
1. INTRODUCTION
Rice is one of the principal staple foods for more than 3.5 billion people in the world [1], and Asia contributes 90% of global rice production [2]. Rice is used as a major staple food by more than two-thirds of the Indian population [3] and also it plays a crucial role in people’s food and livelihood security. The total rice production in India during 2019–2020 is estimated at 116.5 metric tons (Mt), the second-highest in the world [4] and as per estimate approximately 150 Mt of rice will be needed in India by 2050 [5].
Kalanamak rice (Oryza sativa L.) is non-basmati aromatic rice, has high quality and intermediate grain size, and is largely recognized for its aroma and taste. It is indigenous, predominantly cultivated, and consumed in the north-east part of Uttar Pradesh [6]. The government of India has granted it geographical indication tag in 2012. The global value of Kalanamak rice is due to significantly higher antioxidant levels, Vitamin E, grain quality, and aroma [7,8]. According to Chaudhary et al. [6], farmers’ interest in the cultivation of Kalanamak rice has been lessened due to many reasons, namely, tall stature (150 cm), long-duration (145 days), and non-availability of quality seeds. Revitalization of interest for Kalanamak rice required focused breeding steps to improve its genotypes for long and slender grain type, semi-dwarf to dwarf plant, lodging resistance, and resistance to biotic and abiotic stresses, and short duration of maturity [9].
Gamma rays have been extensively used to develop 92% of rice mutant varieties [10]. According to the current database of FAO/IAEA-MVD [11], nearly 22 rice mutant varieties have been developed by physical mutagen (gamma-rays) in India. Recent studies have documented that exposure to gamma-irradiation can improve tolerance to abiotic stress conditions, such as salt and drought [12,13]. The exposure of biological systems to gamma rays can damage molecules, primarily through the ionization of protein and nucleic acid. It can also cause damage to the external medium through the production of reactive oxygen species (ROS) [14]. Accumulation of ROS in plant cells is prevented by the assistance of various ROS detoxifying enzymes and various non-enzymatic antioxidants [15,16]. The seeds of Kalanamak rice were treated with gamma rays at various intensities (100, 150, 200, 250, and 300) Gy. The purpose of this study was to induce genetic variation and to analyze the effects of gamma-irradiation on morpho-agronomic and yield attributes.
2. MATERIALS AND METHODS
2.1. Plant Material and Radiation Treatments
Rice (Oryza sativa L.) seeds were procured by Participatory Rural Development Foundation (PRDF), Gorakhpur, U.P., India. These were exposed to gamma rays at the National Botanical Research Institute (NBRI), Lucknow located at 26.8° N and 80.9° E. The treatment was given at the various intensities of gamma radiation 100, 150, 200, 250, and 300 Gy in 2019 using a Cobalt60 (60Co) source. Seeds were packed in butter paper covers and placed in a gamma chamber 60Co gamma source for a suitable time for an individual dose based on the half-life of the source.
2.2. Seedling Tray Trial
The seedling tray trail experiment was conducted in the Department of Botany, Deen Dayal Upadhyaya Gorakhpur University, Gorakhpur located at 26.7° N and 83.3° E; by following the method of Shimelis et al. [17] with a few modifications. Control and the irradiated gamma-ray seeds at different doses were planted at a depth of about 1 cm in the seedling tray under natural conditions using a fertile farm soil growth medium. In an entirely randomized design, two seeds were sowed per hole with three replications. For each gamma-ray treatment, 196 seeds were planted per replica. The seedlings were watered two times daily using a mist irrigation system.
2.3. Data Collection
Morpho-physiological characteristics were noted from germination to 14 days. After germination, the germination percentage was calculated as the ratio of germinated seeds to the total number of seeds sown on the 7th day after sowing. The seedling height was measured as the length from the plant’s base to the flag leaf’s tip. The Percentage of seedling survival and seedling height, taken after 14 days of seed germination, and the average seedling height were measured based on randomly selected 100 seedlings for each treatment.
2.4. Preparation of Enzyme Extracts
After 14 days of seedling, samples were collected and powdered in liquid nitrogen before homogenization. Enzyme-specific buffers were used for the extraction of homogenized samples in enzymatic analysis. Homogenate was centrifuged at 15,000 × g for 10 min at 4°C. Supernatant of each treatment was separately collected and used for different enzyme assays.
