
1. INTRODUCTION
Currently, biofertilizers and biostimulants are applied in diverse agricultural crops to reduce the problems associated with excessive application of mineral fertilizers, such as soil fertility degradation and environmental pollution. Biostimulants have been shown to improve soil structure, water retention, and microbial activity. In plants, they stimulate growth, enhance seed germination, and elevate metabolic processes. Furthermore, biostimulants contribute to improved photosynthesis, nutrient assimilation, and resilience to abiotic stress, leading to higher yields, better crop quality, and enhanced overall plant functionality. Consequently, biostimulants have become a valuable tool in optimizing agricultural productivity [1-3].
The application of microalgae as biofertilizers and biostimulants has emerged as a natural, environmentally sustainable, biodegradable, and cost-effective strategy, with no reported risks to human or animal health [4]. Microalgal species, particularly those from the genus Nostoc, have been employed by their capacity of fix atmospheric nitrogen and enhance phosphorus bioavailability through solubilization and mobilization, resulting major yields in several crops [5]. Similarly, Spirulina was tested as a biofertilizer for maize, utilizing its nutrient-dense biomass, which is rich in bioavailable micronutrients, to enhance crop productivity [6].
The biostimulant properties of Spirulina (Arthrospira maxima) are attributed to carbohydrates and other compounds [3]. Hydrolyzed Spirulina has also been utilized as a biostimulant in lettuce (Lactuca sativa), where its hydrolyzed proteins release polyamines and amino acids that act as growth promoters [7,8]. These hydrolyzed proteins are essential in activating biochemistry processes to promote plant and root growth, enhance yield, influence soil microbiota composition, optimize water and nutrient uptake, and alleviate the effects of abiotic stress [9].
Hydrolyzed proteins of both animal and plant origins, including those derived from microalgae, increased carbon and nitrogen metabolism by enhancing nitrogen uptake [10]. However, the Spirulina studies on maize, in terms of enhancing bioactive compounds with health benefits, remain largely unexplored. Existing studies on Spirulina in maize have primarily focused on agronomic parameters. The present investigation is the first to evaluate the impact of Spirulina as biostimulants on the bioactive compounds in pigmented maize varieties rich in anthocyanins.
Corn is an ancient culture originating from Mexico and has been a staple food since pre-Hispanic times [11]. It is the most widely cultivated crop worldwide, appreciated for its high yield and versatility. The origin, domestication, and diversity of this crop is Mexico, with a harvested area of 1,021,501 ha, yield of 6.818 t/ha, and harvests of 6,964,201 t [12]. Among the 220 recognized maize landraces, 59 are native to Mexico, including more than 45 pigmented varieties [13].
The pigmentation in maize is predominantly attributed to anthocyanins, primarily cyanidin, the most abundant type, along with peonidins and pelargonidins, depending on the specific variety [14-18]. Anthocyanins, a subclass of flavonoids, are known for their significant health benefits, primarily due to their potent free-radical-scavenging properties and their ability to activate key enzymes involved in cellular oxidative-stress management [19,20].
Anthocyanins have attracted considerable attention by sensory attributes, in addition to an extensive antioxidant activity. They have been shown to inhibit the growth of Helicobacter pylori, a bacterium linked to duodenal ulcers and gastric cancer, and to prevent conditions such as diabetic retinopathy, glaucoma, retinitis pigmentosa, and cataracts. Moreover, anthocyanins enhance neuro-cognitive functions and are DNA protector [21-23]. They also prevent cardiovascular diseases, arthritis, and cancer and are hypoglycemics [24-26].
Given these health benefits, investigating the bioactive compounds in pigmented maize with antioxidant activity and preventive properties against degenerative diseases holds significant relevance. The purpose of this research was to evaluate a biostimulant based on hydrolyzed Spirulina on levels of polyphenols, flavonoids, anthocyanins, and antioxidant activity of three corn varieties.
2. MATERIALS AND METHODS
2.1. Plant Material and Treatments
Maize seeds of black, purple, and red varieties from the western maize landrace from the Valles region, Jalisco, were sown in the experimental field of the University Center of Biological and Agricultural Sciences UDG. Sowing was conducted on June 14, 2022, using a completely randomized block design with four plots of 20 m2 (4 m long × 5 m wide), each containing six furrows measuring 4 m in length and 0.8 m in width and separated by 1 m of distance each experimental units and replicates. One plot was designated for controls, comprising black control (BC), red control, and purple control maize without Spirulina application (two per furrow). The remaining three plots were sown with each color variety individually-black treatment, purple treatment, and red treatment.
A solution of 5 mL/L of commercial hydrolyzed Spirulina (Ficocyan®) diluted in water was applied to seedlings at emergence (June 23, 2022) and subsequently every month until fruiting (October 21, 2022) for a total of four applications. This dosage and frequency of application have been tested to increase yield up to 12% in unpigmented corn in previous studies. Physicochemical analyses were performed on the soil used in the experiment [Table 1].
