM NaCl) and heat (35°C), the lycopene content is not modified with respect to the control; however, when combining these stresses, the lycopene content increases significantly. This response may be due to the degradation of lycopene precursors (phytoene and phytofluene) while maintaining adequate levels of β-carotene despite a reduction in violaxanthin content [99].
4.2. Strategies to Increase Lycopene Content in Crops
4.2.1. Agronomic practices
Knowing the pathways and factors that affect lycopene synthesis, several agronomic tools can be used to increase the content of this carotenoid in the crops of interest. These strategies include grafting, nutrient management, biostimulation, and manipulation of genetic resources (traditional and genetic engineering) [Figure 3].
In this regard, it has been documented that the electrical conductivity of a nutrient solution alters the lycopene content in bell pepper cv. California Wonder [7], whereas grafting tomato increased the lycopene content of the fruit [6]. On the other hand, the contribution of foliar-sprayed silicon (Si) in tomato (0.6–1.8 mM) increases lycopene content due to the activation of PDS, lycopene ε-cyclase (LCY-E), and zeaxanthin cyclooxygenase (ZEP) [39].
4.2.2. Biostimulants
Recently, biostimulation as a tool to promote lycopene content in fruit seems to be a feasible option, and recent studies have shown that the application of protein hydrolysate in tomato increases lycopene content in fruit owing to the activation of metabolism [65]. Likewise, the use of Malvaceae-derived protein hydrolysate increases the lycopene content in tomatoes [100]. Similar reports have shown that periodic applications of bovine gelatin-based protein gels and keratin hydrolysates in the root system of tomato plants during the early vegetative stage promote between 1.23 and 1.44 times the lycopene content in the fruit [101]. Similarly, the application of Chaetomorpha antennina macroalgal extracts increased lycopene content in tomatoes under salt stress, probably because of the sulfate and carbohydrate groups of the macroalgae [102].
Another biostimulant reported is chitosan in tomato plants (20 mg/L), which has led to an increase in lycopene content thanks to the promotion of its synthesis by the high nitrogen content in the leaf [8]. Nitric oxide (sodium nitroprusside) increases lycopene levels [103], probably by blocking β-cyclase activity. On the other hand, the exogenous use of 5-Aminolevulinic acid (200 mg/L) on tomato fruits strongly promotes lycopene biosynthesis due to the positive regulation of geranylgeranyl diphosphate synthase (GGPPS) genes, PSY1, PDS, and LCYB in concert with reduced expression of Mg-chelatase (CHLH) and protochlorophyllide oxidoreductase genes associated with chlorophyll synthesis [104,105]. In contrast, during tomato fruit ripening, hydrogen sulfide (H2S) decreases the expression of genes associated with lycopene synthesis and enhances the enzymatic activity and expression of genes associated with chlorophyll synthesis [106].
Similarly, the biostimulants Nomoren (20% arbuscular mycorrhizal fungi, Glomus spp.), Twin Antistress (Bacillus subtilis, yeast, Ascophyllum nodosum extracts, 1% organic N, 10% organic carbon, and 30% organic matter), and x-stress (0. 5% Cu, 2.0% Fe, 1.0% Mn, and 2.0% Zn, all chelated with glycine) promoted lycopene content in tomato fruit [107]. It has also been reported that biostimulants based on amino acids and soluble peptides increase lycopene content, but this response is highly dependent on the production season [9].
These biostimulants differentially impact lycopene metabolism in plants, although it cannot be ruled out that their combined application could show a certain level of synergy in promoting lycopene. In this regard, and in order to improve and streamline lycopene biosynthesis in crops, it is proposed to apply biostimulants via foliar application and drenching, so that these molecules can enter the plant quickly (via foliar application) and constantly (via drenching), which would maintain continuous stimulation of lycopene-associated metabolism. In addition, biostimulation could improve crop yield and quality, making the field practice potentially accessible and profitable.
4.2.3. Nanotechnology
Recent nanotechnological innovations have been shown as another alternative to promote lycopene; for example, the use of water-soluble hyper-harmonized hydroxyl-modified fullerene (200 mg/L) on cherry tomatoes considerably increases lycopene content, probably by proper activation of plant secondary metabolism [67]. Iron oxide nanoparticles (100 and 300 mg/L) in tomato under Cd stress promoted lycopene accumulation in fruit [108], and zinc oxide nanoparticles primed with proline and betaine (50 and 100 mg·100/g soil) increased lycopene content [109]. Finally, selenium nanoparticles induced an increase in lycopene content [110].
4.2.4. Genetic approaches
Another way to increase lycopene content is by genetic improvement, either through traditional breeding or genetic engineering. For example, in high-lycopene tomatoes, geranylgeranyl pyrophosphate synthase 1, 2, and 3 (GGPPS1, GGPPS2, and GGPPS3), TPT1, SSU II, PSY2, ZDS, CrtISO, CrtISO-like 1 (CrtISO-L1), PSY1, PDS, and ς-carotene isomerase showed higher differential expression, maximizing the metabolic flux for carotenoid (lycopene) biosynthesis. Surprisingly, the conversion of lycopene to carotenoids was prevented by β-LCY2 and ?-lycopene cyclase (ε-LCY) genes. Similarly, differential expression of β-carotene hydroxylase 1, ZEP, violaxanthin desaturase (VDE), and CYP97C11 genes appears to reduce lycopene content in tomato [111]. While in a tomato modified with high pigmentation [hp] genes (HLT-F71), the lycopene content can reach 14.05 mg·100/g [69].
Tomatoes with high lycopene content, developed by introgression of hp and old-gold (og) mutations, amplify the phytochrome-mediated response to light, causing elevated levels of carotenoids, mainly lycopene, and increasing their content with respect to a standard tomato [69]. Similarly, by Targeting Induced Local Lesions IN Genomes, a variant of the lycopene ε-cyclase gene (SlLCY-E) (G/3378/T) alters carotenoid content in tomato plants, promoting a significant increase in β,β-xanthophylls in leaves, whereas lycopene and total carotenoids are increased in ripe fruits. This allelic variant, as a genetic resource, induces tolerance to drought stress and increases lycopene and carotenoid levels in fruits [10].
Likewise, in tomato lines silenced in the microRNA Sly-miR164a, in addition to promoting resistance to salt stress, lycopene content in the fruit was improved [112]. Similarly, in tomatoes, the MYB transcription factor (SlMYB1) appears to bind to the SlLCY1 and SlPSY2 genes, which are associated with lycopene synthesis [113]. The chromoplast-specific carotenoid-associated protein (CHRC) promoter, a member of the plastoglobulins or fibrillins (plastid lipid-associated proteins), appears to play a role in the development and accumulation of carotenoids, such as lycopene, whereas in tomato, the SlCHRC gene regulates the biosynthesis and accumulation of carotenoids during the ripening process, participates in the transition from chloroplasts to chromoplasts, and alters the amount and volume of plastoglobules [11].
In reddish-pigmented carrots, the accumulation of lycopene appears to be due to the low abundance of lycopene ε-cyclase gene (DcLCYE) transcripts and low levels of α-carotene. In contrast, in orange carrots, lycopene is transformed to α-carotene and β-carotene by the high expression of the LCYE and LCYB genes [73]. Therefore, the expression of the PSY gene (EutPSY) can be used as an effective strategy to increase lycopene content in fruits to improve their quality [55].
In this context, overexpression and Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-associated protein 9-mediated gene editing techniques appear promising, but further study is needed. That is why we believe that the use of advanced tomato lines enriched with lycopene using conventional methods is more promising for implementation in commercial tomato cultivation programs (where light-sensitive hp mutation is encouraged) [69]. In addition, these breeding programs are developed in open field conditions, which stabilizes the results obtained.
It is important to note that all of the above strategies are closely associated with the environmental modulation in which the crop is grown. In this regard, farmers and producers can combine these strategies to enhance lycopene accumulation. For example, efficient irrigation management can improve the impact of biostimulants, and these products can even be applied via drenching. Likewise, in the specific case of grafting, it is necessary to modulate factors such as relative humidity and light environments to ensure the promotion of lycopene in the plant.
These data provide fundamental information for increasing lycopene content in fruits by manipulating the aforementioned genes to improve the contribution of this biomolecule to human diets, thus promoting human health.
5. THE ROLE OF LYCOPENE IN HUMAN HEALTH
5.1. Lycopene’s Bioaccessibility
As an antioxidant biomolecule, lycopene can enter the human diet via fresh or processed foods. However, only a fraction of the food component is released from the solid food matrix and remains available for absorption in the gastrointestinal tract. Bioaccessibility is influenced by factors such as food-processing methods, cooking techniques, diet composition, chewing, isomeric configurations of lycopene, and interactions with probiotics [114].
Food processing plays a crucial role in enhancing lycopene bioaccessibility. By altering the cell walls and weakening the binding forces between lycopene and the tissue matrix, processing methods increase the availability of lycopene for absorption; in tomato juice fermented with Saccharomyces cerevisiae ATCC 9763 the bioaccessibility of lycopene was 45.1 mg·100/g FW due to the disorganization and hydrolyzation of cell walls as well as by a lower pectin content and higher formation of mixed micelles [115]. Consequently, lycopene bioaccessibility is lowest in raw foods, moderately improved in lightly processed foods, and reaches its maximum in thermally processed foods and purified oil preparations. However, the benefits of this processing can be offset by improper storage and handling. Therefore, the quality and quantity of lycopene in food may affect its absorption levels, as well as the metabolism rate during its ingestion.
Unfortunately, processing methods that promote bioavailability can impact and/or alter the structure and content of lycopene, requiring in-depth studies and standardized methods that minimize loss or improve the efficiency of its total content in the food matrix. Thus, new technologies such as the use of Pickering emulsions, micelles, liposomes, bigels, pearls, microcapsules, nanoparticles, and electrospinning [12], as well as the use of technologies such as hot–melt extrusion [116], high-pressure homogenization [117], in combination with additives such as olive or corn oil, digestible lipids, whey protein isolate, and sodium alginate [118,119] can stabilize the molecule by preventing its degradation.
5.2. Lycopene’s Absorption, Bioavailability, and Metabolism
5.2.1. Bioavailability
Lycopene is absorbed similarly to lipids [3,37] orally, gastric and intestinal phases [Figure 4] through the processes of (i) release, (ii) emulsification, (iii) micellar, and (iv) transport [12].
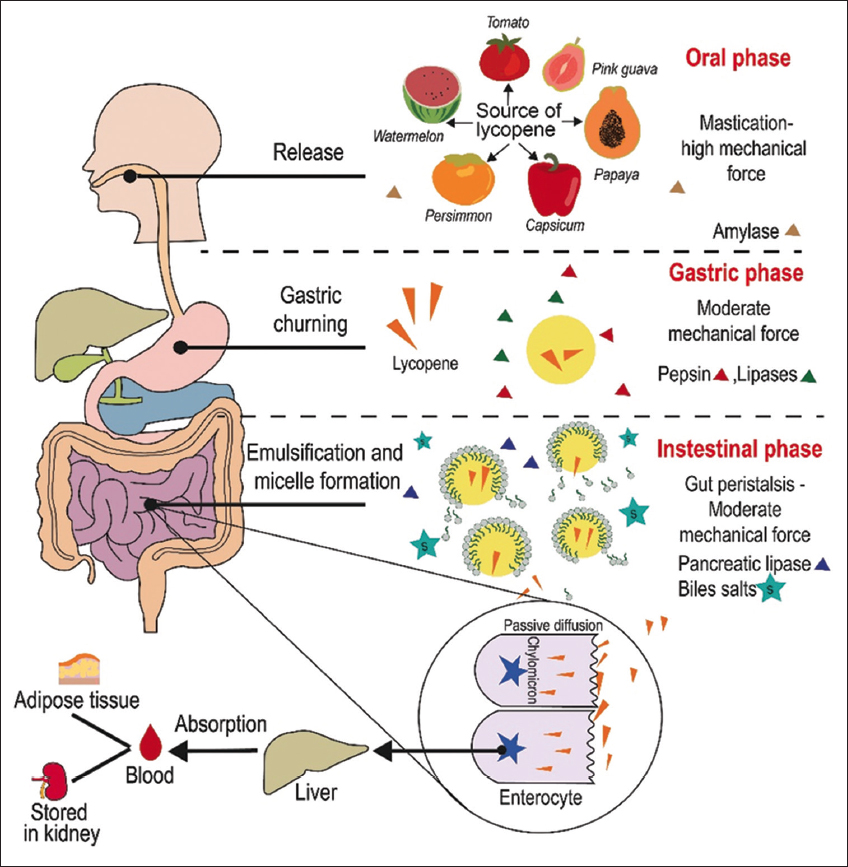 | Figure 4: Lycopene digestion scheme. Adapted from Wu et al. [12]. Created using Adobe Illustrator (Adobe, Inc., San Jose, CA, USA). [Click here to view] |
5.2.2. Gastrointestinal absorption
The oral phase constitutes the first stage of ingestion of the lycopene-loaded food matrix, and mechanical force (chewing) promotes matrix disruption and enhances lycopene release [120]. In the stomach, gastric conditions (low pH, pepsin, amylase, gastric lipase, mechanical forces, and peristalsis) favor the partial release of cis- and trans-isomers of lycopene from the food matrix and its dissolution in the lipid phase. Subsequently, lycopene can be integrated into lipid droplets that pass into the duodenum (small intestine), where pancreatic lipase enzymes and bile acids act, breaking them down into multilamellar lipid vesicles and micelles, favoring absorption by enterocytes. The inclusion of carotenoids in micelles depends on the configuration/structure or composition of micellar lipids, causing limited absorption [3,37,120]. Therefore, consumption of fats in conjunction with lycopene-rich foods may increase the efficiency of their absorption.