2.5. Enzyme Activities
Catalase (CAT) activity was assayed using the method of Ye et al. [18]. The superoxide dismutase (SOD) activity was assayed by adopting the method provided by Imanari [19]. The lipid peroxidation (LPO) products were estimated by the formation of thiobarbituric acid reactive substances (TBARS) and quantified in terms of malondialdehyde (MDA) as described by Haraguchi et al. [20]. The total soluble protein in the seedling extract was determined by Lowry’s method and crystalline bovine serum albumin was used as a standard [21].
2.6. Field Trial
The field trial experiment was conducted in the Educational and Research Botanical Garden (ERBG) of the Department of BotanyDeen Dayal Upadhyaya Gorakhpur University, Gorakhpur. A nursery of gamma-irradiated M1 seeds was implanted in the field with 300 control seeds. After 20 days, seedlings were subsequently transplanted in a randomized block design along with control. After maturation of the surviving seedlings, different agronomic characters, namely, plant height (cm), leaf length (cm), leaf width (cm), the total number of tillers per plant, panicle length (cm), the total number of grains per panicle, percent of filled grains per panicle, and number of primary and secondary branching per panicle were evaluated. During the growth period of the crop, all the suggested cultural practices such as irrigation, weeding, and plant protection methods were followed.
2.7. Statistical Analysis
All experimental data were expressed as the mean ± SD. Error bars were made for the variables. Kolmogorov–Smirnov test was applied to check for the normality of variables. P < 0.05 was considered to be statistically significant. To check for the difference between the control and 150 Gy, a two-sample independent t-test or its corresponding non-parametric test: the Mann–Whitney U test, was applied depending on the normality of the variables in both the groups. One-way analysis of variance (ANOVA) was also performed to check whether there is a significant difference in the means of percentage seed germination (7 DAP), percentage of seedling survival (14 DAP), and seedling height (14 DAP) across different groups. All the analyses were done using PRISM 5.01 and SPSS v20 software.
3. RESULTS AND DISCUSSION
3.1. Seedling Tray Trial
In the seedling tray trial experiment, a gradual reduction in the percentage of germination, percentage of seedling survival, and seedling height with an increased dose of gamma radiation was observed [Table 1]. The percent of seed germination is higher in control (88.43%), followed by 100 and 150 Gy-treated seeds. In control, the percentage of germination, seedling survival, and seedling height was (88.43 ± 1.56), (81.73 ± 6.77) and (12.70 ± 0.15), respectively. While, in 100 Gy-treated seeds, the percentage of seed germination, seedling survival, and seedling height was reported (70.40 ± 1.02), (63.24 ± 7.46) and (9.75 ± 1.28), respectively. A significant reduction was observed in the percentage of seed germination at the higher doses of gamma radiation, namely, 250 Gy (39.96 ± 2.41) and 300 Gy (38.26 ± 1.35). The higher level of DNA damage caused by gamma irradiation to the seed embryo points to a drop in the percentage of seedling survivability to (42.05 ± 5.10) at 250 Gy and (37.80 ± 1.30) at 300 Gy. The seedling height at 250 Gy and 300 Gy has also been severely affected. In the selected mid-range of 150 Gy and 200 Gy, the seed germination percentage is found to be (62.07 ± 0.78) and (58.67 ± 3.06), respectively.
Table 1: Average percentage of seed germination, seedling survival, and height of seedlings in control and gamma ray treated rice plant cultivar Kalanamak.
| Treatment | Percentage of seed germination (7 DAP) | Percentage of seedling survival (14 DAP) | Seedling height (cm) (14 DAP) |
|---|---|---|---|
| Mean±SD | Mean±SD | Mean±SD | |
| Control | 88.43±1.56 | 81.73±6.77 | 12.70±0.15 |
| 100 Gy | 70.40±1.02 | 63.24±7.46 | 9.75±1.28 |
| 150 Gy | 62.07±0.78 | 54.62±1.99 | 8.88±0.44 |
| 200 Gy | 58.67±3.06 | 43.06±3.93 | 8.86±0.01 |
| 250 Gy | 39.96±2.41 | 42.05±5.10 | 4.01±0.44 |
| 300 Gy | 38.26±1.35 | 37.80±1.30 | 3.86±0.13 |
| F-test value | 306.87* | 33.42* | 104.26* |
* P<0.05; DAP: Days after plantation; SD: Standard deviation
3.2. Enzyme Activities
The antioxidant enzymatic assays highlight how a high dose of gamma radiation can induce redox imbalance in seeds leading to an intracellular increase in ROS concentration and subsequent alterations in various oxidative stress markers and drastically affecting the DNA content of seed embryos [22]. Rice seeds were exposed to different doses, namely, 100, 150, 200, 250, and 300 Gy. Although, a dosage of 150 Gy represents very effective antioxidant enzymatic activity compared to control.