Table 1: Physicochemical properties of the soil used for planting the experimental maize varieties.
| Determination | Results | |
|---|---|---|
| pH | 6.5 | |
| Electrical conductivity (ds/m) | 0.31 | |
| Organic matter (%) | 3.01 | |
| Texture | Sandy loam | |
| Sand (%) | 44.92 | |
| Silt (%) | 36.42 | |
| Clay (%) | 15.44 | |
| Macroelements | ppm | |
| Nitrogen (NO3) | 11 | |
| Phosphorus | 18 | |
| Potassium (acetate) | 473 | |
| Calcium (acetate) | 2,073 | |
| Magnesium (acetate) | 321 | |
| Microelements | ppm | |
| Copper | 1.1 | |
| Manganese | 4.5 | |
| Zinc | 1.3 | |
| Iron | 6.7 | |
| Cations | meq/100g | Percentage |
| Sodium | 0.53 | 3.6 |
| Potassium | 1.21 | 8.2 |
| Calcium | 10.37 | 70.2 |
| Magnesium | 2.64 | 17.9 |
2.2. Sample Preparation
Grains from each treatment were harvested separately, dehydrated inside a stove to 45°C, then ground with a Krups mill (Model GX410011V) fitted with a No. 60 sieve.
2.3. Extract Preparation
The extracts for analysis were made by homogenizing 0.5 g of flour from each treatment and control with 10 mL of a mixture of 80% methyl alcohol and 20% water, then subjected to sonication for 20 min using a Branson 3510 sonicator (power: 100 W, 42 kHz), followed by filtration and concentration with an R-215 rotary evaporator (Büchi).
2.4. Polyphenols Analysis
From each triplicate, 1 mL of extract was combined with 9 mL of water and 1 mL of Folin-Ciocalteu 2 N. Five min past, 10 mL of a 7% Na2CO3 solution was added, and the mixture was diluted to 25 mL with water. Samples remained 90 min before being read at 750 nm in a JENWAY 6320D spectrophotometer. Phenol content was determined using gallic acid at different concentrations (mg GAE)/g of dry weight [27].
2.5. Flavonoids Analysis
One mL from each extract by triplicate was taken to blend in 4 mL of high-performance liquid chromatography (HPLC)-grade water and 0.3 mL of 5% NaNO2 solution. Passing 5 min, 0.3 mL of 10% AlCl3 solution was added. Then to 6 min, 2 mL of 1 M NaOH solution was added and diluted to 10 mL with water. Samples were mixed in vortex, and it was read at 510 nm in a JENWAY 6320D spectrophotometer. Flavonoids were determined with (+)-catechin to different concentrations (mg EC)/g [27].
2.6. Anthocyanin Quantification
For this analysis, aliquots of 200 μL were taken by triplicate and added to 1800 μL of sodium acetate buffer (pH 4.5), while another 200 μL was combined with 1800 μL of potassium chloride buffer (pH 1.0). The mixtures were thoroughly vortexed and read at 510 nm and 700 nm in a UV-VIS JENWAY 6320D spectrophotometer [28]. Final absorbance was estimated using the formula:
A = (A510–A700) at pH 1.0–(A510–A700) at pH 4.5.
The content was expressed as mg C3G/g and calculated with the formula:
TAC = A × MW × DF × 1000/ε × 1
TAC = Total anthocyanins content
A = Final absorbance
MW = Molecular weight of cyanidin-3-glucoside (449.2 g/moL)
DF = Dilution factor utilized
ε = Molar absorptivity (26,900 L/cm/mol)
2.7. Determination of Antioxidant Capacity
Two techniques to evaluate the antioxidant capacity of samples were used, with slight modifications. Phenolic extracts (0.2 mL) were mixed with 3.3 mL of DPPH (60 μM) solution in 96-well microplates. After 30 min, the absorbance was read at 510 nm in a Mca. Instruments advanced microwave radiometer (AMR-100) microplate reader [29].
The second technique employed was the ABTS assay. A 7.4 mM ABTS solution was prepared and mixed in 1 mL of 2.6 mM potassium persulfate (K2S2O8) solution. Then, at 12 h to allow for radical formation, and subsequently tuned to absorbance to 0.7 at 734 nm. From this mixture, 3.3 mL was combined with 0.2 mL of the extract in 96-well microplates. Absorbance was read at 734 nm using a Mca. Instruments AMR-100 microplate reader after incubation in the dark for 2 h. Results from both techniques were expressed as millimoles of Trolox equivalents/g [29].
2.8. Statistical Analysis
The results were processed with an analysis of variance and Tukey’s test for mean comparisons. Statistical analyses were performed using Stat Graphics 19 software.