It has been proposed that most of the lycopene is packaged into chylomicrons and, upon leaving the mucosal cells, these chylomicrons are subsequently transferred to the liver through the mesenteric lymphatic system [120]. Lipoprotein lipase acts on these chylomicrons, allowing lycopene and other carotenoids to be passively taken up by adrenal glands, kidneys, adipose tissue, spleen, lungs, prostate, and reproductive organs [3,120]. Subsequently, residual chylomicron residues are eliminated from the liver through chylomicron receptors. In the liver, carotenoids can accumulate or be repacked into very-low-density lipoproteins and released back into the bloodstream [120].
5.2.3. Transport and distribution
Lycopene is transported in plasma within the low-density lipoprotein (LDL) core, a crucial mechanism for its distribution throughout the body. For this transport to occur, lycopene must first be incorporated into lipoprotein molecules, a process that primarily occurs in the hepatocytes and intestinal epithelial cells. In particular, the cis-isomers of lycopene are more readily incorporated into lipoproteins than the trans-isomers. That is, the cis-forms are more bioavailable than the all-trans forms, and their accumulation is inversely related to androgenic status and potentially influenced by energy intake.
While the release of lycopene from raw materials has a significant impact on its bioavailability, the fraction reaching systemic circulation is influenced by factors such as the dose and the presence of other carotenoids, such as β-carotene [25].
5.2.4. Factors influencing bioavailability
Considering that the recommended daily intake (RDI) of lycopene in humans ranges between ~2 and ~20 mg/day [37], some factors may alter the efficacy of lycopene in the body by decreasing its absorption to 2–30% of the total amount ingested [121]. Studies suggest that genetic factors such as age, sex, body composition, hormonal status, blood lipid levels, alcohol and tobacco consumption, tissue redox state, presence of other carotenoids in the diet, and other dietary components affect lycopene absorption [37]. The sex of an individual modifies the lycopene absorption efficiency. It has been observed that postmenopausal women absorb lycopene differently from men of the same age, which could be associated with hormonal variations and body composition [122]. Due to its lipophilic nature, lycopene is influenced by diet, suggesting a minimum fat threshold to promote its absorption in different food matrices [116]. However, it can be inhibited by excess dietary fiber and β-carotene. These matrices and their processing condition the isomeric profile (cis/trans) and condition the preferential absorption of the cis isomer. In this context, we suggest that diet plays a greater relative role in lycopene metabolism.
Considering that the efficiency of lycopene absorption, distribution, and metabolism in the body is affected by a number of factors, it is recommended to consider these aspects when evaluating its consumption and potential health benefits, in addition to discerning the implications of excessive consumption of this carotenoid (lycopenemia) [37]. Finally, it has been reported that people with some pathologies, such as cancer and cardiovascular diseases, demand up to 35–75 mg/day [37]. To achieve the recommended average daily intake, as well as the intake required for certain pathologies, the most practical food sources could be fresh tomatoes and peppers, as well as their derivative products (sauces). We also suggest consuming fruits with low lycopene content (watermelon, papaya, yellow zucchini, and guava). Similarly, the consumption of tablets is another plausible alternative. The above would lead to an increase in demand for both lycopene-rich crops and lycopene tablets in the pharmaceutical sector, stimulating growth in these sectors. After its consumption and absorption throughout the body, lycopene has an impact on different organs participating in and/or promoting human health directly or indirectly, as suggested in the following section.
The existence of discrepancies in lycopene bioavailability data may be due to the variety of food matrices, methods, and study designs that influence the standardization of average lycopene values for the human diet. In this context, we suggest establishing a “potential intake” of lycopene, considering temperature and processing methods as the main covariates. In this regard, in-depth studies should be conducted to address this knowledge gap and establish permissible temperature limits and the best extraction methods. Furthermore, the lack of standardized bioavailability metrics represents a gap in research that may limit comparisons between different food matrices, preventing their weighting against each other. Similarly, this limitation may lead to potential biases in dietary recommendations.
5.3. Lycopene in Human Body Systems
Scientific evidence suggests that lycopene plays a positive role in human health by promoting protection in different body systems [Figure 5]. At the cardiovascular level, it decreases oxidative stress owing to its antioxidant and anti-inflammatory effects [13]. Similarly, lycopene presents a potential mechanism for inhibiting lipoprotein oxidation by improving endothelial function and reducing systemic inflammation. These effects contribute to the prevention of atherosclerosis and, thus, the risk of cardiovascular problems [14].
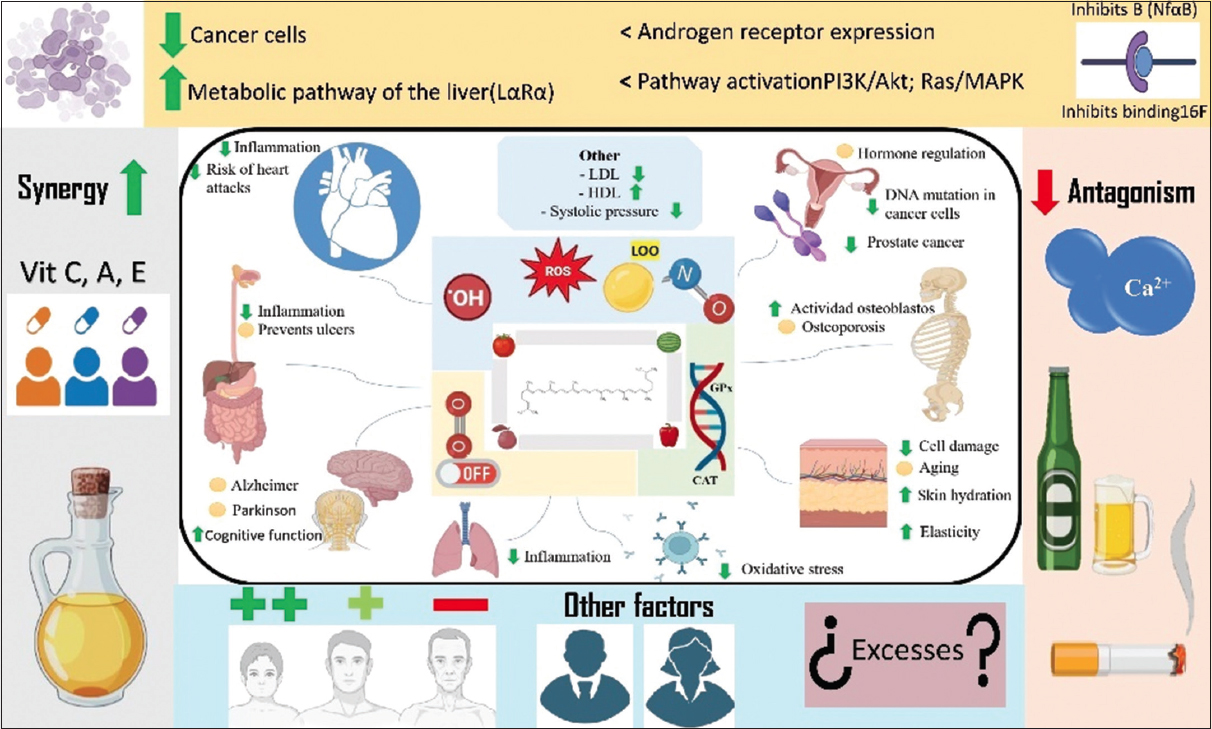 | Figure 5: Probable mechanisms of action of lycopene on human health. Figure our own, created using Microsoft Office PowerPoint (Microsoft Corporation, Redmond, Washington, USA). [Click here to view] |
Owing to its antioxidant properties, lycopene reduces oxidative stress and prevents immune-cell damage. In this environment, there is positive modulation of the immune system that improves the activity of T lymphocytes and other immune cells, allowing a greater defense capacity of the body in situations of infection or inflammation [15]. In contrast, lycopene seems to be involved in the endocrine system by regulating the hormonal load (estrogens) to improve reproductive health, which could contribute to the prevention of hormonal cancers, such as breast cancer. Similarly, several studies have associated lycopene with a molecule that participates in the prevention of prostate cancer owing to its protective effect against DNA mutations in prostate cells and abnormal cell proliferation [16,123].
Some studies have associated lycopene consumption with protection of the digestive system by reducing inflammation in the gastrointestinal tract and preventing gastric ulcers, which may be because lycopene protects the cells of the stomach lining from oxidative damage. Similarly, its antioxidant properties may help prevent gastrointestinal cancer, particularly colorectal cancer, by reducing the formation of carcinogenic compounds [17].
In the nervous system, lycopene exerts neuroprotective effects by preventing neuronal damage (owing to its antioxidant capacity), protecting brain cells against oxidative stress, the main factor associated with brain aging, and neurodegenerative diseases such as Alzheimer’s and Parkinson’s disease. It is also reported that lycopene can improve cognitive functions by attenuating age-related mental decline [18,124]. Lycopene promotes positive impacts on the skin. As an organ constantly exposed to solar radiation and ultraviolet (UV) rays, the skin is prone to accumulating free radicals responsible for premature aging and skin cancer, and acts as an antioxidant that attenuates and/or eliminates excess radicals, as well as improves elasticity and hydration [125].
Finally, some studies have suggested that lycopene may have protective effects on the respiratory system by reducing lung inflammation, which is beneficial in chronic respiratory diseases, such as asthma and Chronic Obstructive Pulmonary Disease. Studies have also suggested that lycopene protects the lungs from damage caused by pollutants and environmental toxins [19,126].
Although there is a gap in knowledge regarding the ideal doses of lycopene for human consumption, several studies offer a rough estimate of between 2.0 and 20.0 mg/day. These values would require specific knowledge of lycopene intake and selection of the food matrix with the highest bioavailability [37,127]. Therefore, it would be advisable to consume foods with a high lycopene content, such as tomatoes and peppers (and their derivatives), as well as taking supplements.
The intake and use of lycopene are important for health because of its beneficial properties against some specific diseases, such as its ability to reduce LDL cholesterol levels and increase high-density lipoprotein cholesterol levels [128]. In addition, it is believed to contribute to the improvement of endothelial function by promoting vessel dilation and efficient blood circulation [129]. It is also reported to have a protective effect against plaque formation in arteries, minimizing the risk of cardiovascular diseases, such as heart attacks and strokes [13], which are common in today’s societies.
It is also associated with the prevention of bone diseases such as osteoporosis by reducing oxidative stress in the bone and promoting the activity of osteoblastic cells, which are responsible for bone formation [130,131].
Overall, scientific evidence suggests that daily dietary lycopene intake could be an effective strategy to (i) improve human health and (ii) mitigate damage caused by lifestyle stress [15]. However, further studies are required to generalize and refine these findings, and it is likely that the potential impact of this biomolecule may also be affected by its interactions with nutrients and other biocompounds.
Here we show the favorable responses to lycopene consumption; however, factors such as bioavailability (e.g., due to the instability of lycopene isomers as a result of temperature), dosage, form of application, duration of treatment, and individual dietary patterns may generate contradictions in the results. Likewise, some methodological limitations, such as the restricted number of individuals, the absence of matched control groups, and the use of unvalidated instruments, could provide masked, contradictory, or less conclusive data. In this regard, there is a significant knowledge gap that prevents the establishment and validation of permissible lycopene intake limits that promote real benefits in each body system based on individuals’ lifestyles and metabolism, and therefore, further studies are required.
5.4. Lycopene-nutrients-bioactive Compounds Interactions
The interaction of lycopene with various nutrients and bioactive compounds may influence its absorption and potential effects on human health [12]. In human studies, synergism between lycopene and Vitamins C, E, and A, as well as xanthophylls and flavonoids, has been reported. This synergy enhances ROS neutralization and reduces oxidative stress and cell damage. In vegetables such as tomatoes, in addition to lycopene, compounds such as ascorbic acid, tocopherol, and β-carotene are found, which together form a complex network that can help reduce inflammation, delay aging processes, protect against degenerative diseases, contribute to the prevention of chronic diseases, and improve endothelial function, which could reduce the risk of heart disease, probably by mitigating the damaging oxidative effects of ROS. Surprisingly, the bioavailability and efficacy of lycopene improve when consumed with other antioxidants, as the combination seems to protect this biomolecule [38,48].
A fundamental factor in the interaction of lycopene with human health is the food matrix, in which the bioactive compound is included, since it has been demonstrated that the absorption process of this metabolite is directly influenced by this and its association with other nutrients. The isomerization of lycopene can also facilitate its absorption; however, it can be affected by factors such as the presence of oil, gastric pH, and the intestinal digestion process. For example, edible vegetable oils (peanut, olive, soybean, and canola) can enhance isomerization and lycopene absorption [26,132]. For example, incorporating dietary fats such as olive pomace oil and corn oil induces greater bioaccessibility of lycopene by 28.1% and 8.1% [119], so these oils may be recommended for use as additives in food matrices.
Carotenoids and minerals are another group of compounds that interact with lycopene. Simultaneous intake of lycopene with other carotenoids, such as lutein and β-carotene, can alter the amount of lycopene absorbed in the intestine [27,133]. Therefore, the consumption of a diet rich in fruits and vegetables that provides lycopene and other bioactive compounds could be associated with lower morbidity and mortality and greater longevity, partially because of the potential effect of lycopene [13,134]. This response may be due to the protective effect of lycopene against the radical molecules associated with cell damage.
5.5. Lycopene-free Radical Interactions
As a carotenoid with a highly conjugated chemical structure, lycopene has potent antioxidant capacity [Table 2]. Its interaction with free radicals is primarily determined by free radical scavenging. In this context, lycopene acts as a scavenger of ROS and RNS because of its multiple conjugated double bonds that allow it to donate electrons to hydroxyl radicals, superoxide, and lipid peroxides, as well as singlet oxygen (+O2) to neutralize them and prevent cell damage [21,135] by reducing or inhibiting lipid peroxidation, which protects cell membranes and LDL [22]. The impact on functional antioxidant activity in vivo may be due to the fact that (i) the amount of lycopene ingested can maximize or minimize its efficiency (depending on fat or fiber consumption, respectively), (ii) the processing method affects the lycopene content of the food matrix, and/or (iii) the alteration in the isomers present.