Antioxidant enzyme assays for CAT, SOD, and MDA are performed within control and numerous gamma-ray irradiated seedlings. After 14 days from germination, the enzyme assays were performed to determine the most favorable dose that maintains their antioxidant status, which may improve the yield and grain quality in the next generation. Three independent experiments were conducted for each antioxidant enzyme in each treatment group that contains 100 seeds.
Superoxide dismutase enzymatic activity in 150 Gy seedlings significantly increased (P < 0.05) after 14 days compared with the control [Figure 1a]. SOD is the decisive antioxidant biomarker that maintains cellular redox balance [23]. Antioxidant enzymatic activity of CAT shows a similar result as SOD (150 Gy treated seedlings) [Figure 1b]. Peroxidase is considered the key enzyme for the decomposition of H2O2, especially under CAT inactivation [24]. Decreased levels of CAT and increased peroxidase activity induced by oxidative stress reflected a general strategy required to confound the stress and rescue the cells against damage. Lipid peroxidation (LPO) is measured in the form of MDA because it is the end product of lipid peroxidation. Increased LPO damages membrane function by reducing membrane fluidity and altering the activity of membrane-bound enzymes and receptors [25]. At the higher dose of gamma radiation, that is, 250 and 300 Gy, LPO significantly (P < 0.05) increased in comparison to 150 Gy treated seedlings [Figure 1c]. Protein is also one of the best parameters to know the physiological activity of plants. The gamma radiation-induced loss of total protein was analyzed by determining the concentration of the total soluble proteins. At the higher dose of gamma radiation (250 and 300 Gy), the soluble protein content significantly (P < 0.05) decreased in comparison to the 150 Gy treatment [Figure 1d]. Hence, we provide experimental evidence for the beneficial effect of a 150 Gy dose of gamma radiation for further study. This dose of gamma radiation could be very effective for screening desired traits in the M2 generation.
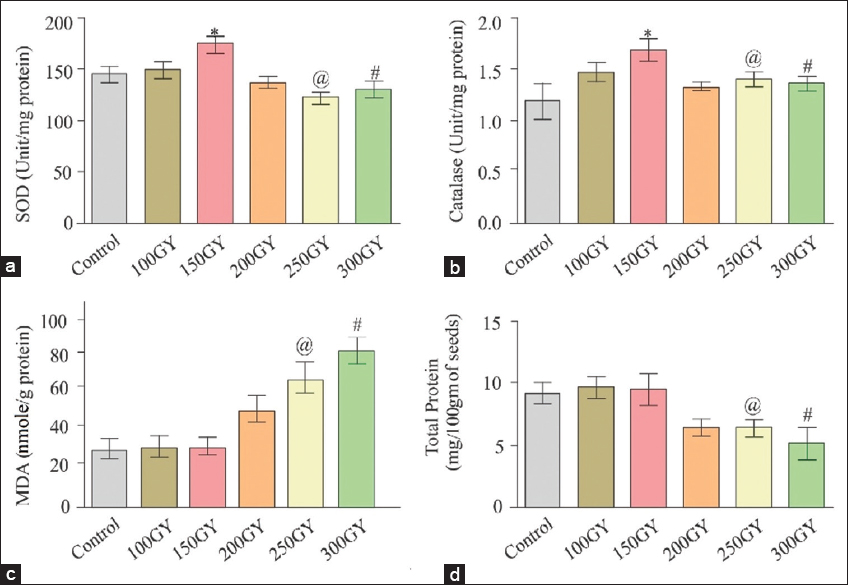 | Figure 1: Influence of gamma-ray irradiation on (a) superoxide dismutase (SOD); (b) Catalase; (c) malondialdehyde (MDA); and (d) total protein. (a) Represents the SOD enzymatic activity. Data are expressed as mean ± standard deviation (SD) of 3 independent experiments and each experimental group contains 100 seeds. *Denotes significantly increased (P < 0.05) in SOD activity when compared with @ and # while decreased significantly (P < 0.05) when compared with in 150 Gy treatment. (b) Represents the catalase (CAT) enzymatic activity. Data are expressed as mean ± SD of 3 independent experiments and each experimental group contains 100 seeds. *Denotes the activity of CAT enzyme which is significantly increased (P < 0.05) when compared with control. While @ and # significant (P < 0.05) decrease when compared with in 150 Gy treatment. (c) Represents lipid peroxidation is measured in the terms of MDA. Data are expressed as mean ± SD of 3 independent experiments and each experimental group contains 100 seeds. @ and # represent significant (P < 0.05) increase when compared with in 150 Gy treatment and control. (d) Represents total protein content. Data are expressed as mean ± SD of 3 independent experiments and each experimental group contains 100 seeds. @ and # represent significant (P < 0.05) decrease when compared with in 150 Gy treatment. [Click here to view] |
3.3. Evaluation of Morpho-agronomic Parameters
