
1. INTRODUCTION
Sustainable agriculture is necessary to produce enough food to feed the world's expanding population. Biological control is often considered a system that ensures most of the benefits of sustainable agriculture, ensuring economic and environmental sustainability. Additionally, new secondary metabolites are always needed to battle antibiotic-resistant bacteria, fungal infections, and viral disorders due to the emergence of pathogenic microorganisms that are resistant to numerous medications [1,2]. This emphasizes how critical it is to find novel, potent bioactive substances with enhanced modes of action.
Bacteria are known to produce a diverse range of biologically active compounds with diverse applications, including their use as antibiotics for treating diseases, as well as demonstrating anticancer and neuroprotective properties. Beyond their significant pharmaceutical potential, bioactive compounds synthesized by plant-associated microorganisms, particularly endophytes, play a crucial role in enhancing plant growth and inducing systemic resistance to protect plants against pathogen invasion [3]. The ability of bacteria to produce antimicrobial substances is often linked to their secondary metabolism, which can be activated under specific environmental conditions or through microbial interactions. This capacity has been harnessed to identify new antibiotics to combat drug-resistant pathogens, as well as to develop sustainable biological solutions for agricultural pest control. Therefore, these beneficial bacteria represent promising candidates for the discovery of bioactive compounds, including novel antibiotics and pest management products.
Microorganisms known as endophytes reside in plant tissues and frequently function as symbionts [4], and they have attracted interest because of their many bioactive metabolites against pathogens, particularly those found in medicinal plants of ethnopharmacological value [5]. These endophytes generate various metabolites, such as biomass, vital enzymes, alkaloids, flavonoids, phenolic acids, quinones, steroids, and terpenoids, all of which have distinctive structures and important biological activities [6]. Endophytes produce secondary metabolites such as siderophores and volatile organic compounds that support nutrient acquisition, promote plant growth, and suppress pathogens, making them vital components of the plant microbiome. Their role in enhancing plant health and resilience is critical for sustainable agriculture and ecological adaptation to changing climatic conditions [7].
However, finding and creating bioactive secondary metabolites in artificially regulated environments is a challenging task. The controlled processes that make up biosynthetic gene clusters (BGCs) in bacteria are essential for their growth and ability to compete in the environment. BGCs are organized groups of genes within bacterial genomes that encode a variety of biosynthetic enzymes, regulatory proteins, and sometimes macromolecules. These components are crucial for the production of secondary metabolites, which often provide resistance to the final natural product. The expression of BGCs is tightly regulated, allowing bacteria to conserve energy by producing secondary metabolites only when necessary for growth or ecological competition. Innovative techniques are required to generate quiet or poorly expressed BGCs because most culturable bacteria do not express their biosynthetic potential outside their native environment. This is difficult to get natural products under laboratory circumstances. Thus, it is clear that new and creative approaches must be used to stimulate the expression of silent and/or poorly expressed BGCs to generate novel biologically active natural compounds under carefully monitored circumstances [8].
It has been demonstrated that changes to growth media, both chemically and physically, can affect the transcription rate of certain genes and induce BGC expression, which would not normally occur in laboratory cultures [9]. These modifications may include co-cultivation with competitors, including precursors, using compounds that function as selective or universal epigenetic elicitors, or changes to the growth media or growth conditions [10]. For example, the synthesis of two extra antibiotics by P. fluorescens HV37a against P. ultimum when cultivated on PA with endogenous glucose levels less than 0.06% was reviewed by Keswani et al. [11]. In terms of the co-culture strategy, Peng et al. [12] reviewed the identification of two novel polycyclic tetramate macrolactams produced by co-culturing the mycolic acid-containing bacterium Tsukamurella pulmonis TP-B0596 with Umezawaea sp. RD066910.
Elicitors, either naturally occurring or synthetically produced chemical compounds, are vital in enhancing systemic resistance in plants. These compounds may originate from microbial sources or result from specific chemical combinations. Depending on their origin, elicitors are classified into biotic, abiotic, biological, chemical, or physical categories, with biotic elicitors specifically referring to those derived from microorganisms. Microbial elicitors not only activate plant immune responses but also aid in the transmission of signaling molecules that further amplify the plant's defense mechanisms. Microbial biocontrol agents utilize these elicitors to combat plant diseases by employing various modes of action, making them valuable tools in agricultural disease management. The potential of elicitors to improve crop resilience and yield has drawn significant attention in agricultural research. Notably, some studies have reported that elicitors can boost plant resistance by up to 85%. A prominent example is chitin (chitooligosaccharide), a well-established fungal elicitor that functions as a pathogenicity molecule [13].
Bacillus velezensis has been widely recognized for its ability to promote plant growth, produce antimicrobial and antifungal substances, and enhance stress tolerance, highlighting its potential for diverse applications in agricultural and pharmaceutical aspects [14,15]. In particular, Bacillus velezensis in this study derived from a wide medicinal plant (Adenosma bracteosum) offers unique advantages compared to other bacteria due to their co-evolutionary relationship with host plants. These bacteria often exhibit enhanced abilities to produce bioactive compounds, such as antimicrobial, antifungal, and antioxidant substances, contributing to the plant's defense mechanisms [16,17]. This specialization makes them valuable sources for discovering novel secondary metabolites with pharmaceutical and agricultural applications.
Most previous studies have primarily focused on utilizing the biomass of beneficial bacteria, such as live or inactivated cells, rather than exploring their secondary metabolites. On the other hand, the exploration of bacterial secondary metabolites has gained attention for their potential in the pharmaceutical and agricultural sectors. These metabolites, including antibiotics and pigments, have been recognized for their antimicrobial, antiviral, antioxidant, and antitumoral properties [18]. Furthermore, the structural diversity of secondary metabolites offers a wide range of biotechnological applications across health, industrial, and environmental sectors [19]. Despite this potential, the exploration and utilization of secondary metabolites have been comparatively under-researched, highlighting the need for further studies in this area to unlock their full potential. Therefore, in this study, we pay attention to the effects of abiotic and especially biotic stress using a new approach co-culture on the production of metabolites in a symbiotic bacterial strain isolated from the Vietnamese medicinal plant (A. bracteosum Bonati). This approach has significant potential for discovering new bioactive compounds, including antibiotics, antifungals, and other therapeutic agents, as well as advancing our understanding of microbial ecology and stress responses.
Hence, building on our previous findings that endogenous bacteria in A. bracteosum Bonati exhibit antibacterial properties, this study aimed to explore the influences of different elicitors on the synthesis of antibacterial substances by B. velezensis strain SB1R13.2 against Escherichia coli, Staphylococcus aureus, and Aeromonas hydrophila and provide a basis for future applications in biological control.