Table 2: The main health impact of lycopene.
| Mechanism | Action | Health effects | References |
|---|---|---|---|
| Antioxidant activity. | ↓ Oxidative stress. Prevents damage to lipids, proteins, and DNA. | Prevention of chronic diseases, cardiovascular diseases, and cancer. | [135,136] |
| Modulation of cellular signaling pathways. | Insulin-like growth factor pathway. Androgen receptor involvement. | Limits cell proliferation. Promotes apoptosis. | [20,136] |
| Cholesterol metabolism regulation. | Activation of pathways of liver metabolism. | Decrease low-density lipoprotein cholesterol concentration. | [137] |
Lycopene also modulates oxidative stress by regulating the expression of antioxidant genes such as superoxide dismutase, catalase (CAT), and glutathione peroxidase (GPx) that minimize the amount of free radicals in the body [23]. Similarly, lycopene inhibits the activation of nuclear factor kappa B, a key regulator of inflammatory response and oxidative stress. By modulating this pathway, lycopene contributes to the reduction of inflammatory processes and the maintenance of cellular homeostasis; that is, it promotes the silencing of proinflammatory genes and promotes apoptosis in cancer cells [20]. It also inhibits insulin-like growth factor 1 (IGF-1) binding to its receptor by increasing IGF-binding protein-1 concentrations, interfering with IGF-1 signaling, and reducing the activation of the phosphatidylinositol 3-kinase and RAS-mitogen-activated protein kinase pathways that limit cell proliferation and promote apoptosis [138].
In prostate cancer cells, lycopene decreases the expression of androgen receptors and modulates the nuclear localization of β-catenin, thereby reducing cell proliferation and cell cycle progression. Likewise, lycopene is associated with the activation of the peroxisome proliferator-activated receptor gamma-activated receptor [127,139], liver X receptor alpha, and ABCA1 transporter pathway, promoting a reduction in cholesterol concentration in prostate cancer cells, thereby reducing their proliferation [137]. These mechanisms highlight the potential of lycopene in the prevention and treatment of diseases such as cancer, especially prostate cancer, and underline the importance of its inclusion in the diet for health promotion. However, the capacity of lycopene can be altered by handling and processing fruits and vegetables during food processing or by the conditions used for its extraction and industrial use.
6. EPIDEMIOLOGICAL AND CLINICAL INSIGHTS
Lycopene has gained significant attention in epidemiological research and clinical studies due to its potential health benefits, particularly in age-related diseases and oral health. Although the current evidence suggests that lycopene may contribute to the management of various health conditions, further research is necessary to confirm its therapeutic potential.
6.1. Epidemiological Evidence
Lycopene-rich diets have been associated with a reduced risk of chronic diseases, particularly those related to aging [140]. As mentioned above, this may be due, in part, to its high antioxidant and anti-inflammatory capacity. However, caution should be exercised in this interpretation, as lifestyle, diet, and metabolism can have a positive or negative impact on this response.
6.1.1. Epidemiological studies
Epidemiological studies have shown an inverse relationship between the intake of lycopene-rich foods and the risk of cardiovascular disease. One study showed that those with a higher lycopene intake had a lower risk of developing cardiovascular disease (up to 30%). It has also been suggested that lycopene may inhibit cholesterol synthesis by reducing the activity of the enzyme 3-hydroxy-3-methylglutaryl-CoA reductase, a key factor in the production of cholesterol in the body. This effect could contribute to lower blood cholesterol levels and thus lower the risk of cardiovascular diseases [14]. In addition, published epidemiological literature suggests an interaction between study design and the relationship between lycopene (or tomato consumption) and lung cancer risk.
6.1.2. Observational studies
These studies have further indicated that higher lycopene intake correlates with lower incidences of certain cancers and cardiovascular diseases, although the outcomes vary depending on the studied population and dietary patterns [141]. It was recently demonstrated that lycopene, as a natural antioxidant agent and thanks to its anti-inflammatory effect, protects against kidney damage by reducing oxidative stress and minimizing the incidence of Chronic Kidney Disease and mortality from this disease. Surprisingly, there is a negative correlation between the incidence of kidney cancer, cisplatin-induced nephrotoxic damage, and lycopene consumption [142].
6.1.3. Cohort studies
Cohort studies generally do not show a significant association, whereas case–control studies indicate a decreased risk of greater lycopene and tomato consumption. Likewise, [143] a prospective cohort study (n = 139,913) reported that high consumption of tomatoes and lycopene was associated with lower mortality from cardiovascular damage. The main pathway carried out by this carotenoid is the regulation of oxidative stress originating from the positive responses mentioned above, which are derived from alterations in 255 regulatory genes [142].
Given that epidemiological studies are used to establish associations between factors and health, this type of study can, in turn, be observational or cohort-based. In the first case, as they are inexpensive and quick, they allow lycopene consumption to be described in relation to the incidence of risks to human health. However, they tend to generalize the response, and there are variables that can mask the real impact of lycopene. In contrast, cohort studies, by selecting a specific population, allow us to understand and infer the potential benefits of lycopene consumption in the diet. However, there may be some changes in the participants’ lifestyle, suggesting alterations in the results. In this sense, the benefits of lycopene consumption should be considered with caution.
6.1.4. Systematic review
In a systematic review and dose-response meta-analysis of prospective cohort studies, Balali et al. [24] found that high lycopene intake and high blood levels are associated with a reduced overall risk of cancer (prostate, lung); in particular, an increase of 10 g/dL of lycopene in the blood resulted in a lower risk (5%). The authors also reported a lower cancer-related death rate owing to lycopene. This positive response may be due to the inhibition of IGF-1 and angiogenesis by suppressing receptors and transporters that trigger abnormal cell growth [24]. Similarly, high tomato intake (as a source of lycopene) is associated with a lower risk of death in adults with diabetes and cancer [144].
It is important to note that the bioavailability and bioaccessibility of lycopene, its metabolism, and the participants’ intake levels and lifestyle can significantly influence the results of these studies, since high consumption of lycopene in matrices that provide stable isomers and are accompanied by fat intake can translate into systemic effects in humans.
6.2. Clinical Trials
Currently, 773 clinical trials are registered in the Cochrane Library, an international network dedicated to producing systematic reviews that support informed health decisions. These trials focus on a wide range of conditions, from general health maintenance to specific diseases, such as prostate cancer, infertility, periodontitis, and cellulitis, as well as investigations into lycopene pharmacokinetics. However, a significant portion of these trials did not publish their results [145].
In a multicenter, randomized, double-blind, phase II pilot study in patients with metastatic colorectal cancer treated with panitumumab, it was found that the administration of lactolycopene (20 mg·100/g of food/matrix) induced a reduction in grade 2–3 panitumumab skin toxicity in 41% of the patients, which was due to the reduction in malondialdehyde production, as well as by the replenishment of antioxidant consumption, a response mediated by the hydrophobic property of lactolycopene itself, which showed an increase in the plasma of the patients [146] [Figure 6]. Therefore, studies have been conducted to define their safety and possible toxicological impacts, which are discussed in the following section.
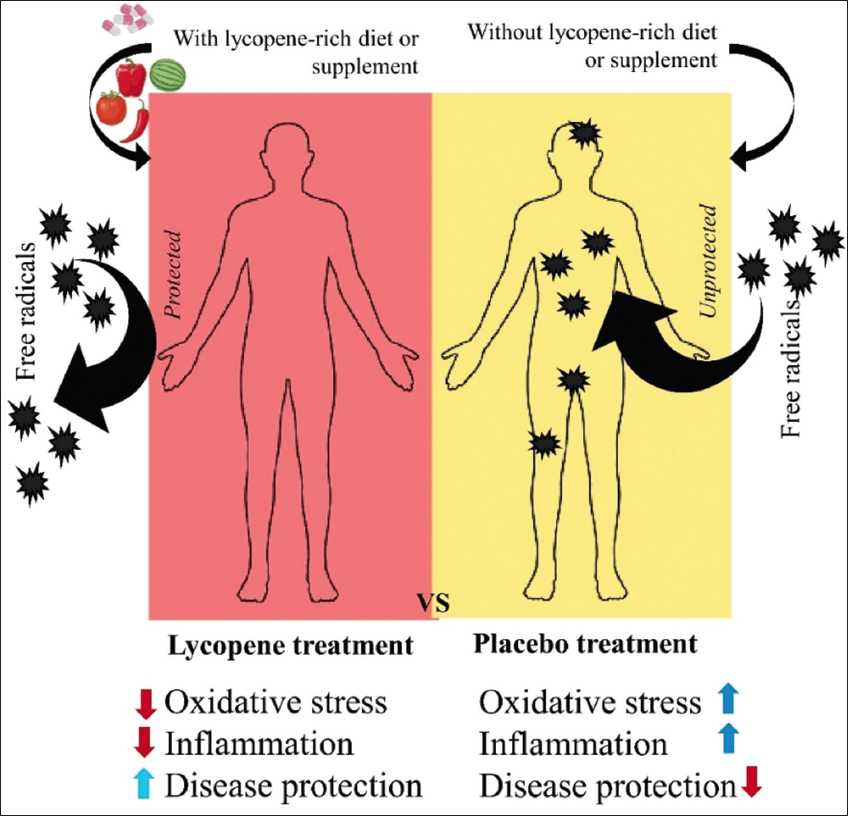 | Figure 6: Potential response to lycopene use in humans in clinical trials. Figure our own, created using Microsoft Office PowerPoint (Microsoft Corporation, Redmond, Washington, USA). [Click here to view] |
Furthermore, the design of clinical trials (i.e., age cohort, lycopene dosage and sources, and duration) may yield contradictory data due to diversity in human metabolism and lycopene itself. Therefore, future studies should consider a nutritional “stabilization” phase that minimizes noise derived from dietary habits and maximizes the potential of lycopene. Similarly, clinical trials may choose to develop specific cohorts of children, young adults, middle-aged adults, and the elderly, as well as comprehensive approaches to the main pathophysiologies. We suggest conducting studies (meta-analyses) that include, in addition to the bioavailability of lycopene, clinical assessment parameters to clarify which variables respond most or best to interventions with this antioxidant.
Overall, the data show a positive impact between lycopene consumption and the reduction of certain specific diseases. However, this causal response must be interpreted holistically and consider parameters such as actual lycopene intake, bioavailability (which, as mentioned above, involves a knowledge gap in itself), time of intake, and the consumption of lycopene “enhancers” (fatty acids) or “limiters” (fiber). We believe that the human-environment-lifestyle (epigenome) interaction may be a parameter that epidemiological studies and clinical trials should strongly consider as a determinant of the favorable response to lycopene, since this interaction may influence how lycopene promotes health.
6.3. Safety and Toxicological Considerations
Lycopene has been extensively studied for its safety as a food additive and dietary supplement. It is currently authorized as a food additive (color) in the European Union under Annex II of Regulation (EC) No 1333/2008, allowing its use in several food categories. The Scientific Panel on food additives, flavorings, processing aids, and materials in contact with food (AFC) evaluated its toxicological data and established an acceptable daily intake (ADI) of 0.5 mg/kg body weight per day, applying a safety factor of 100 [147]; this value corresponds to a combined uncertainty factor (uncertainty factor of 10 times × factor of 10 times relating to interindividual variability in humans) that considers subpopulations at higher risk due to certain factors such as nutritional status, medication use, diet, pregnancy status, and other pathophysiology.
However, safety concerns have been raised due to the possibility that the highest estimated intakes for both children and adults may exceed the established ADI of 0.5 mg/kg body weight [148]. Despite these concerns, most notifications submitted by companies to the European Chemicals Agency through CLP notifications indicate that lycopene is not hazardous [149]. Reproductive and teratological studies support the safety of lycopene. In a teratology study in Wistar rats, lycopene formulations showed no maternal toxicity or teratogenic effect. The only observation was a slight increase in the incidence of extra-thoracic ribs in the treated groups compared with the controls, with no other relevant changes [23].
Several studies have evaluated the safety and toxicity of lycopene and concluded that its extract from natural tomato oleoresin is safe for human consumption. For instance, a no-observed-adverse-effect level of 4500 mg/kg/day was established, which significantly exceeded the typical dietary intake with no significant toxicity reported in previous studies [150]. In addition, tomato oleoresin rich in lycopene Z-isomers (10.9% lycopene, with 66.3% Z-isomer content) demonstrated no mutagenic potential or adverse effects in toxicity tests, including an LD50 >5000 mg/kg body weight in rats [151].
Biddle et al. [152] report that intervention with a daily intake of 29.4 mg of lycopene (through a 330 mL serving of 100% vegetable juice V8 for 30 days) in participants with heart failure promoted plasma lycopene levels up to 0.76 μmol/L, making it safe for consumption. Similarly, consumption of 11 mg of lycopene per day (raw tomatoes at breakfast and lunch) for 8 weeks in overweight postmenopausal women (aged 45–70) with a body mass index >24 kg/m2 was not only safe but also showed a significant response in reducing the risks of metabolic syndrome [153].
6.4. Lycopene-drug Interactions
Research suggests that lycopene can enhance the therapeutic effects of certain medications. For example, it has been shown to significantly reduce blood glucose levels in diabetic models when combined with gliclazide without altering its pharmacokinetics [154]. In addition, the antioxidant properties of lycopene may help mitigate the toxic side effects of chemotherapeutic agents, potentially offering protective benefits during cancer treatment [155].
Lycopene’s effects on drug metabolism further highlight the need for caution in certain contexts. It exhibits a mixed inhibition mechanism for ponatinib metabolism in both rat and human liver microsomes, indicating potential interactions in patients with chronic myeloid leukemia [156]. In addition, lycopene has a moderate inhibitory effect on CYP2E1, suggesting that its co-administration with drugs metabolized by this enzyme requires careful monitoring [157].