3. RESULTS
3.1. Polyphenols
Polyphenol revealed a rise in red maize variety with Spirulina, of 84.2 mg GAE/g in comparison to 75 mg GAE/g in untreated sample. In the black variety, the treated sample exhibited a slightly higher polyphenol content (83.4 mg) in comparison to the control (82.8), without difference. Conversely, the purple variety showed a negative effect, as untreated seeds contained significantly higher polyphenol levels (85) in comparison to treated seeds (76.6) [Table 2].
Table 2: Polyphenol and flavonoid content in black, purple, and red maize from the western maize landrace of the Valles region, Jalisco, under the effect of hydrolyzed Spirulina.
| Polyphenols (mg GAE/g sample) | Flavonoids (mg CE/g sample) | |||
|---|---|---|---|---|
| Sample | Average±SD | VC (%) | Average±SD | VC (%) |
| BC | 82.8±3.15a | 3.81 | 94.6±5.2bc | 5.5 |
| RC | 75±3.52b | 4.7 | 91.6±11.0bc | 12.0 |
| PC | 85±2.1a | 2.46 | 81.0±11.0c | 13.5 |
| BT | 83.4±2.8a | 3.4 | 106.4±16.8bc | 15.8 |
| RT | 84.2±1.47a | 1.75 | 253.5±42.62a | 16.8 |
| PT | 76.6±4.13b | 5.38 | 136.2±47.8b | 35.1 |
BC: Black control, RC: Red control, PC: Purple control, BT: Black treatment, RT: Red treatment, PT: Purple treatment.
Means with the same letter indicate no significant difference (P>0.05).
3.2. Flavonoids
Regarding flavonoid content, an increase was observed in the red maize variety treated with Spirulina, with 253.5 mg EC/g compared to 91.6 mg EC/g in the untreated sample. Similarly, the treated purple variety exhibited higher flavonoid content (136.2 mg EC/g) compared to the untreated sample (81.0 mg EC/g). In the black maize variety, the treated sample showed a slightly higher flavonoid content (106.4 mg EC/g) compared to the control (94.6 mg EC/g) without difference [Table 2].
3.3. Anthocyanins
The Spirulina treatment significantly increased anthocyanin content in the black maize variety, with treated samples exhibiting 67.9 mg C3GE/g compared with 23.6 mg C3GE/g in the BC; no differences were observed with other treatments. In the purple maize variety, the treated samples exhibited slightly higher anthocyanin content (35.0 mg C3GE/g) in comparison to the control (30.2 mg C3GE/g). In red maize variety, comparable levels of total anthocyanins were observed between the treated and control samples [Figure 1].
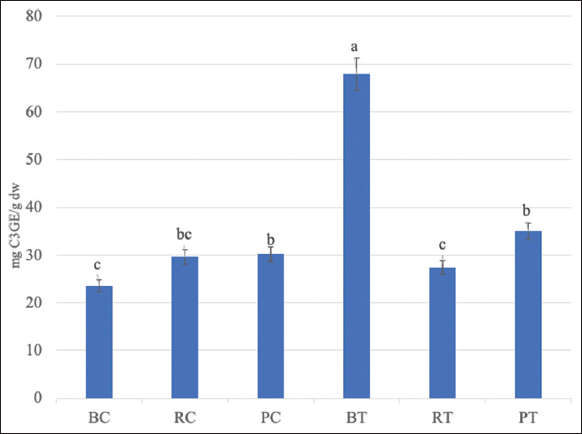 | Figure 1: Anthocyanin content (mg C3GE/g dry weight) in pigmented maize from the western maize landrace of Jalisco under the effect of hydrolyzed Spirulina. BC: Black control, RC: Red control, PC: Purple control, BT: Black treatment, RT: Red treatment, PT: Purple treatment. Means with the same letter indicate no significant difference (P > 0.05). [Click here to view] |
3.4. Antioxidant Activity
The the use of DPPH assay showed no effect of Spirulina treatment. In black maize variety, the control sample exhibited a significantly higher antioxidant activity (1690.7 μmoL TE/g) in comparison to the treated sample (1438.1 μmoL TE/g). Similarly, in red variety, the control disclosed slightly major antioxidant activity in comparison to the treated sample (1490.0 vs. 1404.4 μmoL TE/g sample), without difference. In the purple variety, the treated sample exhibited higher antioxidant activity (1593.1 μmoL TE/g) in comparison to the control (1374.3 μmoL TE/g sample) without difference statistically (P > 0.05).