2. MATERIALS AND METHODS
2.1. Endophytic Strain and Culture Condition
The endophytic strain (identified B. velezensis strain SB1R13.2) was previously isolated from wild populations of A. bracteosum Bonati from Lo Go National Park in Vietnam. This strain exhibited the highest antibacterial activity among isolated bacteria from A. bracteosum Bonati in the previous examination, so it is chosen for further investigations. The strain was stored in 20% glycerol at the Institute of Biotechnology and Food - Can Tho University under −20°C. The endophytic strain was regularly cultured on Yeast Extract Mannitol (YEM) growth medium, and incubated at 28°C ± 2°C for 48–72 hours.
2.2. Extracellular and Intracellular Fluids are Prepared to Investigate Cell Growth and Antibacterial Activity
Preparation of extracellular fluids was performed based on the protocol outlined by Tabbene et al. [20], with modifications: Enriched endogenous bacteria with YEM medium, then adjusted the density to 107 CFU/mL. The bacterial suspension (1 ml) was centrifuged at 12,000 rpm at 4°C for 15 minutes. The residue was removed, and the supernatant (without cells) was collected and filtered through a 0.45 µm membrane filter.
Preparation of intracellular fluids followed the methodology described by Parniakov et al. [21] with several modifications: endogenous bacteria were enriched with YEM medium, then adjust the density to 107 CFU/ml. Bacterial fluid (1 ml) was centrifuged at 12,000 rpm at 4°C for 10 minutes. The supernatant was removed, and the bacterial biomass was obtained. This biomass was dissolved in 1 ml of physiological saline, and a 40 kHz ultrasound at 30°C was applied for 30 minutes to break down bacterial cells. The mixture was then centrifuged at 12,000 rpm at 4°C for 10 minutes, and the supernatant was collected as the bacterial intracellular fluid.
The antibacterial activity of intracellular and extracellular fluids of endogenous bacteria against E. coli, S. aureus, and A. hydrophila was conducted using the agar disk diffusion method. Briefly, 6 mm sterile cotton tips were used to create wells on the agar surface, and 50 µl of the endogenous bacterial culture solution at 24 hours was pipetted into each well. The plates were then incubated at 30°C, and the diameter of the antibacterial ring was observed after 24 hours. The positive control used tetracycline at 10 mg/ml, while the negative control was 0.85% sterile physiological saline. The experiment was arranged randomly and repeated three times. The antibacterial ring diameter was calculated using the formula described by Huemer et al. [22]:
Antibacterial diameter (mm) = D - d
where D is the diameter of the bacterial inhibition zone (mm), and d, equal to 6 mm, is the diameter of the well.
2.3. Effects of Medium Components and Culture Conditions of SB1R13.2 Strain on Their Antibacterial Activity
The bacterial culture solution of SB1R13.2 was prepared by growing the bacteria overnight in 10 ml of YEM medium at 30°C on a rotary shaker at 150 rpm. A 250 ml Erlenmeyer flask with 50 ml of YEM medium supplemented with different nutrients as listed below was inoculated with 107 CFU/ml of endophytic bacteria. Incubation was performed at 30°C for 24 hours [20]. Cell growth was measured using an optical density spectrophotometer set to 600 nm.
Effects of medium composition on the growth and the antibacterial activity of bacteria: SB1R13.2 bacterial suspension (100 µl) was cultivated in falcon tubes containing 20 ml of YEM medium supplemented with carbon sources at 1.5% (w/v) from glucose, fructose, lactose, sucrose, and starch to determine the optimal carbon source for antibacterial activity and cell development. After adding each of these carbon sources separately to the medium, mannitol was extracted. Nitrogen sources including peptone, beef extract, yeast extract, NH4Cl, NH4NO3, and (NH4)2SO4 were supplemented separately at a final concentration of 0.15% (w/v). The impacts of inorganic salts including 2 g/l ZnSO4, 0.15 mg/l MnSO4, 0.135 g/l FeCl3, and 0.88 g/l CaCl2 were also investigated. The initial pH of different supplemented media was adjusted to pH 7.0 before use. All experiments were carried out in triplicate.
Influence of culture conditions on cell growth and antibacterial activity production: Six incubation times 0, 24, 36, 48, 60, and 72 hours were used to establish the optimal incubation period for the antibacterial activity of endophytic bacteria. The other parameters were 30°C, pH 7.0, and 1% inoculation ratio. In terms of pH conditions, 5.0, 6.0, 7.0, and 8.0 were employed and the media were incubated for 48 hours at 30°C with the initial inoculation proportion at 1%. Finally, to ascertain an appropriate temperature, the experiment was conducted using a 1% initial inoculation percentage, a pH of 7.0, and five distinct temperatures 30°C, 37.5°C, 45°C, and 52.5°C applied.
2.4. Impact of Bio-Elicitor (pathogens bacteria) Concentration on the Antibacterial Capability of an Endogenous Bacterial Strain
The experiment was performed according to the method described by Mahmoud, et al. [23] with some modifications. The YEM medium was used to cultivate both pathogenic (107 CFU/ml) and endogenous (107 CFU/ml) bacteria. The study entailed co-culturing pathogenic and endophytic bacteria to culture a suspension of endophytic bacteria enriched with elicitors. In the YEM medium, the proportion of pathogenic to endogenous bacteria changed at 1%, 2%, 3%, 4%, 5%, and 6%. Each type of pathogenic bacteria served as an individual stimulating factor, and a mixture of pathogenic bacteria was not introduced into the culture solution containing the elicitor. The three repetitions of the experiment were set up at random.
2.5. Isolation and Purification of Antibacterial Agents in Co-Culture With Pathogen
The antibiosis experiment involved co-cultivation of the SB1R13.2 strain with the best bio-elicitor in 500 ml of YEM medium (ratio 1:0.1) at 30°C for 3 days with agitation at 120 rpm. All cells were collected by centrifuging at 5,000 rpm at 4°C for 10 minutes. The cell-free supernatant from the treatment was added to an equal volume of ethyl acetate solvent (1:1). The mixture was mixed by vigorous shaking for 30 minutes and allowed to settle. The organic solvent phase was collected and concentrated at 40°C under vacuum, using a rotary evaporator model (IKA, HB10 basic). The ethyl acetate extract was used for Gas chromatography-mass spectrometry (GC-MS) analysis. Methanol is used to elute extract before injecting to GC-MS.
2.6. GC-MS Identification and Characterization
The ethyl acetate extract was analyzed by GC-MS using a Model QP2020NX instrument, Shimadzu, Japan equipped with a splitless injector, a Shimadzu AOC 20i Plus autosampler, and a Shimadzu Rxi 5MS heated silica column (phenyl-methylpolysiloxane 5%, 30 m length, 0.25 mm i.d., 0.25 mm thickness). GC conditions included programmed heating from 50°C to 300°C at 10°C/minute, followed by 10 minutes at 300°C. The injection volume was 1 µl and the injection was maintained at 280°C. The carrier gas was helium at 1.0 ml/minute−1. The MS conditions: electron impact ion source temperature, 230°C; ionization energy, 70 eV; quadrupole temperature, 150°C; scan rate, 3.2 scans/s; mass range, 50–1,000 m/z. Compounds were identified based on their mass spectral matches and retention indices with the NIST/Wiley 275 library (Wiley, New York, USA). The relative abundance of each feature was calculated from the total ion chromatogram by computer [24].