The agro-morphological characteristics are shown in [Table 2]; all ten parameters were compared between the control and 150 Gy gamma-irradiated plants. Before checking for the significant difference, each parameter was checked for normality individually in control and 150 Gy irradiated plants. For variables found to be normally distributed in both, an independent t-test was applied to check if there was any significant difference between the mean of control and plants treated at 150 Gy gamma radiation. However, variables were found to be non-normally distributed between control and 150 Gy irradiated plants, the corresponding non-parametric test – Mann–Whitney U test, was applied to check for a significant difference.
Table 2: Mean performance for different traits in gamma ray treated M1 generation rice cultivar Kalanamak.
| Trait | Control (mean±SD*) | 150 Gy (mean±SD*) | t/Z | P-value |
|---|---|---|---|---|
| Leaf width (cm) | 1.35±0.09 | 1.23±0.10 | −9.321a | <0.05 |
| Leaf length (cm) | 53.73±7.76 | 49.73±9.40 | 3.915b | <0.05 |
| Plant height (cm) | 182.77±9.45 | 152.28±17.78 | 19.384b | <0.05 |
| No. of tillers per plant | 6.51±0.75 | 9.07±3.57 | −6.391a | <0.05 |
| Panicle length (cm) | 27.60±1.34 | 26.32±2.01 | −5.852a | <0.05 |
| No. of grains per panicle | 210.03±24.87 | 234.38±42.17 | −6.271b | <0.05 |
| Percent of filled grains per panicle | 84.62±11.16 | 50.99±27.17 | −10.319a | <0.05 |
| Primary branching number per panicle | 14.58±1.09 | 15.13±1.17 | −3.844a | <0.05 |
| Secondary branching number per panicle | 33.62±6.73 | 37.92±7.30 | −4.715a | <0.05 |
| 1000 seed weight (g) | 14.92±0.56 | 11.82±4.64 | −2.114a | 0.035 |
Leaf width (cm) was found to be non-normally distributed by applying the Kolmogorov–Smirnov test for normality. The plants that received treatment of 150 Gy have significantly lower leaf width than the control. Leaf length (cm) and plant height (cm) were normally distributed. Hence, independent samples t-test was applied to check for the significant difference in the mean of 150 Gy irradiated plants and control.
The number of tillers per plant and panicle length(cm) was found to be irregularly distributed; hence, the Mann–Whitney U test was applied to check for significant differences. Both showed significant P-values at a 5% significance level, indicating that the median between the 150 Gy irradiated plants and control is not the same. There was a significant increase in the number of tillers per plant in 150 Gy irradiated plants compared to the control. In comparison, panicle length (cm) reduced significantly for the plants of 150 Gy. An independent t-test was applied to check if there is a significant difference in the average number of grains per panicle. It was found that the average number of grains per panicle was significantly higher for the plants that received 150 Gy (234.38) as compared to the control (210.03). The percentage of filled grains per panicle, number of primary and secondary branching number per panicle, and 1000 seed weight (gm) were all non-normally distributed; hence, Mann–Whitney U test was applied to check for any significant difference between the control and 150 Gy irradiated plants. The null hypothesis of no difference was rejected for all four indicators. A substantial reduction was observed in the percentage of filled grains per panicle for the plants that received 150 Gy compared to the control. Primary and secondary branching numbers per panicle significantly increased for the 150 Gy irradiated plants. A significant reduction in the 1000 seed weight (g) for the 150 Gy irradiated plants was (11.82) as compared to the control (14.92).