In contrast, ABTS assay revealed differences in red and purple maize varieties treated with Spirulina. The treated red variety exhibited a major antioxidant activity of 1210.1 μmoL TE/g in comparison to 861.8 μmoL TE/g in control, while the treated purple variety showed effect significant of 1127.0 μmoL TE/g compared to 742.7 μmoL TE/g in control. However, BC sample demonstrated major antioxidant activity than the treated black sample, of 1139.7 μmoL TE/g and 814.2 μmoL TE/g sample, respectively [Table 3]. In table 4 it can be seen normality (Shapiro-Wilk) and homogeneity of variance (Levene’s test) by DPPH and ABTS assays
Table 3: Antioxidant activity measured by DPPH and ABTS assays (μmol TE/g sample) in pigmented maize from the Valles region, Jalisco, under the effect of hydrolyzed Spirulina.
| Sample | Average±SD | VC (%) | Average±SD | VC (%) |
|---|---|---|---|---|
| DPPH (μmol TE/g sample) | ABTS (μmol TE/g sample) | |||
| BC | 1690.7±43.8a | 2.6 | 1139.7±26ab | 2.3 |
| RC | 1490±33.4c | 2.2 | 861.7±28c | 3.2 |
| PC | 1374.32±15.3d | 1.1 | 742.7±31.1d | 4.2 |
| BT | 1438.1±26.8b | 1.8 | 814.2±37.2c | 4.6 |
| RT | 1404.4±36.1cd | 2.6 | 1210.1±3.2a | 0.3 |
| PT | 1593.1±33.3cd | 2.1 | 1127.0±18.8b | 1.7 |
BC: Black control, RC: Red control, PC: Purple control, BT: Black treatment, RT: Red treatment, PT: Purple treatment.
Means with the same letter indicate no significant difference (P>0.05).
Table 4: Normality (Shapiro-Wilk) and homogeneity of variance (Levene’s test) by DPPH and ABTS assays.
| Tests | ||
|---|---|---|
| DPPH | ||
| Method | Test statistics | P-value |
| Multiple comparisons | - | 0.326 |
| Levene | 1.00 | 0.447 |
 | ||
| ABTS | ||
| Method | Test statistics | P-value |
| Multiple comparisons | - | 0.543 |
| Levene | 0.41 | 0.838 |
 | ||
4. DISCUSSION
Polyphenol content in pigmented maize has been widely studied. Previous research [14,29,30] reported lower polyphenol values for purple maize varieties compared to the purple variety analyzed in this study (85 mg GAE/g). Similarly, Francavilla and Joye [31] documented lower polyphenol levels for black maize varieties (45.7–54.4 mg GAE/g) compared to our black variety (82.8 mg GAE/g), as well as lower values for purple (7–34) and red varieties (61.15). Red varieties of the Mixtec race, as reported by Herrera-Sotero et al. [26], also exhibited lower polyphenol levels (32.7–37.3 mg GAE/g). However, the polyphenol content observed in the varieties analyzed in this study (75–85 mg GAE/g) was lower compared to other pigmented maize varieties, such as those reported with 340.0 mg GAE/g and Zea mays var. Ceratina, which exhibited 183.43 mg GAE/g [32,33].
Anthocyanin content varies depending on maize variety and pigmentation, as highlighted by Francavilla and Joye [31]. A study on 15 maize genotypes from northeastern Mexico reported anthocyanin levels ranging from 1.38 mg C3G/g in white maize to 85.9 mg C3G/g in purple maize [30,32]. The untreated varieties analyzed in this study fall within this range, with anthocyanin content values of 23.6–30.2 mg C3G/g.
Secondary metabolites are influenced by interactions of edaphoclimatic factors. Variations in latitude, altitude, and season alter the amount of solar radiation received, which affects anthocyanin content. This is attributed to the effect of sunlight on the phenylpropanoid biosynthetic pathways and the accumulation of anthocyanin [34]. Similarly, soil conditions, low pH, and elevated electrical conductivity, combined with elevated levels of organic matter, especially nitrogen, have been shown to stimulate the synthesis of total phenols and anthocyanins in pigmented Mexican landrace accessions [35].
This is partially attributed to the carbon-to-nitrogen (C:N) balance, which may facilitate carbon allocation to the synthesis of phenolic compounds. However, the underlying mechanisms remain poorly understood in maize landraces [34]. Furthermore, agricultural management practices significantly influence maize production parameters. Unfortunately, limited studies have investigated the impact of sowing date and phosphorus-potassium-nitrogen (P-K-N) fertilization on pigmented maize landraces [34].
Our landraces were cultivated under consistent edaphoclimatic conditions, including summer climate and soil with an optimal pH and nitrogen, phosphorus, and potassium levels close to the recommended standards. Therefore, the results obtained could be attributed to the effect of hydrolyzed Spirulina, consistent with findings reported for other biostimulants derived from microalgal extracts [36].