2.7. Statistical Analyses
These data are stored, calculated, and presented in graph form using Microsoft Excel 2016 software. Analysis of significant differences between experimental samples is performed using the one-factor ANOVA statistical method (α = 5%) using Minitab 16.
3. RESULTS AND DISCUSSION
3.1. Effects of Carbon, Nitrogen Sources, and Metal Ions on Cell Growth and Antibacterial Activity of SB1R13.2
The effects of nitrogen sources, carbon sources, and inorganic salts on cell growth and the production of antibacterial activity in intracellular and extracellular fluids of the SB1R13.2 strain were cultured using a YEM medium with added the differences of carbon, nitrogen, and inorganic salts sources. As shown in Figure 1, the SB1R13.2 was grown on various carbon sources. Mannitol supported the highest bacterial growth (OD = 0.677), showing significant differences at the 5% level compared to other carbon sources (Fig. 1A). Similar to the carbon source, the SB1R13.2 strain was also grown on a variety of nitrogen sources, both organic and inorganic (Fig. 1B). Ammonium chloride was found to be an excellent nitrogen source for bacterial growth (OD = 1.461), significantly different from other nitrogen sources at the 5% level. Additionally, ammonium nitrate also stimulated bacterial growth (OD = 1.385).
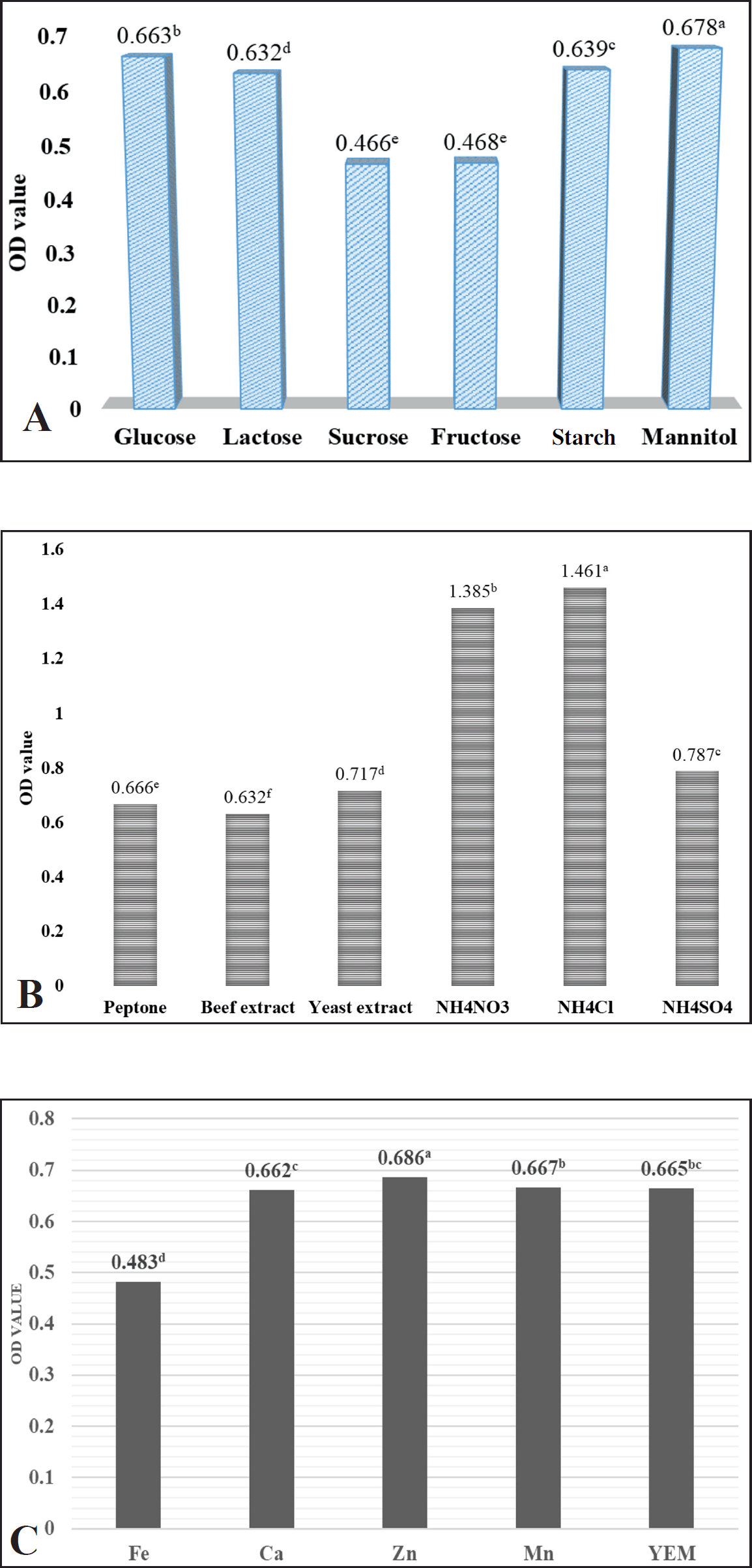 | Figure 1. Influence of the composition of the medium on the growth of SB1R13.2 strain. A: Effect of carbon sources on the growth of SB1R13.2; B: Effect of nitrogen sources on the growth of SB1R13.2; C: Effect of metal ions source on the growth of SB1R13.2; Mean values with the same letters indicate statistically insignificant differences at the 5% significance level according to Tukey's test. [Click here to view] |
Carbon sources serve as both energy providers and the foundation for the carbon skeletons of microbial cells and their metabolites [25]. Nitrogen sources supply essential nitrogen for the synthesis of bacterial proteins, enzymes, nucleic acids, and other critical biological macromolecules required by microorganisms [26]. Metal ions also play a significant role in microbial growth, reproduction, and enzyme activity, as well as their yield. Elements such as calcium, chlorine, sodium, and zinc help maintain optimal osmotic pressure and redox potential for microorganisms. Furthermore, certain metal ions are integral components of enzymes and act as activators in enzymatic reactions. However, some metal ions can inactivate enzymes by binding to thiol groups, disrupting disulfide bonds, or replacing native metal ions in the enzyme structure [27,28]. Hence, selecting appropriate carbon sources, nitrogen sources, and metal ions is essential. Ojha et al. [29] reported that the optimal growth of Bacillus aryabhattai KIIT BE-1 was achieved when starch was used as the carbon source in the growth medium. In contrast, the findings of Karrays et al. [30] demonstrated that Bacillus stearothermophilus exhibited similar growth rates with various carbon sources, including xylose, glucose, starch, lactose, and galactose, with the exception of casein, which supported lower growth.