A pre-clinical study in men with metastatic prostate cancer (phase I trial: NCT0149519) showed that lycopene enhanced the efficacy of docetaxel against prostate cancer when treated with 21-day cycles of 75 mg/m2 docetaxel/and androgen deprivation therapy plus lycopene (30, 90, or 150 mg/day). This suggests that the combination has low toxicity and favorable pharmacokinetics [158].
6.5. Recommended Daily Intake
There is an ongoing debate regarding the amount of lycopene needed to achieve its functional benefits, as studies have reported highly variable results, making comparisons and clear recommendations challenging. Discrepancies among epidemiological, in vitro, and in vivo studies, along with lycopene’s lack of provitamin A activity, contribute to it not being classified as an essential nutrient, thus preventing the establishment of an official RDI. However, some suggestions include consuming 7–10 servings of lycopene-rich food, whereas others recommend consuming 5–35 mg/day. The European Food Safety Authority has set an acceptable ADI of 0.5 mg/day. Since the bioavailability of lycopene depends on processing, fat intake, and the isomeric form of the carotenoid, it is recommended to select the appropriate food matrix (fresh or processed, rich in cis-isomers) and accompany lycopene consumption with fats to achieve the RDI. In this context, the sources of lycopene in humans vary, ranging from fruits and vegetables to dyes and additives. The latter is economically important for the food industry.
7. INDUSTRIAL APPLICATIONS
Among the industrial uses of lycopene, three major categories can be listed, including its use as a natural colorant and antioxidant in foods, its use in dietary supplements, and its use in pharmaceuticals and cosmetics [Table 3]. In all cases, there is a high demand for this molecule, which is why the conceptual association between biosynthesis → environmental modulation → industrial uses is included here. Below, we will briefly delve into each of the sections mentioned.
Table 3: Main industrial uses of lycopene.
| Applications | Benefits | Limitations |
|---|---|---|
| Natural Colorant and Antioxidant in Food | Provides nutraceutical characteristics that are beneficial to the food matrix. High probability of bioavailability. Reduces oxidation of food products by scavenging free radicals. | Molecular stabilization is highly dependent on extraction temperature. Light and pH affect the ratio of isomers in the food matrix. |
| Nutraceuticals and Dietary Supplements | Protective effects against disease. Dosage and intake are known. | Stability of lycopene in tablets. High demand for pure lycopene from fruits and vegetables. Manufacturing costs of supplements. |
| Pharmaceutical and Cosmetic | High antioxidant and anti-inflammatory capacity. Reduces stress caused by ultraviolet radiation and air pollution. Promotes antioxidant enzymes. | Limited extraction systems and methods. Molecule stabilization. |
7.1. Stability and Modification of the Lycopene Content in Food
Owing to its highly unsaturated structure, lycopene is prone to chemical reactions [28] by thermal energy, oxygen, pH, light, and metals, causing a change in its configuration through isomerization [37,38]. Isomerization occurs as a degradation step because cis-isomers are less stable and can cleave more easily than trans-isomers [159]. The stability of lycopene configurations was in the order of all-trans > 9-cis > 13-cis > 15-cis. Lycopene degradation varies depending on fruit and vegetable matrices. For example, lycopene degradation differs widely among tomato peel, puree, and orange matrices [160].
Temperature can accelerate lycopene degradation; in tomatoes crushed at 70°C, 80°C, 90°C, and 100°C for 120 min, lycopene degradation was 23.94%, 30.17%, 45.05%, and 55.24%, respectively [28]. Likewise, the type of fruit extraction (hot air, vacuum drying, and microwave vacuum drying) conditions the degradation of lycopene; the higher the temperature, the greater the degradation, following the first law of degradation kinetics [161]. These data suggest that processing temperature promotes lycopene instability [28].
Light promotes lycopene oxidation, and its isomerization depends on the bond and isomer. Thus, the protection of fruits and vegetables from light could decrease the oxidative degradation and isomerization of lycopene, apparently because cis-isomers are significantly reduced by exposure to light, revealing their instability with respect to all-trans isomers [28].
Considering that the optimal pH range for stabilizing lycopene is 3.5–4.5, probably the alterations in these values may promote cis-bond formation [37]. Similarly, oxygen, which presents an unpaired electron, can cause lycopene degradation because of its high reactivity. Similarly, metal ions (FeCl3) can induce oxidative degradation by exerting catalytic effects on lycopene Z-isomerization [28]. These changes caused by processing, by altering the proportion of isomers, promote oxidation and degradation of the food matrix, which results in reduced or limited bioavailability of lycopene for humans, mainly due to the alteration of the nature of the molecule.
In this sense, proposals have been made to improve lycopene stability, such as encapsulation systems that, in addition to promoting a physical barrier, exhibit solubility [28]. Among these, emulsions, microemulsions, nanoemulsions, nanostructured lipid carriers, hydrogels, Pickering emulsions, and liposomes are notable [162]. In this context, an emulsion loaded with Z-lycopene using ovalbumin-chitosan complexes showed improved carotenoid stability under UV irradiation [39]. Pickering emulsions stabilized by complexes of whey protein isolate fibrils and sodium alginate offer better protection of lycopene against thermal degradation and light [163]. Thus, food processing had a significant influence on lycopene concentrations in the food matrices [Table 4].
Table 4: Lycopene content according to processing.
| Source | Processing or extraction method | Lycopene content (mg·100/g) | References |
|---|---|---|---|
| Fresh tomato pulp | Unprocessed | 16.11–20.45 | [164] |
| Tomato paste | Unprocessed | 184.29–211.73 | |
| Tomato pulp | Dried | 19.30–25.11 | |
| Tomato | Solvent extraction | 22.00 | [165] |
In Psidium guajava L., the freeze-drying dehydration method (low temperature and pressure) allows for higher lycopene conservation (7.66 mg·100–1 dry weight [DW]) compared to hot air and vacuum drying (4.36 mg·100/g DW) [166]. Similar to thermochemical treatment (tomato pulp heated at 80–82°C for 30 min before applying 0.05% potassium metabisulfite, 0.05% sodium metabisulfite, 0.05% potassium sorbate, and 0. 05% sodium benzoate, 0.5% ascorbic acid, and citric acid (1%) promote higher lycopene conservation (4.328–5.987 mg·100/g FW) over 120 days when stored at 30 ± 1°C [167].
Some tomato products, such as sauces and pastes, tend to have a higher lycopene content than fresh tomatoes. Studies report that tomato sauces contain between 9.9 and 13.4 mg·100/g of lycopene, whereas fresh tomato has between 0.88 and 7.74 mg·100/g. It should be noted that dietary sources of lycopene are mainly in the isomeric trans-form [122].
However, the extraction method is crucial for determining the amount of lycopene extracted and recovered [1]. Extraction methods, such as vapor entrainment and organic solvents, showed variations in lycopene extraction efficiency, and solvents such as ethyl acetate showed high lycopene extraction efficiency. Recently, some enzyme-assisted methods from tomato tissues and peels have shown higher yields using cellulases and pectinases, up to 18-fold in lycopene recovery. However, the hexane: acetone:ethanol mixture (50:25:25) optimized the recovery of the compound. Likewise, surfactants have been used to extract lycopene from various sources (watermelon, pink grapefruit, guava, melon, fresh red tomatoes, and papaya) with a recovery of up to 25% lycopene [168]. Taken together, these data suggest a high variability in lycopene content depending on the type of processing and extraction method employed.
As an antioxidant biomolecule, lycopene can enter the human diet through fresh or processed foods; however, in the latter, lycopene content depends on the source, processing, and extraction method. Therefore, consumption of foods rich in lycopene is not necessarily associated with an increase in this antioxidant in humans. To understand and elucidate the benefits of lycopene in humans, in-depth studies have been conducted on animals and humans to understand the impact of this molecule on biology and its potential impact on organisms.
7.2. Lycopene as a Natural Colorant and Antioxidant in the Food Industry
Lycopene offers significant potential as a colorant in various food products due to its vibrant red hue and associated health benefits. As a natural colorant, lycopene aligns with the growing consumer preference for natural ingredients over synthetic alternatives, making it an attractive option for food manufacturers.
7.2.1. Stability and bioavailability
Tomato-based products, such as sauces, pastes, and juices, are the primary sources of lycopene in the diet. The processing of tomatoes enhances the bioaccessibility of lycopene, as heat and mechanical processing break down cell walls and release more lycopene [164]. The antioxidant properties of lycopene are particularly noteworthy, as they help protect food products from oxidative damage, thereby extending shelf life and maintaining quality [1]. This capability is crucial in the food industry, where the preservation of flavor and nutritional value is paramount. Moreover, the effectiveness of lycopene as a colorant is enhanced when consumed along with dietary fats, further increasing its bioavailability and health benefits. Its strong color and non-toxicity make it a suitable alternative to artificial colorants, which are increasingly being scrutinized by health-conscious consumers [Figure 7]. In addition, lycopene stability during processing and storage is a critical factor that supports its use in food products, as it remains effective for extended periods [38].
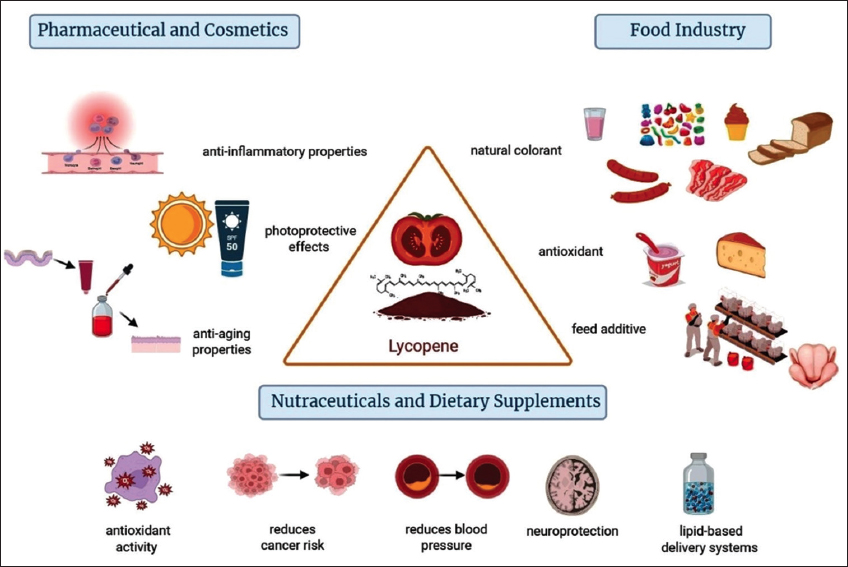 | Figure 7: Main industrial application of lycopene. Figure our own, created with BioRender (biorender.com). [Click here to view] |
7.2.2. Meat products
Lycopene has been shown to improve the color parameters of meat products, increasing redness and overall color saturation, which are desirable attributes for consumer acceptance. This effect is particularly notable in products such as sausages and turkey breasts, where lycopene addition enhances visual appeal [169]. Similarly, lycopene has been employed in emulsified meat products to enhance color stability and reduce oxidative rancidity, and the incorporation of lycopene into meat emulsions, such as pork and mutton, has been shown to increase surface redness (a* value), which is a measure of red color intensity [1].
Recent studies have investigated the replacement of sodium nitrite with lycopene pigments in meat products, particularly sausages. One study evaluated the substitution of sodium nitrite with lycopene pigment in sausages containing 40% meat. The findings indicated that after 30 days of storage, there were no significant differences in physicochemical properties, microbiological tests, lightness index (L*), or sensory attributes between the treated and control samples. These results suggest that lycopene could serve as a viable alternative to sodium nitrite in sausage formulations [29]. Another study examined the incorporation of tomato paste, a rich source of lycopene, into Frankfurt-type sausages. This study found that higher levels of tomato paste enhanced consumer preference owing to improved color attributes. Moreover, reducing sodium nitrite levels while incorporating tomato paste did not negatively affect the product quality. These findings support the potential of lycopene as a natural alternative to synthetic additives such as sodium nitrite in processed meat products, offering both coloring properties and potential health benefits [170].
In animal production, the use of lycopene as a feed additive has been shown to enhance the antioxidant capacity, leading to improved meat quality. In poultry production, lycopene supplementation has been associated with improved growth performance, particularly in broilers exposed to aflatoxin B1, by enhancing antioxidant capacity and reducing oxidative stress [171]. In addition to increasing antioxidant content and providing nutritional benefits, lycopene can improve meat texture and flavor. Furthermore, as a carotenoid, it contributes to a more appealing meat color, which consumers often associate with freshness and quality [33].
7.2.3. Dairy products
Studies have demonstrated its successful incorporation into cheese. Here, we evaluated the substitution of butter with lycopene oil in processed cheese and assessed its effects on physicochemical and sensory properties. The results demonstrated improvements in dry matter, ash content, lycopene concentration, antioxidant activity, nutritional profile, and meltability, and enhanced sensory attributes and consumer appeal [30].
Tomato peel powder (TPP) is a rich source of lycopene, which serves as a natural colorant and antioxidant in yogurt. This incorporation enhances the redness and antioxidant capacity of the product. For instance, yogurt containing 8% TPP exhibited the highest redness value and increased radical-scavenging activity [31]. TPP also enriches yogurt with Vitamin C, thereby improving the nutritional profile. Sensory evaluation studies have shown a high consumer acceptability of yogurt with 0.1% TPP incorporation [172].
7.2.4. Bakery products
The incorporation of lycopene as a colorant in bread enhances its nutritional profile by increasing antioxidant capacity [173]. Tomato powder, a natural source of lycopene, also contains essential vitamins and minerals, which further improve the overall nutritional value of bread [32]. The stability of lycopene during storage is a crucial factor, with amylose-lycopene complexes demonstrating good stability, though their degradation rates are influenced by temperature. In addition, the inclusion of lycopene-rich ingredients, such as tomato powder, can enhance sensory attributes such as color and flavor, which are key to consumer acceptance [173]. In addition to its nutritional and sensory benefits, lycopene also affects dough’s physical properties, impacting extensibility and fermentation, which in turn influences the final texture and volume of bread [174].