The untreated maize varieties exhibited major anthocyanin content in comparison to pigmented maize studied by several authors [14,17,26,32]. Previous studies reported total anthocyanin values fluctuated from 0.01 to 7.54 mg ECG/g and 0.99–3.79 mg C3G/g in blue maize, 0.265–1.439 mg C3G/g in purple maize, 0.76–1.20 mg C3G/g in black maize, and 0.025–6.96 mg C3G/g in red maize. Similarly, other authors [15,37,38] stated anthocyanin values of 16.0, 3.2, and 5.0, respectively, in purple maize values lower than those observed in the untreated maize varieties analyzed in this study.
It has also been reported that for the purple maize variety, C3G is the major anthocyanin [37,38], who reported concentrations of 1.0–2.9 mg/g. However, anthocyanin of our maize variety was lower than reported for purple maize from China (55.8 mg C3G/100 g) [39]. In contrast, Peruvian black maize exhibited a higher anthocyanin content (48.50 mg C3G/g) [40] compared to the untreated black maize variety in this study. Notably, this value was surpassed by the Spirulina -treated black maize variety analyzed here, which exhibited an anthocyanin content of 67.9 mg C3G/g.
The combination of anthocyanins, such as cyanidin and delphinidin, and their derivatives has been reported to induce black pigmentation in plants [41]. Therefore, black maize varieties may contain higher levels of these anthocyanins, potentially stimulated by Spirulina hydrolysate. This hypothesis could be confirmed through HPLC of individual anthocyanins. The observed increase in these specific anthocyanins may result from interaction between genotypic traits and the abiotic factors discussed previously.
Reports on antioxidant activity in pigmented maize varieties are limited. Francavilla and Joye [31] reported DPPH values of 1400, 2200, and 2700 μmoL TE/g samples in purple, black, and red maize varieties, respectively, while Rodriguez-Salinas et al. [29] found a value of 1875.70 μmoL TE/g in purple maize. These values are higher than those observed in both treated and untreated maize varieties analyzed in this study for the same colors. However, the DPPH values in our red maize samples exceed those reported by Rodriguez-Salinas et al. (1127.70–1875.70 μmoL TE/g) and Herrera-Sotero et al. (10.5–12.7 μmoL TE/g) [26,29] for red maize.
Similarly, Harakotr et al. [30] reported ABTS values of 94.6 μmoL TE/g in Z. mays var. cera, which are significantly lower than those observed in this study (742.7–1210.1 μmoL TE/g). In contrast, Rodriguez-Salinas et al. [29] reported higher ABTS values, ranging from 2826.90 to 4263.90 μmoL TE/g in purple maize varieties.
Different techniques have been reported to evaluate the antioxidant capacity, which is determined by diverse factors. DPPH evaluate antioxidants that directly react with DPPH radicals, by finding a proton-donor substrate like an antioxidant, the radical is caught, and the absorbance decreases. DPPH is the principal radical used to measure antioxidant capacity in phenolic compounds [42].
ABTS methods are used by flavonoids and phenolic acids and are more specific than DPPH radicals. Scavenging takes place by electron donation. ABTS is oxidized to its radical cation, and ABTS+, is intensely colored [43].
However, few studies on biostimulants have focused on enhancing active compounds in maize. Most research has examined the use of biostimulants derived from macroalgae extracts to improve yield parameters [4].
Hydrolyzed Spirulina releases polyamines such as spermidine, putrescine, and spermine, which are hormone-like natural compounds capable of controlling biochemical and physiological functions in plants. These effects are likely mediated by the influence on gene and enzyme interactions, leading to improvements in crop quality and chemical composition, including polyphenol content [10,44].
This study focused on the stimulating effect of Spirulina as a promoter of bioactive compound production, with results showing considerable variability depending on the maize variety. A increase in polyphenol was shown in red maize variety in comparison with untreated sample, while no significant differences were detected in the black and purple varieties. Further investigation into the individual phenolic composition of these varieties is necessary to better understand these differences.
Anthocyanins are synthesized in plants through the phenylpropanoid pathway, with phenylalanine serving as the precursor. Although this biosynthetic mechanism is well established, few studies have examined how biostimulants influence this pathway in plants. Therefore, it is important to investigate the biochemical and molecular processes involved in their mechanism of action [45]. Only a few genes encoding anthocyanin biosynthetic enzymes have been cloned, primarily from maize, snapdragon, and petunia. The first flavonoid biosynthetic gene to be isolated was chalcone synthase, identified in parsley [46].
It is probable that the Spirulina hydrolysate stimulated the production of other non-anthocyanin phenolic compounds or carotenoids in the red maize variety, which may contribute to its coloration and are present in lower amounts in the black and purple maize varieties. Therefore, it is essential to conduct an analysis of individual phenolic compounds, as purple maize kernels are generally richer in anthocyanins compared to red ones [47].