Interestingly, while the SB1R13.2 strain can utilize a variety of carbon, nitrogen, and mineral sources for cell growth, the antibacterial activity of intracellular and extracellular fluids against the pathogen was not produced in this strain. Singh et al. [31] reported that the composition and concentration of the carbon and nitrogen sources could exert a decisive influence on the production of antimicrobial compounds. For instance, the inhibition of Xanthomonas sp. growth was improved when fructose and yeast extract were used as the carbon and nitrogen sources in the culture medium of B. velezensis 8–2. In another study, Hu et al. [32] depicted the improvement of antifungal substances in B. velezensis LZN01 by optimizing the fermentation condition. In this study, D-fructose and NH4Cl were the best carbon and nitrogen sources in the culture medium to obtain the highest inhibition rate. However, these results are consistent with the results of [33] showing the repression of antibiotic activity of Brevibacillus laterosporus EA62 by wheat bran, whereas a high growth rate was observed when using wheat bran as a carbon source. Similarly, no antibacterial activity was observed in the presence of the nitrogen sources used in this investigation. This means that there was no direct relationship between the growth of the organism and antibiotic formation. In addition, the secondary metabolite pathways of different microorganisms are very different. Therefore, further investigation including using other carbon and nitrogen sources or the combination of these sources to enhance the antibacterial activity of SB1R13.2.
Inorganic ions play an important role in the growth of Bacillus, such as cell composition and energy transfer. Parallelly, bacterial SB1R13.2 was grown on inorganic salts (Fig. 1C). The highest cell growth (OD = 0.686) was observed with the addition of 0.2 g/l ZnSO4, significantly different from other inorganic salts at the 5% level. The addition of Fe ions inhibited SB1R13.2 cell growth compared to treatments without metal ions (OD = 0.483). In contrast, Pan et al. [34] exhibited the highest promotive effect of Fe2+, followed by Al3+ on the growth of Bacillus thuringiensis. Regarding the production of antibacterial compounds, when MgSO4+CaCO3 were used as metal ion sources in the culture medium, Brevibacillus laterosporus EA62 displayed the highest antibacterial activity [33].
3.2. Effects of pH, Temperature, and Incubation Time on Cell Growth and Antibacterial Activity of SB1R13.2
The effects of pH, temperature, and incubation time on cell growth and the production of antibacterial activity in intracellular and extracellular fluids of the SB1R13.2 strain were cultured using the YEM medium with the differences in pH, temperature, and incubation time. The results of Figure 2A demonstrated that the SB1R13.2 strain was grown on various pH. The optimal pH for SB1R13.2 growth was pH 7.0, significantly differing from other pH levels at the 5% significance level. pH levels higher or lower than this hindered bacterial growth. Furthermore, the inoculation temperature and time affected on bacterial growth of the SB1R13.2 as depicted in Figures 2B and 2C. The best incubation temperature for SB1R13.2 growth was 30°C, significantly differing from other temperatures at the 5% significance level. Elevated temperatures suppressed bacterial growth, with complete inhibition observed at 52.5°C. Besides that SB1R13.2 strain displayed substantial growth starting from 72 hours with significant differences observed at the 5% significance level compared to other times. Culture temperature, pH, and incubation time are the essential factors influencing bacterial growth [30,35,36]. For example, B. stearothermophilus exhibited maximum growth at 55°C, pH 10, and at 24 hours after inoculation [30].
Conversely, the ability of this strain to produce intracellular and extracellular products against the pathogen bacteria was not observed when grown alone in the YEM medium under any SB1R13.2 culture conditions. This result is in agreement with the study of [37] indicated less significant effects of incubation time and pH on the production of bioactive compounds produced by B. velezensis strain RP137. In contrast, using the appropriate pH contributed to the production of antifungal substances by B. velezensis LZN01 [32]. In addition, the best antibiotic activity of B. laterosporus EA62 was obtained at 37°C and pH 7.0, and after 72 hours of incubation [33].
In conclusion, this study observed an increase in bacterial biomass under optimal conditions; however, this was not accompanied by a corresponding enhancement in antimicrobial activity. This phenomenon may be attributed to the uncoupling of antibacterial substance production from bacterial growth, as antimicrobial synthesis appeared to occur predominantly during the stationary phase or under resting cell conditions [38]. Furthermore, stress factors, such as nutrient limitations or specific environmental signals, are known to induce secondary metabolism. In nutrient-rich environments, bacteria may not perceive sufficient stress to initiate the production of antimicrobial compounds.
3.3. Effect of Biostimulants on the Growth and Antibacterial Capacity of SB1R13.2 Strain
In the experiment assessing antibacterial activity without using elicitors (pathogens bacteria), the intracellular fluid and extracellular fluid of the endogenous bacterial strain SB1R13.2 did not exhibit a clear zone when tested against pathogenic bacteria (Fig. 3). For experiments evaluating antibacterial activity using elicitors: The intracellular fluid of SB1R13.2 did not exhibit a clear zone at any concentration of elicitors. However, the extracellular fluid of SB1R13.2 formed a clear zone.
The absence of antibacterial activity in the experiment conducted without elicitors may be attributed to the quiescent or low expression of BGCs in SB1R13.2 under standard culture conditions [8]. Co-culturing has been shown to activate BGCs that remain silent under standard laboratory conditions [39]. These genes are transcribed in response to the induced stress conditions created during co-culture, leading to the production of cryptic compounds that are not detectable in axenic cultures of the producing strain. This response is likely a survival strategy triggered by competition or antagonism in the co-culture environment [40]. This mechanism may explain the observed antibacterial activity when SB1R13.2 was co-cultured with pathogenic bacteria. The results of this study, which demonstrate the production of antibacterial compounds by SB1R13.2 during co-culture with E. coli, A. hydrophila, and S. aureus, are consistent with findings from previous research [41,42].