7.2.5. Packaging applications
In addition to its role as a colorant, lycopene exhibits potent antioxidant properties, effectively neutralizes free radicals, and prevents oxidative damage [175]. This characteristic is particularly beneficial for extending the shelf life of perishable food. Moreover, studies have explored the incorporation of lycopene into packaging materials to enhance food preservation by creating active packaging systems that inhibit oxidation and microbial contamination [176]. Lycopene not only provides functional benefits but also promotes health, this makes it a valuable ingredient in health-oriented food products.
Changes resulting from processing on the color and antioxidant activity of lycopene alter the isomer ratios and stability of the molecule, which can potentially affect the bioaccessibility/bioavailability of lycopene in humans.
7.3. Applications in Nutraceuticals and Dietary Supplements
Lycopene has been playing a dual role as a dietary supplement and nutraceutical [Figure 6]. Its properties have been linked to various health benefits, including potential protective effects against chronic diseases.
Its health-promoting properties have been extensively studied owing to its capacity as a nutraceutical. Its antioxidant activity is instrumental in neutralizing free radicals, thereby mitigating oxidative stress, which is a key contributor to various chronic diseases [137]. In addition, lycopene has been associated with cardiovascular benefits such as improved lipid profiles and reduced blood pressure, which may collectively decrease the risk of atherosclerosis and hypertension [2]. Moreover, emerging research indicates that lycopene may play a role in neuroprotection, potentially offering benefits in conditions such as depression by enhancing synaptic plasticity [2].
As a dietary supplement, lycopene is available in various formulations, such as capsules and tablets, aimed at augmenting daily intake, particularly in individuals with insufficient dietary consumption [137]. The bioavailability of lycopene is notably influenced by its formulation and the presence of dietary fats, by improving its absorption up to ~4.4 times more than a fat-free diet [120]. Studies have demonstrated that lycopene incorporated into lipid-based delivery systems such as oil suspensions or emulsions exhibits improved bioavailability. For instance, lycopene dissolved in olive oil (20 mL) has shown higher absorption rates than lycopene dissolved in other media [120]. This enhanced bioavailability is crucial for maximizing the physiological benefits.
7.4. Development of Pharmaceutical and Cosmetic Applications
Lycopene has attracted considerable attention for its pharmaceutical potential due to its strong antioxidant and anti-inflammatory properties. Its ability to neutralize free radicals helps prevent cellular damage and reduces the risk of chronic diseases, making it a promising candidate for various medical applications. Studies have suggested that lycopene may contribute to cardiovascular health by reducing oxidative stress and inflammation, thereby lowering blood pressure and improving lipid profiles, which can mitigate the risk of atherosclerosis, heart attack, and stroke [177]. In cancer prevention, antioxidant activity plays a crucial role in preventing DNA damage and inhibiting tumor growth, with research indicating its potential to induce apoptosis in cancer cells [178]. In addition, the photoprotective effects of lycopene have been explored to reduce UV-induced skin damage and potentially lower the risk of skin cancer. Emerging evidence also highlights its neuroprotective properties, suggesting its benefits in neurodegenerative diseases such as Alzheimer’s and Parkinson’s, as it may reduce oxidative stress and inflammation in the brain [179]. These diverse therapeutic applications underscore the significance of lycopene in pharmaceutical research; however, further studies are required to optimize its bioavailability and therapeutic efficacy.
In the cosmetic industry, lycopene and other carotenoids are highly valued for their skin-protective and anti-aging properties. Lycopene’s potent antioxidant activity helps shield the skin from environmental stressors such as UV radiation and pollution, thereby preventing premature aging. Commonly incorporated into serum, creams, and sunscreens [Figure 6], lycopene contributes to skin rejuvenation and protects against UV-induced damage; for instance, lycopene-infused facial creams are marketed to reduce fine lines and wrinkles while enhancing skin elasticity and overall appearance; mainly by significantly reducing the parameters associated with skin damage (Δa*, MMP-1, ICAM-19) associated with a significant increase in minimum erythema dose, skin thickness, and skin density [35]. Similarly, carotenoids serve as natural colorants, imparting hues from yellow to orange without relying on synthetic additives [180] and combating oxidative stress by strengthening the endogenous antioxidant defenses of the skin, including enzymes such as CAT and GPx, as well as non-enzymatic elements such as L-ascorbic acid and α-tocopherol [34]. Moreover, clinical trials have confirmed their effectiveness in reducing the signs of premature aging, and their ability to absorb UV light and neutralize free radicals (mainly 1O2) offers robust photoprotection against sun-induced damage [181].
The data observed reveal that lycopene extraction and handling techniques and methods are a fundamental challenge in the industrial sector and represent a key element that must be implemented and developed to promote greater optimization, efficiency, and effectiveness. However, achieving optimal stability of this molecule on an industrial scale suggests a major challenge involving the concatenation of theoretical knowledge and, perhaps, the development of machines and new methods, for example, using artificial intelligence and/or mathematical algorithms.
8. OUTLOOKS, FUTURE DIRECTIONS, AND RESEARCH GAPS
Lycopene is a biomolecule with high antioxidant potential that can enter the human diet through fruits and vegetables or processed foods. However, the synthesis of this carotenoid is dependent on the environmental factors in which the plants develop. Therefore, it is necessary to establish mechanisms and management practices to promote fruit and vegetable production. Therefore, there are still great challenges in the management and promotion of alternative crops that diversify the sources of lycopene in people’s diets beyond tomatoes.
Another challenge is that lycopene intake in humans is usually very low or even zero; therefore, strategies must be implemented to increase the content of this biomolecule (biofortification) to increase its consumption. For example, targeted applications of nanotechnologies, mineral nutrients, and/or precursor molecules for lycopene biosynthesis, as well as the implementation of genomic techniques in crops such as tomatoes and/or chili peppers, may be simple, accessible, and practical strategies for promoting lycopene content in plants. However, it is also essential to improve the food matrix to release a greater amount of this compound (bioaccessibility) into the digestive tract, This can be achieved through biofortification, which can increase the content of vitamin C and fatty acids. Similarly, the processing methods for lycopene sources have a significant impact on its content (bioavailability). The use of nanoencapsulation, emulsifiers, and new technologies can be used as tools to improve the intake of this carotenoid.
Therefore, it is imperative to consider industrial and post-harvest handling. In this regard, emerging areas such as nanotechnology and omics could serve as tools for enhancing the stability and content of lycopene in fruits, vegetables, and processed foods. In addition, the food industry requires more comprehensive studies to expand the use of lycopene as a natural additive (colorant) to improve the appearance, texture, and flavor of ultra-processed products (meats, sausages, and yogurt). Finally, owing to its quality, lycopene has gained relevance in skin care; however, further studies are needed.
There are relatively few clinical studies and pharmacological evidence developed between lycopene consumption and various diseases; therefore, the potential of this carotenoid is not yet fully clear and known, requiring large-scale studies that consider its interaction with various drugs that people commonly consume. In addition, discrepancies in epidemiological, in vitro, and in vivo studies must be addressed to standardize the plausible effects of lycopene. Similarly, because the consumption of this carotenoid is scarce in most populations, it is complicated to design, implement, and investigate the positive association between its intake and human health. Although evidence reveals a plausible effect on its consumption and reduction of oxidative stress in humans, these areas of study need to be strengthened.
Similarly, it is recommended that the food industry establish direct links with producers of lycopene-rich crops to increase the efficiency of lycopene recovery, as well as to study and implement efficient and accessible extraction methods and develop affordable products enriched with lycopene. Similarly, in order to integrate the health benefits of lycopene consumption, it is essential that the health sector (doctors and nutritionists) make recommendations for diets rich in this carotenoid. Finally, it is necessary to implement public policies that permeate society. Eventually, all these recommendations will allow for the practical relevance of lycopene consumption and its benefits.
9. CONCLUSION
Given its diverse biological roles and wide range of applications, lycopene stands out as a compound of great interest in both scientific research and industry. Its natural origin, strong antioxidant capacity, and relevance in health promotion make it a promising target for further development in agriculture, nutrition, and biotechnology. Continued interdisciplinary research is essential to fully harness its potential to improve human health and enhance functional product development. This molecule has a promising future in the health sector (mainly in cardiovascular health and disease prevention), in industry (as an antioxidant), in the food sector (as a food additive), and in the development of functional foods enriched with lycopene. Therefore, at the research level, new studies should focus on elucidating the impact of lycopene on humans, as well as its interaction with lifestyles and its benefits in most of the diseases that afflict society. Meanwhile, the industry needs to strengthen its efforts to efficiently extract lycopene and/or stabilize it within the food matrix.
10. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
11. FUNDING
There is no funding for this report.
12. CONFLICT OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
13. ETHICAL APPROVAL
This study does not involve experiments on animals or human subjects.
14. DATA AVAILABILITY
All data generated and analyzed are included within this research article.
15. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors, or the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
16. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Li Z, Yu F. Recent advances in lycopene for food preservation and shelf-life extension. Foods. 2023;12:3121. [CrossRef]
2. Joung HR, Gang G, Go G. Lycopene and cardiovascular health: A systematic review reveals promising benefits in blood pressure, lipid profiles, and oxidative stress. Food Eng Prog. 2023;27:271-7. [CrossRef]
3. Nunes L, Hashemi N, Gregersen SB, Tavares GM, Corredig M. Compartmentalization of lutein in simple and double emulsions containing protein nanoparticles: Effects on stability and bioaccessibility. Food Res Int. 2023;173:113404. [CrossRef]
4. Saini RK, Ranjit A, Sharma K, Prasad P, Shang X, Gowda KGM, et al. Bioactive compounds of Citrus fruits:A review of composition and health benefits of carotenoids, flavonoids, limonoids, and terpenes. Antioxidants (Basel). 2022;11:239. [CrossRef]
5. Astija A, Febriani VI, Alibasyah L, Isnainar I. Climate and cultivar of tomato (Licopersicum esculentum L.) affect the lycopene contents. Arch Pharm Pract. 2023;14:39-43. [CrossRef]
6. Hashem A, Bayoumi Y, El-Zawily AES, Tester M, Rakha M. Interspecific hybrid rootstocks improve productivity of tomato grown under high-temperature stress. HortScience. 2024;59:129-37. [CrossRef]