Similar findings have been reported with biostimulants from seaweed extracts and hydrolyzed bean protein in sage (Salvia officinalis L.), which increased the yield of essential oils. However, other compounds, including total phenols, rosmarinic acid, and antioxidant capacity, were lower in treated plants compared to untreated ones [48].
The use of commercial biostimulants, such as AgroGain from red alga Kappaphycus alvarezii, has been reported to enhance growth parameters in maize plants. These biostimulants promote chlorophylls, carotenoids, sugars, amino acids, phenols (47% increase), and flavonoids (51% increase) by improving microbial counts in soil. In addition, they enrich metabolic pathways implicated in the assimilation of nitrogen, sulfur, and phosphorus [49]. These results are comparable to those observed in the red and purple maize varieties analyzed in this study.
Regarding anthocyanins, limited research has investigated the effect of biostimulants on maize. One notable study reported that using Ascophyllum nodosum algae extract enhanced maize plant growth and induced the accumulation of metabolites, including anthocyanins, by modulating genes [50].
Similarly, in Phaseolus vulgaris var. Toska, the application of Ecklonia maxima seaweed extracts increased phenol and anthocyanin levels. However, antioxidant activity by ABTS assay was lower compared to untreated samples [51]. This effect is attributed to the composition of E. maxima extract, which contains phytohormones, spermine and putrescine, brassinosteroids, phlorotannins, and abscisic acid, all of which contribute to the production of secondary metabolites [52].
5. CONCLUSION
Our results demonstrated a positive effect of hydrolyzed Spirulina on bioactive compounds in the evaluated maize landraces. Specifically, the red variety exhibited an increase in polyphenol and flavonoid content, likely due to the stimulation of other phenolic compounds by the Spirulina hydrolysate. The purple variety showed a significant increase in flavonoid content, while the black variety experienced the greatest enhancement in anthocyanin content following Spirulina treatment. Antioxidant capacity, by the DPPH assay, showed no significant differences in the treatments. In contrast, the ABTS assay revealed positive effects in the purple and red varieties. These findings underscore the potential of natural biostimulants like Spirulina to promote bioactive compounds in traditional Mexican foods, such as pigmented maize, in an environmentally sustainable manner. Notably, this is the first study to report such effects. Furthermore, these findings may contribute to the production of compounds with preventive properties against degenerative diseases, such as anthocyanins.
6. ACKNOWLEDGMENTS
The authors are grateful to Microalgas Oleas de México for providing the hydrolyzed spirulina-based biostimulant.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to the conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be authors as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Brown P, Saa S. Biostimulants in agriculture. Front Plant Sci. 2015;6:671. [CrossRef]
2. Meddich A. Biostimulants for resilient agriculture-improving plant tolerance to abiotic stress:A concise review. Gesunde Pflanzen. 2023;75:709-27. [CrossRef]
3. Vanajothi R, Bhavaniramya S, Vishnupriya S. Exploring the biostimulants in plant science. In:Harikesh BS, Anukool V, editors. New and Future Developments in Microbial. Biotechnology and Bioengineering. Amsterdam:Elsevier Inc.;2022. 1-25. [CrossRef]
4. Al-Temimi AH, Al-Hilfy IH. Effect of applying biostimulants on the agronomic parameters of corn varieties. Rev Bras Cienc Agra. 2021;16(2):1-6. [CrossRef]
5. Singh JS, Kumar A, Rai AN, Singh DP. Cyanobacteria:A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front Microbiol. 2016;7:529. [CrossRef]
6. Tuhy L, Samoraj M, Witkowska Z, Chojnacka K. Biofortification of maize with micronutrients by Spirulina. Open Chem. 2015;13(1):1119-26. [CrossRef]
7. Mógo ÁF, Ördög V, Lima GP, Mógor Z, Mogor G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. J Appl Phycol. 2018;30:453-60.[CrossRef]
8. Colla G, Nardi S, Cardarelli M, Ertani A, Lucini L, Canaguier R, et al. Protein hydrolysates as biostimulants in horticulture. Sci Hortic. 2015;196:28-38. [CrossRef]
9. Colla G, Hoagland L, Ruzzi M, Cardarelli M, Bonini P, Canaguier R, et al. Biostimulant action of protein hydrolysates:Unraveling their effects on plant physiology and microbiome. Front Plant Sci. 2017;8:2202. [CrossRef]
10. Calvo P, Nelson L, Kloepper JW. Agricultural uses of plant biostimulants. Plant Soil. 2014;383:3-41. [CrossRef]
11. Staller JE. Maize Cobs and Cultures:History of Zea MaysL. London, New York USA:Springer Heidelberg Dordrecht;2010. 262. [CrossRef]
12. Agri-food and Fisheries Information Service. AFFIS);2019. Available from: https://infosiap.siap.gob.mx:8080/ agricola_siapgobmx/avancenacionalsinprograma/do;jsessionid=55a99094657f225a0d5566211981209
13. Suárez RF, Chavez LA, Mariscal AG. Importance of Mexico's native in the national diet:An indispensable review. Rev Fitotec Mex. 2013;36 (Suppl 3-A):275-83. [CrossRef]
14. Salinas-Moreno Y, Pérez-Alonso JJ, Vázquez-Carrillo G, Aragón Cuevas F, Velázquez-Cardelas GA. Anthocyanins and antioxidant activity in maize (Zea mays L.) the chalqueño, elotes conicos and bolita breeds. Agrociencia. 2012;46(7):693-706.