3.4. Mutants Showing Alterations
On screening M1 plants of 150 Gy, variations in phenotypic characters such as plant height, tiller count per plant, unproductive tillers, panicle length, and numbers of primary and secondary branches in a panicle are shown in [Table 3]. The panicle length of control plants was in the range of (23.5-30.5cm). In the 150 Gy mutant plants, the primary branching number per panicle (PBNPP) in 15 plants was observed to be more than (16). Although only in two mutant plants, the secondary branching number per panicle (SBNPP) was more than (54). The PBNPP and SBNPP of the control were in the mean range of (11.6–16.6) and (16–54.3), respectively.
Table 3: FOM (%) observed for different traits in 150 Gy M1 generation.
| Trait | Range | No. of mutant plants | FOM (%) |
|---|---|---|---|
| Plant height | Tall (151–203 cm)* | ? | ? |
| Semi-dwarf (120–151 cm) | 71 | 35.5 | |
| Dwarf (<120 cm) | 10 | 5 | |
| Tiller count/plant | 5–11* | ? | ? |
| >11 | 46 | 23 | |
| Panicle length | 23.5–30.5 cm* | ? | ? |
| >30.5 cm | 2 | 1 | |
| Primary branching number per panicle | 11.6–16.6* | ? | ? |
| >16.0 | 15 | 7.5 | |
| Secondary branching number per panicle | 16–54.3* | ? | ? |
| >54.3 | 2 | 1 |
FOM (%): Frequency of mutation (%);
* Control
4. CONCLUSION
The experimental evidence from the present study reflects the positive effects of gamma radiation on the agronomic traits of Kalanamak rice. Several variants have shown high yield and less plant height than the control. The retrieved germplasm could be utilized to make improved lines. Selection of abiotic stress resistance genotypes can also be achieved in the future as the effective dose of gamma radiation did not negatively affect the antioxidant potential. This study will be helpful for researchers working on genetic improvement aspects of the same genotype or other related cultivars.
5. ACKNOWLEDGMENT
We are grateful to Dr. R.C. Chaudhary, Chairman PRDF, Gorakhpur, U.P., India, for providing us with the germplasm of the cultivar Kalanamak of rice (Oryza sativa L.), and to Dr. Hemant Kumar Yadav, Principal Scientist, National Botanical Research Institute (NBRI), Lucknow for his support in the treatment of seeds with the gamma radiation. We express our thanks to the Director, Botanical Survey of India, Kolkata for his valuable support. I am also very grateful to Mr. Raushan Kumar, research scholar, Department of Biochemistry, University of Allahabad, Allahabad, U.P., for his support in enzymatic assays and Mr. Vishnu Kumar, research scholar, Department of Botany, DDU Gorakhpur University, U.P., for his valuable suggestion in the preparation of manuscript.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors requirements/guidelines.
7. FUNDING
There is no financial aid for conducting this research.
8. CONFLICTS OF INTREST
The authors of this manuscript have no conflicts of interest.
9. ETHICAL APPROVALS
This research did not involve any experiments on animal
10. DATA AVAILABILITY
All experimental data are available within this manuscript.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Chen T, Shabala S, Niu Y, Chen ZH, Shabala L, Meinke H, et al. Molecular mechanisms of salinity tolerance in rice. Crop J 2021;9:506-20. [CrossRef]
2. Ngalimat MS, Hata EM, Zulperi D, Ismail SI, Ismail MR, Zainudin NA, et al. Plant growth-promoting bacteria as an emerging tool to manage bacterial rice pathogens. Microorganisms 2021;9:682. [CrossRef]
3. Jambhulkar NN, Panigrahi US, Bisen J, Mondal B, Mishra SK, Kumar GA. Growth rate and instability analysis of rice area, production and yield in Punjab. Pharm Innov J 2021;10:352-5.
4. Kaur R, Kaur S, Deol JS, Sharma R, Kaur T, Brar AS, et al. Soil properties and weed dynamics in wheat as affected by rice residue management in the rice wheat cropping system in South Asia:A review. Plants 2021;10:953. [CrossRef]
5. Pathak H, Nayak AK, Jena M, Singh ON, Samal P, Sharma SG. Rice research for enhancing productivity, profitability and climate resilience. In:Pathak H, Nayak AK, Jena M, Singh ON, Samal P, Sharma SG, editors. Rice research for enhancing productivity, profitability and climate resilience. Cuttack, Odisha, India:ICAR-National Rice Research Institute;2018. 1-17.