7. Preciado-Rangel P, Rueda-Puente EO, Valdez-Aguilar LA, Reyes- Pérez JJ, Gallegos-Robles MA, Murillo Amador B. Electrical conductivity of nutrient solution and their effect on bioactive compounds and yield of bell pepper (Capsicum annuum L.). Trop Subtrop Agroecosyst. 2021;24:12. [CrossRef]
8. Krupa-Ma?kiewicz M, Ochmian I. Sustainable approaches to alleviate heavy metal stress in tomatoes: Exploring the role of chitosan and nanosilver. Agronomy. 2024;14:2477. [CrossRef]
9. PatanèC, Pellegrino A, Saita A, Calcagno S, Cosentino SL, Scandurra A, et al. A study on the effect of biostimulant application on yield and quality of tomato under long-lasting water stress conditions. Heliyon. 2025;11:e41187. [CrossRef]
10. Petrozza A, Summerer S, Melfi D, Mango T, Vurro F, Bettelli M, et al. A lycopene ε-cyclase TILLING allele enhances lycopene and carotenoid content in fruit and improves drought stress tolerance in tomato plants. Genes (Basel). 2023;14:1284. [CrossRef]
11. Wang Y, Tian C, Na Q, Zhu C, Cao H, Zhang M, et al. The role of SlCHRC in carotenoid biosynthesis and plastid development in tomato fruit. Int J Biol Macromol. 2024;281:136354. [CrossRef]
12. Wu H, Wu Y, Cui Z, Hu L. Nutraceutical delivery systems to improve the bioaccessibility and bioavailability of lycopene: A review. Crit Rev Food Sci Nutr. 2024;64:6361-79. [CrossRef]
13. Sharifi-Rad J, Rodrigues CF, Sharopov F, Docea AO, Can Karaca A, Sharifi-Rad M, et al. Diet, lifestyle and cardiovascular diseases: Linking pathophysiology to cardioprotective effects of natural bioactive compounds. Int J Environ Res Public Health. 2020;17:2326. [CrossRef]
14. Przybylska S, Tokarczyk G. Lycopene in the prevention of cardiovascular diseases. Int J Mol Sci. 2022;23:1957. [CrossRef]
15. Khan UM, Sevindik M, Zarrabi A, Nami M, Ozdemir B, Kaplan DN, et al. Lycopene: Food sources, biological activities, and human health benefits. Oxid Med Cell Longev. 2021;2021:2713511. [CrossRef]
16. Rakha SI, Ateya AI, Safhi FA, Abdellatif AM. Ameliorative effect of lycopene on follicular reserve depletion, oxidative damage, apoptosis rate, and hormonal profile during repeated superovulations in mice. Vet Sci. 2024;11:414. [CrossRef]
17. Meng Q, Zhang Y, Li J, Shi B, Ma Q, Shan A. Lycopene affects intestinal barrier function and the gut microbiota in weaned piglets via antioxidant signaling regulation. J Nutr. 2022;152:2396-408. [CrossRef]
18. Adeyeye TA, Shallie OF, Saula TR, Afolabi SA, Shallie PD. Neuroprotective effects of lycopene on lipopolysaccharide-induced cerebellar damage in rats: Implication for Alzheimer's disease therapy. J Exp Clin Anat. 2024;21:346-57. [CrossRef]
19. Waisundara V. Antioxidants - Benefits, Sources, Mechanisms of Action. London, United Kingdom:IntechOpen;2021. [CrossRef]
20. Kulawik A, Cielecka-Piontek J, Czerny B, Kami?ski A, Zalewski P. The relationship between lycopene and metabolic diseases. Nutrients. 2024;16:3708. [CrossRef]
21. Murotomi K, Umeno A, Shichiri M, Tanito M, Yoshida Y. Significance of singlet oxygen molecule in pathologies. Int J Mol Sci. 2023;24:2739. [CrossRef]
22. Lakshminarayana R, Paul B. Free radical chemistry of carotenoids and oxidative stress physiology of cancer. In: Handbook of Oxidative Stress in Cancer: Therapeutic Aspects. Singapore:Springer Singapore;2022. 1-22. [CrossRef]
23. Ibrahim IM, Althagafy HS, Abd-Alhameed EK, Al-Thubiani WS, Hassanein EHM. Promising hepatoprotective effects of lycopene in different liver diseases. Life Sci. 2022;310:121131. [CrossRef]
24. Balali A, Fathzadeh K, Askari G, Sadeghi O. Dietary intake of tomato and lycopene, blood levels of lycopene, and risk of total and specific cancers in adults:A systematic review and dose-response meta-analysis of prospective cohort studies. Front Nutr. 2025;12:1516048. [CrossRef]
25. Hariprasanna K, Chetankumar B, Venkateswarlu R, Niharika G. Approaches for enhancing the nutrients bioavailability. In: Tonapi VA, Talwar HS, Are AK, Bhat BV, Reddy CR, Dalton TJ, editors. Sorghum 21st Century Food - Fodd.-Feed-Fuel a Rapidly Chang. World, Singapore:Springer Singapore;2020. 809-35. [CrossRef]
26. Shahidi F, Pan Y. Influence of food matrix and food processing on the chemical interaction and bioaccessibility of dietary phytochemicals:A review. Crit Rev Food Sci Nutr. 2022;62:6421-45. [CrossRef]
27. Sharma I, Khare N, Rai A. Carotenoids: Sources, Bioavailability and their Role in Human Nutrition. Diet Carotenoids - Sources, Probiotics Play a Crucial Role in Human Health. London:IntechOpen;2024. [CrossRef]
28. Bakhshizadeh M, Moghaddam TN, Tavassoli M, Mousavi Khaneghah A, Ansarifar E. Characterization, extraction, and encapsulation technologies of lycopene and applications in functional food products: An updated review. Food Bioprocess Technol. 2024;18:3059-99. [CrossRef]
29. Nateghi L, Zarei F, Afshari KP. The effect of sodium nitrite replacement with lycopene pigment in german sausage and evaluation of its physicochemical, antimicrobial and sensory properties. J Nutr Food Secur. 2024;9:265-74. [CrossRef]
30. Bakry A. Novel processed cheese production using lycopene oil obtained by green-extraction technique of tomato peels waste. Nutr Food Process. 2021;4:1-10. [CrossRef]
31. Meegahawaththa W, Sinhalage D, Mudannayake D. Tomato (Lycopersicon esculentum L.) peel powder as a source of natural antioxidant and a colorant in stirred yoghurt. Food Life. 2020;2020:87-97. [CrossRef]
32. Ramya HN, Kandkur S, Ashwini A, Bai RSR. Utilization of tomato powder as a technological functional ingredient in bakery products:A review. Int J Plant Soil Sci. 2023;35:157-61. [CrossRef]
33. Long Y, Paengkoum S, Lu S, Niu X, Thongpea S, Taethaisong N, et al. Physicochemical properties, mechanism of action of lycopene and its application in poultry and ruminant production. Front Vet Sci. 2024;11:1364589. [CrossRef]
34. Bin-Jumah M, Alwakeel SS, Moga M, Buvnariu L, Bigiu N, Zia-Ul-Haq M. Application of carotenoids in cosmetics. In:Carotenoids:Structure and Function in the Human Body. Cham:Springer International Publishing;2021, 747-56. [CrossRef]
35. Zhang X, Zhou Q, Qi Y, Chen X, Deng J, Zhang Y, et al. The effect of tomato and lycopene on clinical characteristics and molecular markers of UV-induced skin deterioration:A systematic review and meta-analysis of intervention trials. Crit Rev Food Sci Nutr. 2024;64:6198-217. [CrossRef]
36. Huang JJ, Xu W, Lin S, Cheung PCK. The bioactivities and biotechnological production approaches of carotenoids derived from microalgae and cyanobacteria. Crit Rev Biotechnol. 2024;45(2):276-304. [CrossRef]
37. Caseiro M, Ascenso A, Costa A, Creagh-Flynn J, Johnson M, Simões S. Lycopene in human health. LWT. 2020;127:109323. [CrossRef]
38. Tufail T, Bader Ul Ain H, Noreen S, Ikram A, Arshad MT, Abdullahi MA. Nutritional benefits of lycopene and beta? carotene: A comprehensive overview. Food Sci Nutr. 2024;12:8715-41. [CrossRef]
39. Wang L, Jin N, Xie Y, Zhu W, Yang Y, Wang J, et al. Improvements in the appearance and nutritional quality of tomato fruits resulting from foliar spraying with silicon. Foods. 2024;13:223. [CrossRef]
40. Wang H, Lin Y, Liu Q, Zhou A, Bian H, Zhang W, et al. Antioxidant, anticancer activity and molecular docking study of lycopene with different ratios of Z-isomers. Curr Res Food Sci. 2023;6:100455. [CrossRef]
41. Kössler S, Armarego-Marriott T, TarkowskáD, Ture?kováV, Agrawal S, Mi J, et al. Lycopene β-cyclase expression influences plant physiology, development, and metabolism in tobacco plants. J Exp Bot. 2021;72:2544-69. [CrossRef]
42. Syawalluddin NS, Abdul Rahman H, Lim SJ, Wan Mustapha WA, Mohd Razali NS, Kasim KF, et al. Lycopene and β-carotene thermal degradation kinetics and colour-antioxidant changes in gac (Momordica cochinchinensis) fruit aril paste. Int J Food Sci Technol. 2024;59:7808-17. [CrossRef]
43. Buaban P, Beckles DM, Mongkolporn O, Luengwilai K. Lycopene accumulation in pummelo (Citrus Maxima [burm.] merr.) is influenced by growing temperature. Int J Fruit Sci. 2020;20:149-63. [CrossRef]
44. Asaduzzaman M, Hasan N, Begum K, Hoque SMZ. Degradation kinetics of lycopene from red amaranth &preparation of winter melon jelly using this lycopene and comparison with commercial jelly. Heliyon. 2024;10:e31135. [CrossRef]
45. Majumder S, Mishra N, Vikrant. Edible flowers of India as alternate source of high quantity of lycopene. Vegetos. 2023;37:433-7. [CrossRef]
46. Martínez-Ispizua E, Martínez-Cuenca MR, Marsal JI, Díez MJ, Soler S, Valcárcel JV, et al. Bioactive compounds and antioxidant capacity of valencian pepper landraces. Molecules. 2021;26:1031. [CrossRef]
47. Flores-Velazquez J, Mendoza-Perez C, Rubiños-Panta JE, Ruelas-Islas JR. Quality and yield of bell pepper cultivated with two and three stems in a modern agriculture system. Horticulturae. 2022;8:1187. [CrossRef]
48. Saini RK, Prasad P, Lokesh V, Shang X, Shin J, Keum YS, et al. Carotenoids:Dietary sources, extraction, encapsulation, bioavailability, and health benefits-a review of recent advancements. Antioxidants. 2022;11:795. [CrossRef]
49. Silva KG, Cavalcanti MT, Martinsa LP, Alves RC, Lucena FA, Santos MSA, et al. Coatings based on gelatin and chitosan in the conservation of papaya (Carica papaya L.) minimally processed. Horticulturae. 2023;9:729. [CrossRef]
50. Kumarasinghe HS, Kim JH, Kim SL, Kim KC, Perera RMTD, Kim SC, et al. Bioactive constituents from Carica papaya fruit:Implications for drug discovery and pharmacological applications. Appl Biol Chem. 2024;67:103. [CrossRef]
51. Nadeem M, Navida M, Ameer K, Iqbal A, Malik F, Nadeem MA, et al. A comprehensive review on the watermelon phytochemical profile and their bioactive and therapeutic effects. Korean J Food Preserv. 2022;29:546-76. [CrossRef]
52. Chen L, Chen QY, Huang LB, Yang MY, Cen YJ, Xu CB, et al. Carotenoid gene expression and metabolic changes in the pericarp of Mandarin 'Shatangju'under net shed shading during growth. J Food Compos Anal. 2024;126:105862. [CrossRef]
53. Carmona L, Alquézar B, Peña L. Biochemical characterization of new sweet orange mutants rich in lycopene and β-carotene antioxidants. Antioxidants. 2024;13:994. [CrossRef]
54. Vanessa BG, Alice TI, William DA, Adelaide DM, Gisèle LE, Condurache NN, et al. Effect of fertilizers on yield, phytochemical, and antioxidant properties of Cucurbita moschata fruits. Food Sci Nutr. 2024;12:6742-51. [CrossRef]
55. Wang T, Hou Y, Hu H, Wang C, Zhang W, Li H, et al. Functional validation of phytoene synthase and lycopene ?-cyclase genes for high lycopene content in autumn olive fruit (Elaeagnus umbellata). J Agric Food Chem. 2020;68:11503-11. [CrossRef]
56. Paiva YF, Figueirêdo RMF, Queiroz AJM, Amadeu LTS, Santos FS, Reis CG, et al. Physicochemical aspects, bioactive compounds, phenolic profile and in vitro antioxidant activity of tropical red fruits and their blend. Molecules 2023;28:4866. [CrossRef]
57. Chumroenphat T, Bunyatratchata A, Siriamornpun S. Under-utilized wild fruit Lepisanthes rubiginosa (Roxb.) Leenh:A discovery of novel lycopene and anthocyanin source and bioactive compound profile changes associated with drying conditions. Dry Technol. 2023;41:2297-308. [CrossRef]
58. Zhou W, Niu Y, Ding X, Zhao S, Li Y, Fan G, et al. Analysis of carotenoid content and diversity in apricots (Prunus armeniaca L.) grown in China. Food Chem. 2020;330:127223. [CrossRef]
59. Imran M, Ghorat F, Ul-Haq I, Ur-Rehman H, Aslam F, Heydari M, et al. Lycopene as a natural antioxidant used to prevent human health disorders. Antioxidants. 2020;9:706. [CrossRef]
60. Kaur H, Ghosh M. Probiotic fermentation enhances bioaccessibility of lycopene, polyphenols and antioxidant capacity of guava fruit (Psidium guajava L). J Agric Food Res. 2023;14:100704. [CrossRef]
61. Dhatwalia J, Kumari A, Guleria I, Kumar Shukla R, Saleh N, El-Nashar HAS, et al. Effect of altitude and harvest year on nutraceutical characteristics of Rubus ellipticus fruits. Front Nutr. 2024;11:1419862. [CrossRef]
62. Hamdi D, Hafidi A, Lemaire JJ, Messaoud C. A comparative study of secondary metabolites profiling and biological activity of Smyrnium olusatrum L. leaf, flower and fruit. Nat Prod Res. 2024;39(13):3806-3820. [CrossRef]
63. Martínez JP, Fuentes R, Farías K, Lizana C, Alfaro JF, Fuentes L, et al. Effects of salt stress on fruit antioxidant capacity of wild (Solanum chilense) and domesticated (Solanum lycopersicum var. cerasiforme) tomatoes. Agronomy. 2020;10:1481. [CrossRef]
64. Takács S, Pék Z, Csányi D, Daood HG, Szuvandzsiev P, Palotás G, et al. Influence of water stress levels on the yield and lycopene content of tomato. Water (Switzerland). 2020;12:2165. [CrossRef]
65. Francesca S, Cirillo V, Raimondi G, Maggio A, Barone A, Rigano MM. A novel protein hydrolysate-based biostimulant improves tomato performances under drought stress. Plants. 2021;10:783. [CrossRef]