15. Petroni K, Pilu R, Tonelli C. Anthocyanins in corn:A wealth of genes for human health. Planta. 2014;240:901-11. [CrossRef]
16. Pedreschi R, Cisneros-Zevallos L. Phenolic profiles of Andean purple corn (Zea mays L.). Food Chem. 2007;100(3):956-63. [CrossRef]
17. Zilic S, Serpen A, Akillioglu G, Gokmen V, Vanc J. Phenolic compounds, carotenoids, anthocyanins, and antioxidant capacity of colored maize (Zea mays L.) Kernels. J Agric Food Chem. 2012;60:1224-31. [CrossRef]
18. Barrientos Ramirez L, Ramírez Salcedo HE, Fernández Aulis MF, Ruiz-López MA, Navarro-Ocaña A, Vargas-Radillo JJ. Anthocyanins from rose maize (Zea mays) grains. Interciencia. 2018;43(3):188-92.
19. Khandelwal N, Abraham SK. Protective effects of common anthocyanidins against genotoxic damage induced by chemotherapeutic drugs in mice. Planta Med. 2014;80:1278-83. [CrossRef]
20. Prior RL, Wu X. Anthocyanins:Structural characteristics that result in unique metabolic patterns and biological activities. Free Radic Res. 2006;40(10):1014-28. [CrossRef]
21. Pojer E, Mattivi F, Johnson D, Stockley CS. The case for anthocyanin consumption to promote human health:A review. Compr Rev Food Sci Food Saf. 2013;12(5):483-508. [CrossRef]
22. Zafra-Stone S, Yasmin T, Bagchi M, Chatterjee A, Vinson JA, Bagchi D. Berry anthocyanins as novel antioxidants in human health and disease prevention. Mol Nutr Food Res. 2007;51(6):675-83. [CrossRef]
23. Miguel MG. Anthocyanins:Antioxidant and/or anti-inflammatory activities. J Appl Pharm Sci. 2011;1:7-15.
24. Tena N, Martín J, Asuero AG. State of the art of anthocyanins:Antioxidant activity, sources, bioavailability, and therapeutic effect in human health. Antioxidants (Basel). 2020;9(5):451. [CrossRef]
25. Khoo HE, Azlan A, Tang ST, Lim SM. Anthocyanidins and anthocyanins:Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr Res. 2017;61(1):1361779. [CrossRef]
26. Herrera-Sotero M, González-Cortés F, García-Galindo H, Juarez-Aguilar E, Rodríguez Dorantes M, Chávez-Servia J, et al. Anthocyanin profile of red maize native from mixteco race and their antiproliferative activity on cell line DU145. In:Justino GC, editor. Flavonoids - from Biosynthesis to Human Health. London:InTech Open;2017. 484. [CrossRef]
27. Fernández-Rodriguez VE, Ruiz-López MA. Polyphenol content, antioxidant capacity and toxicity of Solanum ferrugineum (Solanaceae) with medicinal potential. Acta Biol Colomb. 2021;26(3):414-22. [CrossRef]
28. Giusti MM, Wrolstad RE. Characterization and measurement of anthocyanins by UV-visible spectroscopy. In:Wrolstad RE, Acree TE, Decker EA, Penner MH, Reid DS, Schwartz SJ, et al., . Current Protocols in Food Analytical Chemistry. New York:John Wiley and Sons Inc.;2001. p. F1.2.1-13. [CrossRef]
29. Rodriguez-Salinas PA, Zavala-García F, Urías-Orona V, Muy-Rangel D, Heredia JB, Niño-Medina G. Chromatic, nutritional and nutraceutical properties of pigmented native maize (Zea mays L.) from the Northeast of Mexico. Arab J Sci Eng. 2020;45:95-112. [CrossRef]
30. Harakotr B, Suriharn B, Tangwongchai R, Scott MP, Lertrat K. Anthocyanin, phenolics and antioxidant activity changes in purple waxy corn as affected by traditional cooking. Food Chem. 2014;164:510-7. [CrossRef]
31. Francavilla A, Joye IJ. Anthocyanins in whole grain cereals and their potential effect on health. Nutrients. 2020;12(10):2922. [CrossRef]
32. López-Martínez LX, Oliart-Ros RM, Valerio-Alfaro G, Lee CH, Parkin KL, Garcia HS. Antioxidant activity, phenolic compounds and anthocyanins content of eighteen strains of Mexican maize. LWT Food Sci Technol. 2009;42(6):1187-92.[CrossRef]
33. Mairuae N, Palachai N, Noisa P. An anthocyanin-rich extract from Zea mays L. . ceratina alleviates neuronal cell death caused by hydrogen peroxide-induced cytotoxicity in SH-SY5Y cells. BMC Complement Med Ther. 2024;24:162. [CrossRef]
34. Elisa DH, Marcela GM, Janet Alejandra GU, Elena DH. The nutraceutical value of maize (Zea mays L.) landraces and the determinants of its variability:A review. J Cereal Sci. 2022;103:103399.[CrossRef]
35. Martínez-Martínez R, Vera-Guzman AM, Chavez-Servia JL, Bolanos EN, Carrillo-Rodríguez JC, Perez-Herrera A. Bioactive compounds and antioxidant activities in pigmented maize landraces. Interciencia. 2019;44:549-56.