6. Chaudhary RC, Mishra SB, Yadav SK, Ali J. Extinction to distinction:Current status of Kalanamak, the heritage rice of eastern Uttar Pradesh and its likely role in farmers'prosperity. Gene 2012;16:16.
7. Rajendran V, Sivakumar HP, Marichamy I, Sundararajan S, Ramalingam S. Phytonutrients analysis in ten popular traditional Indian rice landraces (Oryza sativa L.). J Food Meas Charact 2018;12:2598-606. [CrossRef]
8. Yadav SK, Chaudhary RC, Kumar S, Mishra SB. Breakthrough in improving farmers'income sustainably by producing Kalanamak rice. J Agri Search 2019;6:1-5. [CrossRef]
9. Kumar S, Deo I. Studies on genetic variability, heritability and genetic advance in advance lines of Kalanamak aromatic rice (Oryza sativa L.). Eco Env Cons 2016;22:891-5.
10. Food and Agriculture Organization of the United Nations/International Atomic Energy Agency-Mutant variety database [FAO/IAEA-MVD]. Available from:https://www.mvd.iaea.org/#!Search [Last accessed on 2019 Mar 30].
11. Food and Agriculture Organization of the United Nations/International Atomic Energy Agency mutant variety database [FAO/IAEA-MVD]. Available from https://www.mvd.iaea.org/#!Search [Last accessed on 2021 Mar 09].
12. Viana VE, Pegoraro C, Busanello C, de Oliveira A. Mutagenesis in rice:The basis for breeding a new super plant. Front Plant Sci 2019;10:1326. [CrossRef]
13. Oladosu Y, Rafii MY, Abdullah N, Hussin G, Ramli A, Rahim HA, et al. Principle and application of plant mutagenesis in crop improvement:A review. Biotechnol Biotechnol Equip 2016;30:1-16. [CrossRef]
14. Esnault MA, Legue F, Chenal C. Ionizing radiation:Advances in plant response. Environ Exp Bot 2010;68:231-7. [CrossRef]
15. Macovei A, Garg B, Raikwar S, Balestrazzi A, Carbonera D, Buttafava A, et al. Synergistic exposure of rice seeds to different doses of-ray and salinity stress resulted in increased antioxidant enzyme activities and gene-specific modulation of TC-NER pathway. BioMed Res Int 2014;2014:676934. [CrossRef]
16. Silva AS, Danielowski R, Braga EJ, Deuner S, Junior AM, Peters JA. Development of rice seedlings grown from pre-hydrated seeds and irradiated with gamma rays. Ciênc Agrotec 2011;35:1093-100. [CrossRef]
17. Shimelis HA, Olaolorun BM, Mathew I, Laing MD. Optimising the dosage of ethyl methanesulphonate mutagenesis in selected wheat genotypes. S Afr J Plant Soil 2019;36:357-66. [CrossRef]
18. Ye N, Li H, Zhu G, Liu Y, Liu R, Xu W, et al. Copper suppresses abscisic acid catabolism and catalase activity, and inhibits seed germination of rice. Plant Cell Physiol 2014;55:2008-16. [CrossRef]
19. Imanari T. Improved assay method for superoxide dismutase. Igakuno Ayumi 1977;101:496-7.
20. Haraguchi H, Saito T, Okamura N, Yagi A. Inhibition of lipid peroxidation and superoxide generation by diterpenoids from (Rosmarinus officinalis L.). Planta Med 1995;61:333-6. [CrossRef]
21. Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem 1951;193:265-75. [CrossRef]
22. Kurek K, Plitta-Michalak B, Ratajczak E. Reactive oxygen species as potential drivers of the seed aging process. Plants 2019;8:174. [CrossRef]
23. Kurutas EB. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress:Current state. Nutr J 2015;15:1-22. [CrossRef]
24. ?uba V, PavelkováT, Mú?ka V. Effect of dose and dose rate of gamma radiation on catalytic activity of catalase. J Radioanal Nucl Chem 2010;286:619-24. [CrossRef]
25. Taso OV, Philippou A, Moustogiannis A, Zevolis E, Koutsilieris M. Lipid peroxidation products and their role in neurodegenerative diseases. Ann Res Hosp 2019;3:1-10. [CrossRef]