 | Figure 2. Effect of culture conditions on the growth of SB1R13.2 strain. A: Effect of pH value on the growth of SB1R13.2; B: Effect of incubation temperature on the growth of SB1R13.2; C: Effect of incubation time on the growth of SB1R13.2. This inhibitory effect of Fe ions is likely attributed to the oxidative stress induced by Fe ions through the Fenton reaction, which generates reactive oxygen species (ROS) [75]. These ROS can damage cellular components, including DNA, proteins, and lipids, ultimately impairing cell viability and growth [76]. Mean values with the same letters indicate statistically insignificant differences at the 5% significance level according to Tukey's test. [Click here to view] |
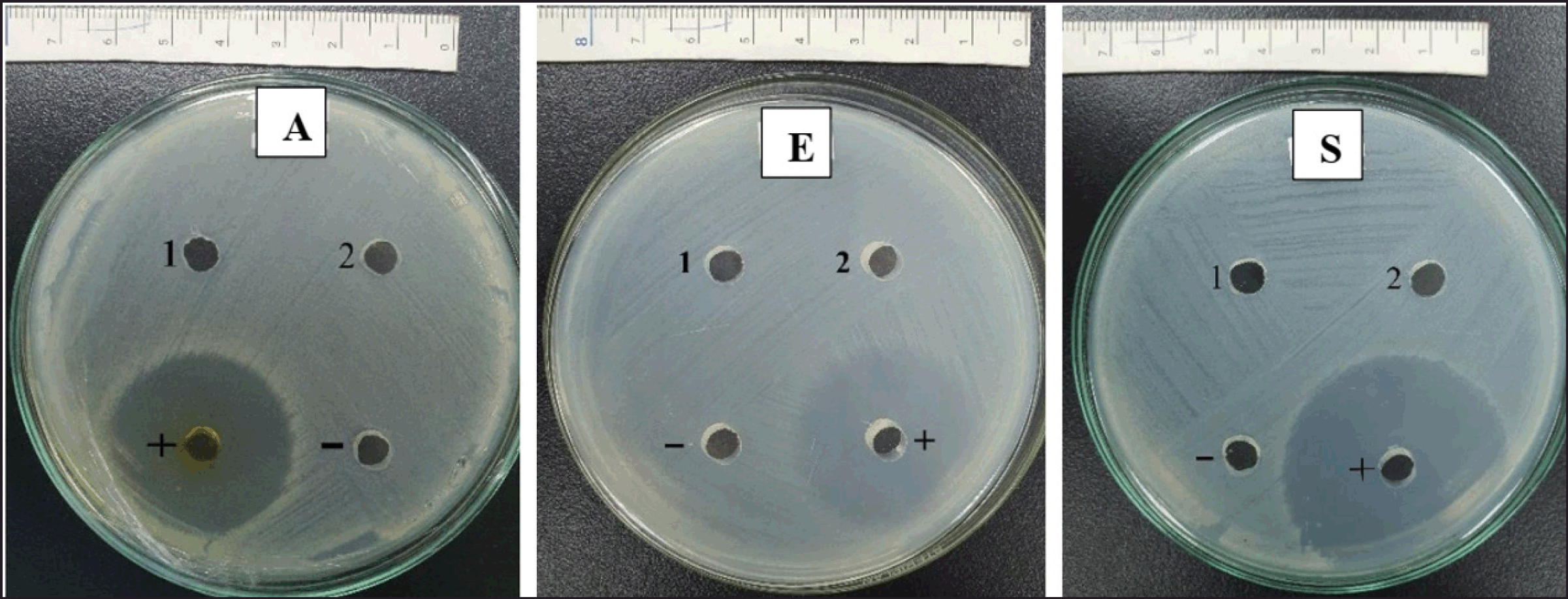 | Figure 3. Antibacterial activity of extracellular and intracellular fluid of SB1R13.2 against pathogens. A: Aeromonas hydrophila; E: Escherichia coli; S: Staphylococcus aureus; 1: Extracellular fluid; 2: Intracellular fluid; (+) positive control; (–) negative control. [Click here to view] |
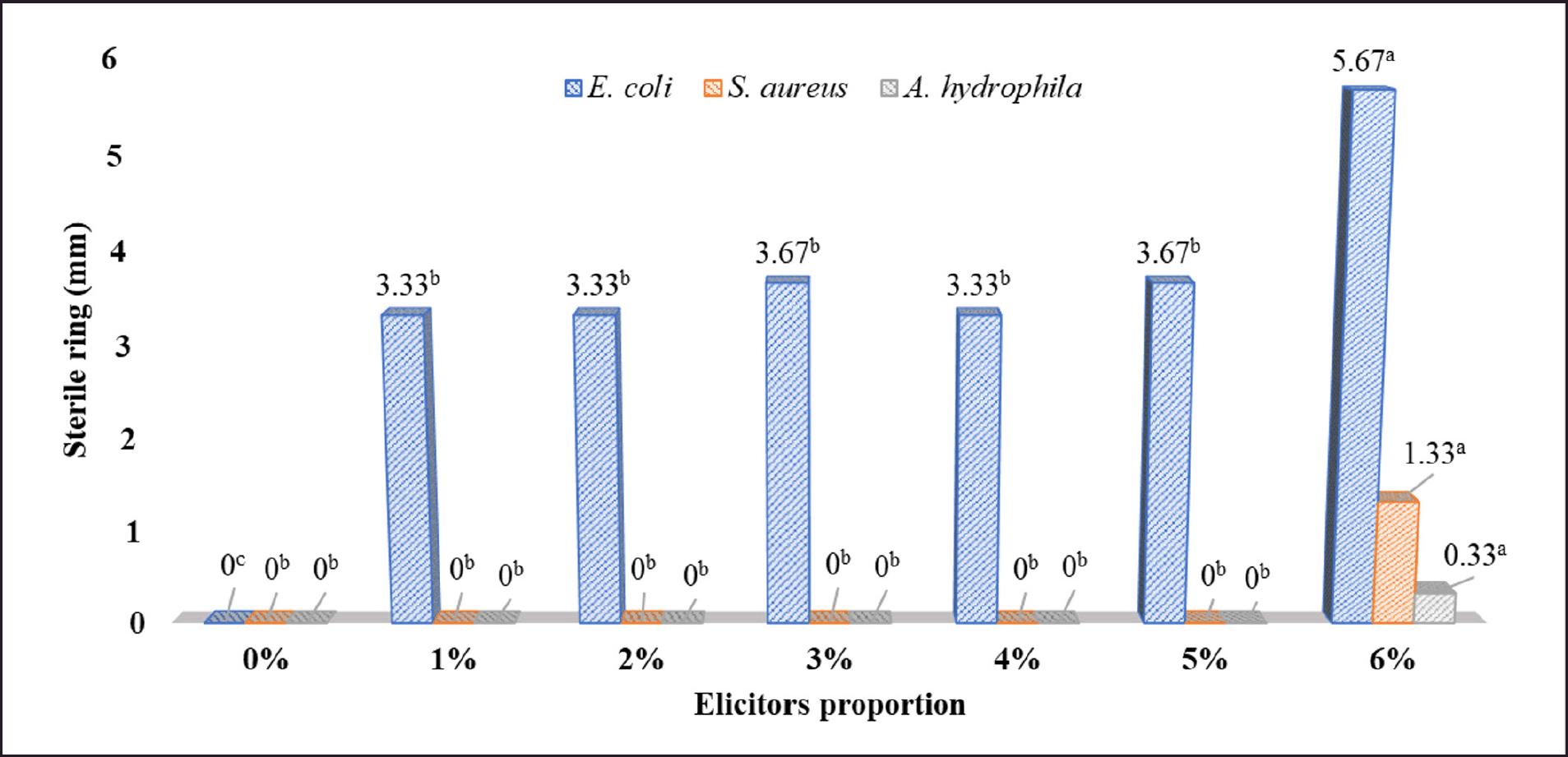 | Figure 4. Antibacterial extracellular products secreted from SB1R13.2 while co-cultured with different elicitors. Mean values with the same letters indicate statistically insignificant differences at the 5% significance level according to Tukey's test for each elicitor proportion. [Click here to view] |
The results shown in Figure 4 show that the SB1R13.2 strain demonstrates the ability to produce antibacterial extracellular products when stimulated at different densities corresponding to different pathogenic bacterial strains. The anti-E. coli activity was assessed using the extracellular of the co-culture solution that was produced after the original inoculation of E. coli with a density range of 106 to 6 × 106 CFU/ml. There was no difference in the diameters of inhibition zones formed between the treatment using E. coli with the density at 106 CFU/ml and 5 × 106 CFU/ml. However, when the density of the elicitor increased to 6 × 106 CFU/ml, the inhibition zones rose to 5.67 mm. Notably, with the concentration of 6 × 106 CFU/ml for A. hydrophila and S. aureus elicitors, the co-culture extracellular solution of SB1R13.2 A. hydrophila and S. aureus exhibited the antibacterial activity, with the inhibition zones of 0.33 mm and 1.33 mm, respectively. This suggests that the use of elicitors at the density of 6 × 106 CFU/ml triggers the strain SB1R13.2 to secrete antibacterial products.