66. PatanèC, Siah S, Pellegrino A, Cosentino SL, Siracusa L. Fruit yield, polyphenols, and carotenoids in long shelf-life tomatoes in response to drought stress and rewatering. Agronomy. 2021;11:1943. [CrossRef]
67. Suboti?A, Jevremovi?S, Miloševi?S, Trifunovi?-Mom?ilov M, ?uri?M, Koruga ?. Physiological response, oxidative stress assessment and aquaporin genes expression of cherry tomato (Solanum lycopersicum L.) exposed to hyper-harmonized fullerene water complex. Plants (Basel). 2022;11:2810. [CrossRef]
68. Bianchi AR, Vitale E, Guerretti V, Palumbo G, De Clemente IM, Vitale L, et al. Antioxidant characterization of six tomato cultivars and derived products destined for human consumption. Antioxidants. 2023;12:761. [CrossRef]
69. Romdhane A, Riahi A, Ujj A, Ramos-Diaz F, Marjanovi?J, Hdider C. Comparative nutrient and antioxidant profile of high lycopene variety with hp genes and ordinary variety of tomato under organic conditions. Agronomy. 2023;13:649. [CrossRef]
70. Igbakaa DN, Girgih AT, Ahure D, Ejoha PO. Bioactive and sensory properties of sweet Italian (Capsicum annuum) and habanero (Capsicum chinense) pepper commonly consumed in benue state of Nigeria. Asian J Biotechnol Bioresour Technol. 2023;9:1-9. [CrossRef]
71. Ezekiel A, Abiodun O, Funmilayo AD, Tolulope IO, Oluwatosin O, Ogola-Emma E. Identification of ascorbic acid and phytochemical profiles of Scotch Bonnet pepper (Capsicum chinense) using high-performance liquid chromatography-ultraviolet detector (HPLC-UV). J Appl Nat Sci. 2023;15:1475-83. [CrossRef]
72. Jiang Q, Ye J, Zhu K, Wu F, Chai L, Xu Q, et al. Transcriptome and co-expression network analyses provide insights into fruit shading that enhances carotenoid accumulation in pomelo (Citrus grandis). Hortic Plant J. 2022;8:423-34. [CrossRef]
73. Wang YH, Zhang YQ, Zhang RR, Zhuang FY, Liu H, Xu ZS, et al. Lycopene ε-cyclase mediated transition of α-carotene and β-carotene metabolic flow in carrot fleshy root. Plant J. 2023;115:986-1003. [CrossRef]
74. Karata?F, Mukhtar ZG, Ibrahim MS, Ibrahim YI. Amounts of vitamin A, vitamin E, vitamin C, β-carotene, lycopene, ghrelin, glutathione, and mda in fruits of diospyros kaki L. G?da. 2019;44:585-92. [CrossRef]
75. Bakar B, Çakmak M, Ibrahim MS, Özer D, Saydam S, Karatas F. Investigation of amounts of vitamins, lycopene, and elements in the fruits of Opuntia ficus-indica subjected to different pretreatments. Biol Trace Elem Res. 2020;198:315-23. [CrossRef]
76. Goztepe B, Kayacan S, Bozkurt F, Tomas M, Sagdic O, Karasu S. Drying kinetics, total bioactive compounds, antioxidant activity, phenolic profile, lycopene and β-carotene content and color quality of Rosehip dehydrated by different methods. LWT-Food Sci Technol. 2022;153:112476. [CrossRef]
77. González-Moscoso M, Martínez-Villegas NV, Cadenas-Pliego G, Benavides-Mendoza A, Del Carmen Rivera-Cruz M, González-Morales S, et al. Impact of silicon nanoparticles on the antioxidant compounds of tomato fruits stressed by arsenic. Foods. 2019;8:612. [CrossRef]
78. Li B, Shukla MK, Du T. Combined environmental stresses induced by drip irrigation positively affect most solar greenhouse grown tomato fruit quality. Sci Hortic (Amsterdam). 2021;288:110334. [CrossRef]
79. Tan S, Ke Z, Chai D, Miao Y, Luo K, Li W. Lycopene, polyphenols and antioxidant activities of three characteristic tomato cultivars subjected to two drying methods. Food Chem. 2021;338:128062. [CrossRef]
80. Vega-López B, Carvajal-Miranda Y, Brenes-Peralta L, Gamboa-Murillo M, Venegas-Padilla J, Rodríguez G, et al. Phytonutraceutical evaluation of five varieties of tomato (Solanum lycopersicum) during ripening and processing. LWT-Food Sci Technol. 2022;164:113592. [CrossRef]
81. Rouen A, Baghizadeh A, Roghami M, Mousavi A. Physiological and biochemical responses of watermelon landraces to water deficit stress. Russ J Plant Physiol. 2023;70:1-13. [CrossRef]
82. He Y, Wang J, Yang J, Bai P, Feng J, Wu Y, et al. Enhancement of tomato fruit quality through moderate water deficit. Foods. 2024;13:3540. [CrossRef]
83. Li X, Wang X, Zhang Y, Zhang A, You CX. Regulation of fleshy fruit ripening: from transcription factors to epigenetic modifications. Hortic Res. 2022;9:uhac013. [CrossRef]
84. Quian-Ulloa R, Stange C. Carotenoid biosynthesis and plastid development in plants: The role of light. Int J Mol Sci. 2021;22:1184. [CrossRef]
85. Ziaei N, Talebi M, Sayed Tabatabaei BE, Sabzalian MR, Soleimani M. Intra-canopy LED lighting outperformed top LED lighting in improving tomato yield and expression of the genes responsible for lycopene, phytoene and vitamin C synthesis. Sci Rep. 2024;14:19043. [CrossRef]
86. Lee SY, Jang SJ, Jeong HB, Lee SY, Venkatesh J, Lee JH, et al. A mutation in Zeaxanthin epoxidase contributes to orange coloration and alters carotenoid contents in pepper fruit (Capsicum annuum). Plant J. 2021;106:1692-707. [CrossRef]
87. Liu Z, Dai Y, Mao L, Yang S, Yang B, Sun Y, et al. Integrated metabolome and transcriptome analysis revealed carotenoid metabolism difference in pepper (Capsicum annuum L.) yellowing mutants under different light quality. Veg Res. 2022;2:21. [CrossRef]
88. Panjai L, Röhlen-Schmittgen S, Ellenberger J, Noga G, Hunsche M, Fiebig A. Effect of postharvest irradiation with red light on epidermal color and carotenoid concentration in different parts of tomatoes. J Food Meas Charact. 2021;15:1737-46. [CrossRef]
89. Conti V, Romi M, Guarnieri M, Cantini C, Cai G. Italian tomato cultivars under drought stress show different content of bioactives in pulp and peel of fruits. Foods. 2022;11:270. [CrossRef]
90. Ni K, Lu X, Li S, Li F, Zhang Y, Cui R, et al. GhLCYε-3 characterized as a lycopene cyclase gene responding to drought stress in cotton. Comput Struct Biotechnol J. 2024;23:384-95. [CrossRef]
91. Rabbi RHM, Aktar N, Mahamud MA, Paul NC, Halder D, Imran S. Impact of different zinc concentrations on growth, yield, fruit quality, and nutrient acquisition traits of tomato (Lycopersicon esculentum L.) grown under salinity stress. Arch Biol Sci. 2024;76:71-82. [CrossRef]
92. Christou A, Georgiadou EC, Zissimos AM, Christoforou IC, Christofi C, Neocleous D, et al. Uptake of hexavalent chromium by tomato (Solanum lycopersicum L.) plants and mediated effects on their physiology and productivity, along with fruit quality and safety. Environ Exp Bot. 2021;189:104564. [CrossRef]
93. Dasgan HY, Dere S, Akhoundnejad Y, Arpaci BB. Effects of high-temperature stress during plant cultivation on tomato (Solanum lycopersicum L.) fruit nutrient content. J Food Qual. 2021;2021:7994417. [CrossRef]
94. Singh R, Saffeullah P, Umar S, Abass S, Ahmad S, Iqbal N. Comparing the individual and combined effects of nano zinc and conventional zinc fertilization on growth, yield, phytochemical properties, antioxidant activity, and secoisolariciresinol diglucoside content in linseed. Plant Nano Biol. 2024;10:100098. [CrossRef]
95. Mesa T, Polo J, Arabia A, Caselles V, Munné-Bosch S. Differential physiological response to heat and cold stress of tomato plants and its implication on fruit quality. J Plant Physiol. 2022;268:153581. [CrossRef]
96. Li L, Wang X, Lv J, Duan W, Huang T, Zhao K, et al. Overexpression of Sly-miR167a delayed postharvest chilling injury of tomato fruit under low temperature storage. Postharvest Biol Technol. 2023;204:112420. [CrossRef]
97. Lafuente MT, Sampedro R, Vélez D, Romero P. Deficient copper availability on organoleptic and nutritional quality of tomato fruit. Plant Sci. 2023;326:111537. [CrossRef]
98. Karem MH, Al-Abedy AN, Kadhim JH. Effect of Tomato yellow leaf curl virus (TYLCV) Infection of some Tomato (Solanum lycopersicom L.) genotypes on fruits content of lycopene and some vitamins. IOP Conf Ser Earth Environ Sci. 2023;1259:012092. [CrossRef]
99. Botella MÁ, Hernández V, Mestre T, Hellín P, García-Legaz MF, Rivero RM, et al. Bioactive compounds of tomato fruit in response to salinity, heat and their combination. Agriculture. 2021;11:534. [CrossRef]
100. Cardarelli M, Ceccarelli AV, El Nakhel C, Rouphael Y, Salehi H, Ganugi P, et al. Foliar applications of a Malvaceae-derived protein hydrolysate and its fractions differentially modulate yield and functional traits of tomato under optimal and suboptimal nitrogen application. J Sci Food Agric. 2024;104:7603-16. [CrossRef]
101. Luta G, Balan D, Stanca M, Jerca O, Jurcoane S, Niculescu M, et al. Innovative protein gel treatments to improve the quality of tomato fruit. Gels. 2024;10:10. [CrossRef]
102. Chanthini KMP, Senthil-Nathan S, Pavithra GS, Asahel AS, Malarvizhi P, Murugan P, et al. The macroalgal biostimulant improves the functional quality of tomato fruits produced from plants grown under salt stress. Agriculture. 2023;13:6. [CrossRef]
103. Elkelish A, Ibrahim MFM, Ashour H, Bondok A, Mukherjee S, Aftab T, et al. Exogenous application of nitric oxide mitigates water stress and reduces natural viral disease incidence of tomato plants subjected to deficit irrigation. Agronomy. 2021;11:87. [CrossRef]
104. Wang J, Zhang J, Li J, Dawuda MM, Ali B, Wu Y, et al. Exogenous application of 5-aminolevulinic acid promotes coloration and improves the quality of tomato fruit by regulating carotenoid metabolism. Front Plant Sci. 2021;12:683868. [CrossRef]
105. Wang J, Yuan H, Wu Y, Yu J, Ali B, Zhang J, et al. Application of 5-aminolevulinic acid promotes ripening and accumulation of primary and secondary metabolites in postharvest tomato fruit. Front Nutr. 2022;9:1036843. [CrossRef]
106. Chen X, Man X, Huang D, Fang H, Yang R, Liao W. Hydrogen sulfide inhibits tomato fruit ripening by restraining fruit softening and ethylene production. LWT-Food Sci Technol. 2024;200:116210. [CrossRef]
107. Fernandes Â, Chaski C, Pereira C, Kosti?M, Rouphael Y, Sokovi?M, et al. Water stress alleviation effects of biostimulants on greenhouse-grown tomato fruit. Horticulturae. 2022;8:645. [CrossRef]
108. Ahmad A, Javad S, Iqbal S, Shahid T, Naz S, Shah AA, et al. Efficacy of soil drench and foliar application of iron nanoparticles on the growth and physiology of Solanum lycopersicum L. exposed to cadmium stress. Sci Rep. 2024;14:27920. [CrossRef]
109. Hanif S, Farooq S, Kiani MZ, Zia M. Surface modified ZnO NPs by betaine and proline build up tomato plants against drought stress and increase fruit nutritional quality. Chemosphere. 2024;362:142671. [CrossRef]
110. Ahmad A, Javad S, Iqbal S, Shahzadi K, Gatasheh MK, Javed T. Alleviation potential of green-synthesized selenium nanoparticles for cadmium stress in Solanum lycopersicum L:Modulation of secondary metabolites and physiochemical attributes. Plant Cell Rep. 2024;43:113. [CrossRef]
111. Duduit JR, Kosentka PZ, Miller MA, Blanco-Ulate B, Lenucci MS, Panthee DR, et al. Coordinated transcriptional regulation of the carotenoid biosynthesis contributes to fruit lycopene content in high-lycopene tomato genotypes. Hortic Res. 2022;9:uhac084. [CrossRef]
112. Wan X, Wang Z, Duan W, Huang T, Song H, Xu X. Knockdown of Sly-miR164a enhanced plant salt tolerance and improved preharvest and postharvest fruit nutrition of tomato. Int J Mol Sci. 2023;24:4639. [CrossRef]
113. Yin Z, Liu J, Zhao H, Chu X, Liu H, Ding X, et al. SlMYB1 regulates the accumulation of lycopene, fruit shape, and resistance to Botrytis cinerea in tomato. Hortic Res. 2023;10:uhac282. [CrossRef]
114. Grundy MML, Moughan PJ, Wilde PJ. Bioaccessibility and associated concepts:Need for a consensus. Trends Food Sci Technol. 2024;145:104373. [CrossRef]
115. Lu Y, Mu K, McClements DJ, Liang X, Liu X, Liu F. Fermentation of tomato juice improves in vitro bioaccessibility of lycopene. J Funct Foods. 2020;71:104020. [CrossRef]
116. Kulawik A, Wdowiak K, Kulawik M, Rosiak N, Paczkowska-Walendowska M, Cielecka-Piontek J, et al. Application of hot-melt extrusion in modifying the solubility of lycopene. Appl Sci. 2026;16(1):17. [CrossRef]
117. Carpentieri S, Ferrari G, DonsìF. High-pressure homogenization for enhanced Bioactive recovery from tomato processing by-products and improved lycopene bioaccessibility during in vitro digestion. Antioxidants (Basel). 2023;12(10):1855. [CrossRef]
118. Liang X, Yan J, Guo S, McClements DJ, Ma C, Liu X, et al. Enhancing lycopene stability and bioaccessibility in homogenized tomato pulp using emulsion design principles. Innov Food Sci Emerg Technol. 2021;67:102525. [CrossRef]
119. Ntzimani A, Tsevdou M, Katsouli M, Thanou I, Tsimogiannis D, Giannakourou M, et al. Recovery of carotenoids via novel extraction technologies for the valorization of tomato by-products. Processes. 2025;13(9):2964. [CrossRef]