36. Sithole N, Gupta S, Dube Z, Ogbe A, Van Staden J. Algae and cyanobacteria-based biostimulants in controlling plant-parasitic nematodes:A sustainable approach for crop protection. Phytoparasitica. 2023;51:803-13. [CrossRef]
37. Abdel-Aal J, Young JC, Rabalski I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J Agric Food Chem. 2006;54(13):4696-4704. [CrossRef]
38. Moreira V, Stanquevis R, Amaral EP, Lajolo FM, Hassimotto NM. Anthocyanins from purple maize (Zea mays L.) lipopolysaccharide-induced peritonitis in mice by modulating the myD88 signaling pathway. Pharma Nutr. 2021;16:100265. [CrossRef]
39. Yang Z, Zhai W. Identification and antioxidant activity of anthocyanins extracted from the seed and cob of purple corn (Zea mays L.). Innov Food Sci Emerg Technol. 2010;11(1):169-76.[CrossRef]
40. Baraud J, Genevois L, Panart JP. Anthocyanins of corn. J Agric Trop Bot Appl. 1974;11:55-9.[CrossRef]
41. Ahmad S, Chen J, Chen G, Huang J, Zhou Y, Zhao K, et al. Why black flowers?An extreme environment and molecular perspective of black color accumulation in the ornamental and food crops. Front Plant Sci. 2022;13:885176. [CrossRef]
42. Charles DJ. Antioxidant Properties of Spices, Herbs and Other Sources. Vol. 9., Ch. 2. New York:Springer Science+Business Media;2013. [CrossRef]
43. Gulcin I. Antioxidants and antioxidant methods:An updated overview. Arch Toxicol. 2020;94:651-715. [CrossRef]
44. Kumar B. Plant bio-regulators for enhancing grain yield and quality of legumes:A review. Agric Rev. 2020;42(2):175-82. [CrossRef]
45. Mannino G, Gentile C, Ertani A, Serio G, Bertea CM. Anthocyanins:Biosynthesis, distribution, ecological role, and use of biostimulants to increase their content in plant foods-a review. Agriculture. 2021;11:212. [CrossRef]
46. Holton'TA, Cornish EC. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell. 1995;7:1071-83.[CrossRef]
47. Colombo R, Ferron L, Papetti A. Colored corn:An up-date on metabolites extraction, health implication, and potential use. Molecules. 2021;26:199. [CrossRef]
48. Farruggia D, Di Miceli G, Licata M, Leto C, Salamone F, Novak J. Foliar application of various biostimulants produces contrasting response on yield, essential oil and chemical properties of organically grown sage (Salvia officinalis L.). Front Plant Sci. 2024;15:1397489. [CrossRef]
49. Nivetha N, Shukla PS, Nori SS, Kumar S, Suryanarayan S. A red seaweed Kappaphycus alvarezii-based biostimulant (AgroGain®) improves the growth of Zea mays and impacts agricultural sustainability by beneficially priming rhizosphere soil microbial community. Front Microbiol. 2024;15:1330237. [CrossRef]
50. Shukla PS, Prithiviraj B. Ascophyllum nodosum biostimulant improves the growth of Zea mays grown under phosphorus impoverished conditions. Front Plant Sci. 2021;11:601843. [CrossRef]
51. Kocira A, Swieca M, Kocira S, Zlotek U, Jakubczyk A. Enhancement of yield, nutritional and nutraceutical properties of two common bean cultivars following the application of seaweed extract (Ecklonia maxima). Saudi J Biol Sci. 2018;25(3):563-71. [CrossRef]
52. Papenfus HB, Stirk WA, Finnie JF, Van Staden J. Seasonal variation in the polyamines of Ecklonia maxima. Bot Mar. 2012;55(5):539-46. [CrossRef]