In addition, the response of SB1R13.2 to exhibit higher antibacterial activity at elevated density of elicitors (6 × 106 CFU/ml) might be attributed to the higher accumulation of metabolites produced by the co-cultured pathogen. The production of secondary metabolites in one organism is often initiated in response to sensing secondary metabolites produced by another organism [43]. An increased concentration of these metabolites has been shown to enhance antimicrobial activity [44]. This underscores the importance of signaling molecule or elicitor concentrations exceeding a critical threshold to effectively induce the production of secondary metabolites in co-culture systems. This process is frequently associated with quorum sensing or stress signaling mechanisms, both of which rely on the concentration of elicitors to activate specific biosynthetic pathways.
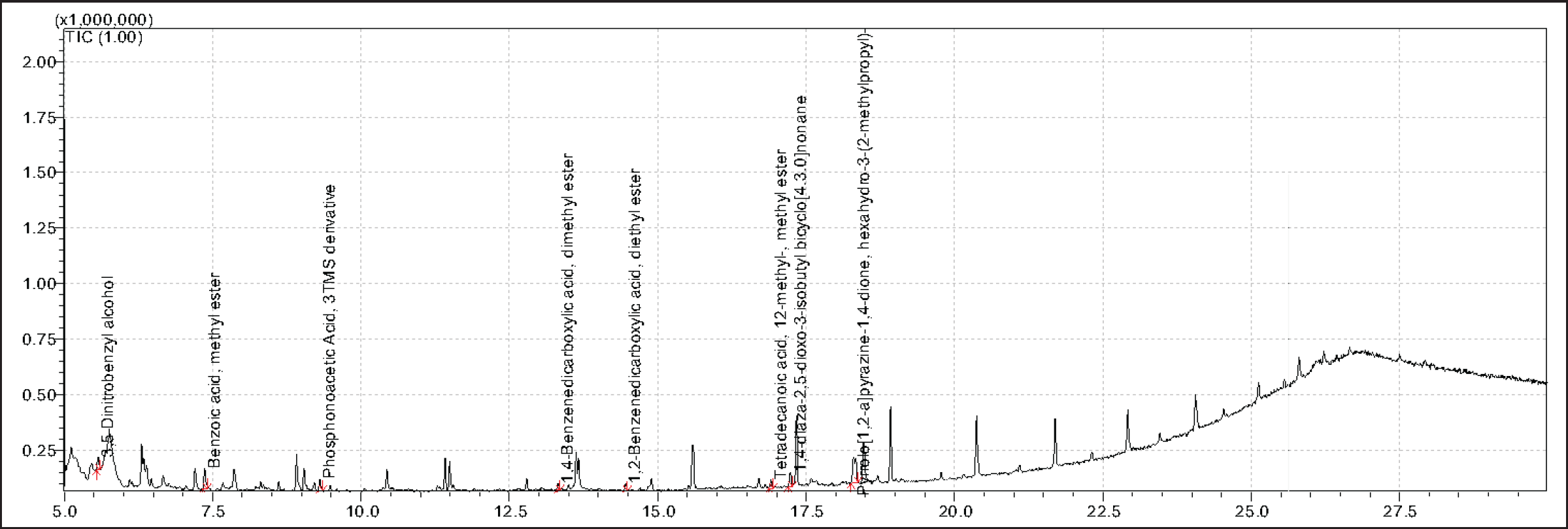 | Figure 5. GC-MS chromatogram of SB1R13.2 extract co-cultured with E. Coli. [Click here to view] |
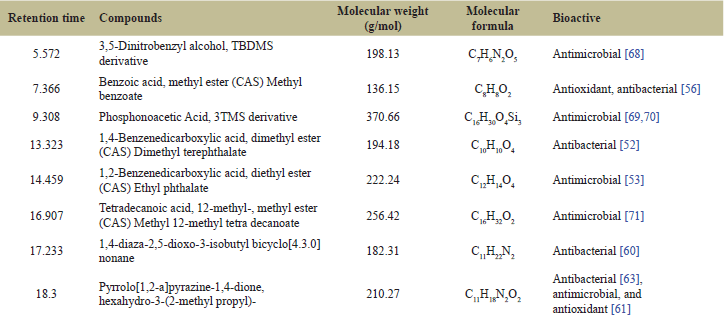 | Table 1. Bioactive chemical compounds identified in the ethyl acetate extract of co-culture SB1R13.2 with E. Coli. [Click here to view] |
The bacterial strain SB1R13.2 exhibited potent antibacterial activity when co-cultured with E. coli, S. aureus, and A. hydrophila. SB1R13.2 demonstrated strong resistance to E. coli, with low bio-elicitor concentrations stimulating the production of extracellular antibacterial substances and forming a large zone of inhibition. This finding aligns with studies showing that lactic acid bacteria isolated from Ori cereal can reduce drug-resistant E. coli populations compared to controls [45]. Additionally, Lacticaseibacillus rhamnosus GG and Bifidobacterium lactic strains completely inhibited E. coli growth during 24 hours of co-culture [46].
Recently, a novel strategy to boost metabolite production in plants has been implemented through the use of elicitors [47]. Elicitors can either be abiotically derived from non-living sources or biotically derived from living sources. This study demonstrates that co-cultivating endogenous bacteria in medicinal plants with pathogenic bacteria (bio-elicitors) also exerts significant impacts on the formation of biologically active compounds.
3.4. Phytochemical Profiling by GC-MS Analysis
To identify the predominant organic compounds produced by the highly bioactive SB1R13.2 strain, ethyl acetate extracts of cell supernatant were concentrated from the co-culture of SB1R13.2 with E. coli. GC-MS analysis revealed the presence of various features (putative compounds). These features were tentatively identified by comparing their spectra with those in the NIST database (USA) [24,48]. The spectrum of unknown components was cross-referenced with known components stored in the NIST library [48]. Additionally, references to the biological activity of compounds discovered and published by previous authors, along with interpretations based on Phytochemical and Ethnobotanical Databases, were considered [24].