120. Arballo J, Amengual J, Erdman JW. Lycopene:A critical review of digestion, absorption, metabolism, and excretion. Antioxidants (Basel). 2021;10:342. [CrossRef]
121. Przybylska S. Lycopene –a bioactive carotenoid offering multiple health benefits:A review. Int J Food Sci Technol. 2020;55:11-32. [CrossRef]
122. Walallawita US, Wolber FM, Ziv-Gal A, Kruger MC, Heyes JA. Potential role of lycopene in the prevention of postmenopausal bone loss:Evidence from molecular to clinical studies. Int J Mol Sci. 2020;21:7119. [CrossRef]
123. Marzocco S, Singla RK, Capasso A. Multifaceted effects of lycopene:A boulevard to the multitarget-based treatment for cancer. Molecules. 2021;26:5333. [CrossRef]
124. Ratto F, Franchini F, Musicco M, Caruso G, Di Santo SG. A narrative review on the potential of tomato and lycopene for the prevention of Alzheimer's disease and other dementias. Crit Rev Food Sci Nutr. 2022;62:4970-81. [CrossRef]
125. Meléndez-Martínez AJ, Stinco CM, Mapelli-Brahm P. Skin carotenoids in public health and nutricosmetics:The emerging roles and applications of the UV radiation-absorbing colourless carotenoids phytoene and phytofluene. Nutrients. 2019;11:1093. [CrossRef]
126. Mustra Rakic J, Wang XD. Role of lycopene in smoke-promoted chronic obstructive pulmonary disease and lung carcinogenesis. Arch Biochem Biophys. 2020;689:108439. [CrossRef]
127. Saini RK, Rengasamy KRR, Mahomoodally FM, Keum YS. Protective effects of lycopene in cancer, cardiovascular, and neurodegenerative diseases:An update on epidemiological and mechanistic perspectives. Pharmacol Res. 2020;155:104730. [CrossRef]
128. Rejali L, Ozumerzifon S, Nayeri H, Asgary S. Risk reduction and prevention of cardiovascular diseases:Biological mechanisms of lycopene. Bioact Compd Health Dis. 2022;5:202. [CrossRef]
129. Upadhyay RK. High cholesterol disorders, myocardial infarction and its therapeutics. World J Cardiovasc Dis. 2023;13:433-69. [CrossRef]
130. Mannino F, D'Angelo T, Pallio G, Ieni A, Pirrotta I, Giorgi DA, et al. The nutraceutical genistein-lycopene combination improves bone damage induced by glucocorticoids by stimulating the osteoblast formation process. Nutrients. 2022;14:4296. [CrossRef]
131. Marcucci G, Domazetovic V, Nediani C, Ruzzolini J, Favre C, Brandi ML. Oxidative stress and natural antioxidants in osteoporosis:Novel preventive and therapeutic approaches. Antioxidants (Basel). 2023;12:373. [CrossRef]
132. Ashraf W, Latif A, Lianfu Z, Jian Z, Chenqiang W, Rehman A, et al. Technological advancement in the processing of lycopene:A review. Food Rev Int. 2022;38:857-83. [CrossRef]
133. Guo Z, Liu Y, Luo Y. Mechanisms of carotenoid intestinal absorption and the regulation of dietary lipids:Lipid transporter-mediated transintestinal epithelial pathways. Crit Rev Food Sci Nutr. 2024;64:1791-816. [CrossRef]
134. Rippin HL, Maximova K, Loyola E, Breda J, Wickramasinghe K, Ferreira-Borges C, et al. Suboptimal intake of fruits and vegetables in nine selected countries of the World Health Organization European Region. Prev Chronic Dis. 2023;20:230159. [CrossRef]
135. Varela E, Gomes A, da Silva Barbosa dos Santos A, de Carvalho E, Vale V, Percário S. Potential benefits of lycopene consumption:rationale for using it as an adjuvant treatment for malaria patients and in several diseases. Nutrients. 2022;14:5303. [CrossRef]
136. Khalaf RA, Awad M. Lycopene as a potential bioactive compound:Chemistry, extraction, and anticancer prospective. Curr Cancer Drug Targets. 2023;23:634-42. [CrossRef]
137. Shafe MO, Gumede NM, Nyakudya TT, Chivandi E. Lycopene:A potent antioxidant with multiple health benefits. J Nutr Metab. 2024;2024:6252426. [CrossRef]
138. Crudele L, Piccinin E, Moschetta A. Visceral adiposity and cancer:Role in pathogenesis and prognosis. Nutrients. 2021;13:2101. [CrossRef]
139. Mirahmadi M, Azimi-Hashemi S, Saburi E, Kamali H, Pishbin M, Hadizadeh F. Potential inhibitory effect of lycopene on prostate cancer. Biomed Pharmacother. 2020;129:110459. [CrossRef]
140. Abir MH, Mahamud AGMSU, Tonny SH, Anu MS, Hossain KHS, Protic IA, et al. Pharmacological potentials of lycopene against aging and aging?related disorders:A review. Food Sci Nutr. 2023;11:5701-35. [CrossRef]
141. Zamani M, Behmanesh Nia F, Ghaedi K, Mohammadpour S, Amirani N, Goudarzi K, et al. The effects of lycopene and tomato consumption on cardiovascular risk factors in adults:A grade assessment systematic review and meta-analysis. Curr Pharm Des. 2023;29:1671-700. [CrossRef]
142. Pan X, Zhu R, Pei J, Zhang L. Lycopene:A potent antioxidant to alleviate kidney disease. Int Immunopharmacol. 2025;151:114363. [CrossRef]
143. Jung H, Shin S. Association between tomato and lycopene consumption and risk of all-cause and cause-specific mortality: Korean genome and epidemiology study cohort. Mol Nutr Food Res. 2023;67:2200452. [CrossRef]
144. Liu Y, Liu H, Liu J, Liu W, Yang Y, Liu Y. Associations between dietary intake of tomato and lycopene with all-cause and cancer-specific mortality in US adults with diabetes: Results from a cohort study. Nutr Cancer. 2024;76:974-84. [CrossRef]
145. Cochrane Library;2025. Available from: https://www.cochranelibrary.com/search [Last accessed on 2025 Feb 17]
146. Moroni M, Pirovano M, Brugnatelli S, Zucca M, Morreale M, Rizzo V, et al. Lycopene minimizes skin toxicity and oxidative stress in patients treated with panitumumab-containing therapy for metastatic colorectal cancer. J Funct Foods. 2021;83:104533. [CrossRef]
147. López-Rodríguez R, Vermetten J, Domínguez L, Fernández-Ruiz V, Cámara M. A critical review of synthetic novel foods within the European regulation:Proposed classification, toxicological concerns and potential health claims. Crit Rev Food Sci Nutr. 2025;65:6688-708. [CrossRef]
148. Turck D, Bohn T, Castenmiller J, De Henauw S, Hirsch?Ernst KI, Maciuk A, et al. Safety of yellow/orange tomato extract as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2023;21:7994. [CrossRef]
149. (ECHA) TECA. Substance Infocard;2025. Available from: https://echa.europa.eu/substance-information/-/substanceinfo/100.007.227 [Last accessed on 2025 April 22]
150. Matulka RA, Hood AM, Griffiths JC. Safety evaluation of a natural tomato oleoresin extract derived from food-processing tomatoes. Regul Toxicol Pharmacol. 2004;39:390-402. [CrossRef]
151. Honda M, Higashiura T, Fukaya T. Safety assessment of a natural tomato oleoresin containing high amounts of Z?isomers of lycopene prepared with supercritical carbon dioxide. J Sci Food Agric. 2017;97:1027-33. [CrossRef]
152. Biddle MJ, Lennie TA, Bricker GV, Kopec RE, Schwartz SJ, Moser DK. Lycopene dietary intervention:A pilot study in patients with heart failure. J Cardiovasc Nurs. 2015;30(3):205-12. [CrossRef]
153. Chen CY, Chien YW. Fresh tomato (Lycopersicon esculentum Mill.) in the Diet improves the features of the metabolic syndrome:A randomized study in postmenopausal women. Biology. 2024;13(8):588. [CrossRef]
154. Nugroho H, Sianturi M, Retnoningrum D, Susiloningsih N. Lycopene enhances the beta cell capacity and antihyperlipidemic effects of metformin on type 2 diabetic rats. Indones Biomed J. 2024;16:208-91. [CrossRef]
155. Puah BP, Jalil J, Attiq A, Kamisah Y. New insights into molecular mechanism behind anti?cancer activities of lycopene. Molecules. 2021;26:3888. [CrossRef]
156. Chen J, Hong F, Xia H, Shen Y, Chen X, Wu H, et al. Investigation of ponatinib metabolism and drug-drug interactions with lycopene and shikonin in vitro and in vivo. Chem Biol Interact. 2024;404:111265. [CrossRef]
157. Li M, Tang S, Peng X, Sharma G, Yin S, Hao Z, et al. Lycopene as a therapeutic agent against aflatoxin B1-related toxicity:Mechanistic insights and future directions. Antioxidants (Basel). 2024;13:452. [CrossRef]
158. Lilly MB, Wu C, Ke Y, Chen WP, Soloff AC, Armeson K, et al. A phase I study of docetaxel plus synthetic lycopene in metastatic prostate cancer patients. Clin Transl Med. 2024;14(3):e1627. [CrossRef]
159. Tahmasebi M, Emam-Djomeh Z. Lycopene degradation and color characteristics of fresh and processed tomatoes under the different drying methods:A comparative study. Chem Pap. 2021;75:3617-23. [CrossRef]
160. Shi K, Hu T, Zhang P, Zhang S, Xu Y, Zhang Z, et al. Thermal conditions and active substance stability affect the isomerization and degradation of lycopene. Food Res Int. 2022;162:111987. [CrossRef]
161. Sonawane A, Chauhan OP, Semwal SD, Semwal AD. Drying characteristics and lycopene degradation kinetics of tomato soup. Chem Data Collect. 2021;35:100757. [CrossRef]
162. Liang L, Zhu J, Zhang Z, Liu Y, Wen C, Liu X, et al. Pickering emulsion stabilized by tea seed cake protein nanoparticles as lutein carrier. Foods. 2022;11:1712. [CrossRef]
163. Zhu Q, Qiu Y, Zhang L, Lu W, Pan Y, Liu X, et al. Encapsulation of lycopene in Pickering emulsion stabilized by complexes of whey protein isolate fibrils and sodium alginate:Physicochemical property, structural characterization and in vitro digestion property. Food Res Int. 2024;191:114675. [CrossRef]
164. Górecka D, Wawrzyniak A, J?drusek-Goli?ska A, Dziedzic K, Hamu?ka J, Kowalczewski P?, et al. Lycopene in tomatoes and tomato products. Open Chem. 2020;18:752-6. [CrossRef]
165. Rowles III, JL. The role of tomato and lycopene consumption on prostate carcinogenesis and prostate cancer treatment. 2020. (Doctoral dissertation, University of Illinois at Urbana-Champaign). University of Illinois Library. https://hdl.handle.net/2142/107997.
166. Rakariyatham K, Boonyapranai K, Laokuldilok T, Utama-ang N, Nutprem A, Kaewprasit K, et al. Impact of different dehydration methods on physicochemical and functional properties of guava (Psidium guajava L.) powder prepared from white and pink pomaces. Appl Food Res. 2025;5:100696. [CrossRef]
167. Kabir MS, Roy U, Suvo SP, Sobhan A, Kamal MM, Akhter MJ, et al. Comparative assessment of fresh and processed tomato (Solanum lycopersicum) pulps:Impact of processing on physicochemical, antioxidant, and enzymatic behavior. Appl Food Res. 2024;4:100550. [CrossRef]
168. Sánchez M, Laca A, Laca A, Díaz M. Value?added products from fruit and vegetable wastes: A review. Clean Soil Air Water. 2021;49:2000376. [CrossRef]
169. Mikami N, Yamashiro Y, Nagaoka S, Akamatsu Y, Maeda H, Shimada K. Effects of different tomato products on the lipid oxidation, color, and texture of uncured cooked pork sausages in storage. Food Sci Technol Res. 2023;29:FSTR-D-23-00016. [CrossRef]
170. Stoica M, Antohi VM, Alexe P, Ivan AS, Stanciu S, Stoica D, et al. New strategies for the total/partial replacement of conventional sodium nitrite in meat products: A review. Food Bioprocess Technol. 2022;15:514-38. [CrossRef]
171. Ma H, Chen Q, Yang H, Wan X. Effects of lycopene on the growth performance, meat quality, and antioxidant capacity of broiler chickens challenged with aflatoxin B 1. J Food Sci. 2024;89:96-103. [CrossRef]
172. Jefrin JA, Kamaleshraj RS, Kumar AP, Gandhi R. Development of yogurt enriched with freeze-dried tomato peel powder. Int J Eng Appl Sci Technol. 2022;7:192-8. [CrossRef]
173. Raczyk M, Kruszewski B, Zachariasz E. Effect of tomato, beetroot and carrot juice addition on physicochemical, antioxidant and texture properties of wheat bread. Antioxidants. 2022;11:2178. [CrossRef]
174. Vartolomei N, Turtoi M. Effect of rosehip powder addition on dough extensographic, amylographic and rheofermentographic properties and sensory attributes of bread. Processes. 2023;11:1088. [CrossRef]
175. Kulawik A, Cielecka-Piontek J, Zalewski P. The importance of antioxidant activity for the health-promoting effect of lycopene. Nutrients. 2023;15:3821. [CrossRef]
176. Roy S, Deshmukh RK, Tripathi S, Gaikwad KK, Das SS, Sharma D. Recent advances in the carotenoids added to food packaging films: A review. Foods. 2023;12:4011. [CrossRef]
177. Tsiantoulas D, Eslami M, Obermayer G, Clement M, Smeets D, Mayer FJ, et al. APRIL limits atherosclerosis by binding to heparan sulfate proteoglycans. Nature. 2021;597:92-6. [CrossRef]
178. Hadi F, Kulaberoglu Y, Lazarus KA, Bach K, Ugur R, Beattie P, et al. Transformation of naked mole-rat cells. Nature. 2020;583:E1-7. [CrossRef]
179. Bunescu A, Abdelhamid Y, Gaunt MJ. Multicomponent alkene azidoarylation by anion-mediated dual catalysis. Nature. 2021;598:597-603. [CrossRef]
180. Baran MT, Miziak P, Bonio K. Characteristics of carotenoids and their use in the cosmetics industry. J Educ Health Sport. 2020;10:192-6. [CrossRef]
181. Mendes-Silva TCD, Andrade RFS, Ootani MA, Mendes PVD, SáRAQC, Silva MRF, et al. Biotechnological potential of carotenoids produced by extremophilic microorganisms and application prospects for the cosmetics industry. Adv Microbiol. 2020;10:397-410. [CrossRef]