The GC-MS analysis of crude extracts from SB1R13.2 cultivated in co-culture with E. coli revealed at least eight features (Fig. 5 and Table 1). The chemical compounds identified in the ethyl acetate extract of SB1R13.2 exhibited various biological activities, such as antibacterial, antioxidant, and antimicrobial properties. The eight compounds produced in the extract using the elicitor are outlined in Table 1.
3,5-dinitrobenzyl alcohol, Benzoic acid, methyl ester (CAS) Methyl benzoate, Phosphonoacetic acid, 3TMS derivative, 1,4-Benzenedicarboxylic acid, dimethyl ester (CAS) Dimethyl terephthalate, 1,2-Benzenedicarboxylic acid, and diethyl ester (CAS) Ethyl phthalate possess antimicrobial properties [49–53]. However, there is limited information on their production by bacteria.
Whereas, Methyl benzoate (benzoic acid, methyl ester) is a naturally occurring ester found in various plants [54]. Studies have shown that methyl benzoate exhibits antimicrobial activity against certain bacterial and fungal strains, including S. aureus and Candida albicans [55], S. aureus and Pseudomonas aeruginosa [56]. Additionally, methyl benzoate has been investigated for its potential as an environmentally safe insecticide, indicating its broader antimicrobial properties [54]. The identification of methyl benzoate in the culture broth of SB1R13.2 represents the first documented instance of this antibacterial compound being produced by a bacterial strain.
Competitive interactions within co-culture systems are believed to enhance the production of valuable compounds and promote the synthesis of novel metabolites [57,58]. For example, Bacillus was able to produce its secondary metabolites, such as biofilm and surfactin lipopeptides upon perception of the siderophore pyochelin produced by a Pseudomonas competitor [59]. Another intriguing case of this phenomenon involves the identification of two novel polycyclic tetramate macrolactams produced by co-culturing the mycolic acid-containing bacterium T. pulmonis TP-B0596 with Umezawaea sp. RD066910 [25].
Besides, 1,4-Diaza-2,5-dioxo-3-isobutyl bicyclo[4.3.0]nonane detected in this study has been isolated from marine endophytic actinomycete Nocardiopsis sp. DMS 2(MH900226). This compound has demonstrated significant antibacterial and anti-biofilm activities, particularly against biofilm-forming Klebsiella pneumoniae [60]. Finally, Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl) is a bioactive compound produced by various bacterial species including Bacillus tequilensis MSI45 [61], Actinomycetia strain ANP14ARS [62], and Kocuria sp. SRS88 [63] exhibits notable antibacterial properties. These findings suggest that the antibacterial activity of SB1R13.2 may be attributed to the bioactive compounds identified. This underscores the potential of harnessing various beneficial metabolites through the cultivation of the bacterial strain SB1R13.2.
The relationship between endophytic bacterial phytochemicals and phytopathogen control has been observed in various studies. For instance, phytochemicals from endophytic Stenotrophomonas maltophilia, Azotobacter chroococcum S11, S. marcescens S14, and Bacillus sp. SV81 has been implicated in controlling Fusarium oxysporum in Solanaceae plants [64]. Similarly, the phytochemicals of Pseudomonas putida BP25, endophytic in black pepper, showed the potential to suppress various pathogens [65,66]. Rhizobacterial phytochemicals, classified into different chemical classes, have also been associated with disease control [67].
In conclusion, the antagonistic activities of extracellular products produced by symbiotic bacteria can be effectively utilized to improve the health and growth of A. bracteosum and other plant species. These effects are achieved through various mechanisms, including the suppression of plant pathogens, the induction of systemic resistance, and the inhibition of competitive microorganisms within the rhizosphere. Furthermore, these extracellular products contribute to improved nutrient availability, stimulate plant growth through the production of phytohormones, and enhance the ability of plants to tolerate abiotic stresses such as drought and salinity. Harnessing these bioactive compounds facilitates the development of bio-inoculants or biostimulants, offering a sustainable approach to promoting plant health, reducing disease incidence, and advancing agricultural productivity.
4. CONCLUSION
The SB1R13.2 strain excreted extracellular antibacterial products when co-cultured with bio-stimulating factors (106 CFU/ml for E. coli as the elicitor, and 6 × 106 CFU/ml for S. aureus and A. hydrophila). However, the SB1R13.2 strain did not produce antibacterial products under varied growth conditions without these bio-stimulating factors. Culturing SB1R13.2 with the bio-elicitor (E. coli) led to the production of phytochemical constituents, with eight compounds, including known antimicrobial agents, identified through GC-MS analysis. This study suggests that SB1R13.2 is a promising antibacterial agent and provides new insights into its phytochemical constituent production.
In conclusion, co-culturing not only enhances the production of secondary metabolites through microbial interactions but also mimics natural ecological processes, providing a more sustainable and environmentally friendly alternative to chemical fertilizers and pesticides. Additionally, co-culture systems promote the discovery of novel bioactive compounds by activating silent biosynthetic pathways, which are often not expressed in monocultures. These methods can also improve microbial community stability, resilience under abiotic stresses, and nutrient cycling efficiency, further supporting their potential in sustainable agriculture and biotechnology. Co-culture methods also provide a more realistic platform to study microbial ecology, revealing symbiotic, commensal, or antagonistic relationships. Additionally, they enhance antimicrobial activity, reduce development time and costs, and support biotransformation processes by leveraging the combined enzymatic potential of the participating organisms. By emphasizing these broader implications, the argument for leveraging co-culturing as a bio-stimulant approach becomes more compelling and aligns with the goals of sustainable and innovative agricultural practices.
While co-culture methods hold significant promise for enhancing secondary metabolite production and fostering microbial interactions, they are not without limitations that may impact the reliability and reproducibility of results. A primary challenge lies in the specificity of bio-elicitors, as strain SB1R13.2 exhibits variable responses to different elicitors. Consequently, the findings of this study are reliable and reproducible only when E. coli, S. aureus, and A. hydrophila are used to induce the antibacterial activity of strain SB1R13.2. Furthermore, the production of secondary metabolites in co-cultures is highly sensitive to minor variations in factors such as inoculum size, nutrient availability, pH, and temperature. These variations can result in inconsistencies across different experiments or laboratories, as demonstrated by the dose-dependency experiments in this study. These findings underscore the need for future optimization of culture conditions to ensure consistent and efficient production of antibacterial substances in this research.
The findings in this research highlight the real-time application potential of EPs in sustainable agriculture, including their use for pathogen suppression, stress mitigation, and ecological restoration, particularly in the face of environmental challenges posed by climate change. In addition, SB1R13.2 holds potential as a promising source for metabolic production, with applications spanning sustainable agriculture, veterinary practices, and pharmaceuticals. Further studies are warranted, including the isolation and quantification of individual bioactive compounds present in the co-culture broth of SB1R13.2 and pathogenic bacteria. Additionally, it is essential to investigate the impact of environmental stresses on the production of these compounds to optimize their use for large-scale industrial applications.
5. ACKNOWLEDGMENTS
The authors would like to thank the colleges that facilitated the work in their laboratories.
6. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. FINANCIAL SUPPORT AND SPONSORSHIP
There is no financial support or sponsorship.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All the data are available with the authors and shall be provided upon request.
11. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing the manuscript, and no images were manipulated using AI.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors, and the reviewers. This journal remains neutral about jurisdictional claims in published institutional affiliation.
REFERENCES
1. Andryukov B, Mikhailov V, Besednova N. The biotechnological potential of secondary metabolites from marine bacteria. J Marine Sci Eng 2019;7:176.
2. Seukep AJ, Kuete V, Nahar L, Sarker SD, Guo M. Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J Pharm Anal 2020;10:277–90.
3. Haruna A, Yahaya, SM. Yahaya, recent advances in the chemistry of bioactive compounds from plants and soil microbes: a review. Chem Afri 2021;4:231–48.
4. Bilal S, Ali L, Khan AL, Shahzad R, Asaf S, Imran M, et al. Endophytic fungus Paecilomyces formosus LHL10 produces sester-terpenoid YW3548 and cyclic peptide that inhibit urease and α-glucosidase enzyme activities. Arch Microbiol 2018;200:1493–502.
5. Toghueo RM, Zabalgogeazcoa I, de Aldana BV, Boyom FF. Enzymatic activity of endophytic fungi from the medicinal plants Terminalia catappa, Terminalia mantaly and Cananga odorata. South Afri J Bot 2017;109:146–53.
6. Bhoonobtong A, Sawadsitang S, Sodngam S, Mongkolthanaruk W. Characterization of endophytic bacteria, Bacillus amyloliquefaciens for antimicrobial agents production. Int Proc Chem Biol Environ Eng 2012;40:6–11.
7. Qadir M, Iqbal A, Hussain A, Hussain A, Shah F, Yun BW, et al. Exploring plant–bacterial symbiosis for eco-friendly agriculture and enhanced resilience. Int J Mol Sci 2024;25:12198.
8. de Felício R, Ballone P, Bazzano CF, Alves LF, Sigrist R, Infante GP, et al. Chemical elicitors induce rare bioactive secondary metabolites in deep-sea bacteria under laboratory conditions. Metabolites 2021;11:107.
9. Zhang X, Elliot MA. Unlocking the trove of metabolic treasures: activating silent biosynthetic gene clusters in bacteria and fungi. Curr Opin Microbiol 2019;51:9–15.
10. Tomm HA, Ucciferri L, Ross AC. Advances in microbial culturing conditions to activate silent biosynthetic gene clusters for novel metabolite production. J Indus Microbiol Biotechnol 2019;46:1381–400.
11. Keswani C, Singh HB, García-Estrada C, Caradus J, He YW, Mezaache-Aichour S, et al. Antimicrobial secondary metabolites from agriculturally important bacteria as next-generation pesticides. Appl Microbiol Biotechnol 2020;104:1013–34.
12. Peng XY, Wu JT, Shao CL, Li ZY, Chen M, Wang CY. Co-culture: stimulate the metabolic potential and explore the molecular diversity of natural products from microorganisms. Marine Life Sci Technol 2021;3:1–12.
13. Meena M, Yadav G, Sonigra P, Nagda A, Mehta T, Swapnil P, et al. Role of elicitors to initiate the induction of systemic resistance in plants to biotic stress. Plant Stress 2022;5:100103.
14. Khalid F, Khalid A, Fu Y, Hu Q, Zheng Y, Khan S, et al. Potential of Bacillus velezensis as a probiotic in animal feed: a review. J Microbiol 2021;59:627–33.
15. Sun X, Xu Z, Xie J, Hesselberg-Thomsen V, Tan T, Zheng D, et al. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J 2022;16:774–87.
16. Vaou N, Stavropoulou E, Voidarou C, Tsakris Z, Rozos G, Tsigalou C, et al. Interactions between medical plant-derived bioactive compounds: focus on antimicrobial combination effects. Antibiotics 2022;11:1014.
17. Nguyen TD, Nguyen TT, Pham MN, Duong HN, Pham TT, Nguyen TP, et al. Relationships between endophytic bacteria and medicinal plants on bioactive compounds production. Rhizosphere 2023;27:100720.
18. Fouillaud M, Dufossé L. Microbial secondary metabolism and biotechnology. Microorganisms 2022;10:123.
19. Bodade R, Kumar A, Sharma VK, Angelini P. Recent advances in biotechnological applications of microbial secondary metabolites. Front Microbiol 2024;15:1525146.
20. Tabbene O, Slimene IB, Djebali K, Mangoni ML, Urdaci MC, Limam F. Optimization of medium composition for the production of antimicrobial activity by Bacillus subtilis B38. Biotechnol Prog 2009;25:1267–74.
21. Parniakov O, Apicella E, Koubaa M, Barba FJ, Grimi N, Lebovka N, et al. Ultrasound-assisted green solvent extraction of high-added value compounds from microalgae Nannochloropsis spp. Bioresour Technol 2015;198:262–7.
22. Huemer M, Mairpady Shambat S, Brugger SD, Zinkernagel ASAntibiotic resistance and persistence—Implications for human health and treatment perspectives. EMBO Rep 2020;21:e51034.
23. Mahmoud ST, Luti KJ, Yonis RW. Enhancement of prodigiosin production by Serratia marcescens S23 via introducing microbial elicitor cells into culture medium. Iraqi J Sci 2015;56:1938–51.
24. Mohamad OA, Li L, Ma JB, Hatab S, Xu L, Guo JW, et al. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front Microbiol 2018;9:924.
25. Li Y, Tang X, Chen L, Xu X, Li J. Characterization of a nattokinase from the newly isolated bile salt-resistant Bacillus mojavensis LY-06. Foods 2022;11:2403.
26. Wu R, Chen G, Pan S, Zeng J, Liang Z. Cost-effective fibrinolytic enzyme production by Bacillus subtilis WR350 using medium supplemented with corn steep powder and sucrose. Sci Rep 2019;9:6824.
27. Yao Z, Kim JA, Kim JH. Characterization of a fibrinolytic enzyme secreted by Bacillus velezensis BS2 isolated from sea squirt jeotgal. Food Microbiol Biotechnol 2019;29:347–56.
28. Xin X, Ambati RR, Cai Z, Lei B. Purification and characterization of fibrinolytic enzyme from a bacterium isolated from soil. 3 Biotech 2018;8:1–8.
29. Ojha SK, Singh PK, Mishra S, Pattnaik R, Dixit S, Verma SK. Response surface methodology based optimization and scale-up production of amylase from a novel bacterial strain, Bacillus aryabhattai KIIT BE-1. Biotechnol Rep 2020;27:e00506.
30. Karray A, Alonazi M, Horchani H, Ben Bacha A. A novel thermostable and alkaline protease produced from Bacillus stearothermophilus isolated from olive oil mill sols suitable to industrial biotechnology. Molecules 2021;26:1139.
31. Singh V, Haque S, Niwas R, Srivastava A, Pasupuleti M, Tripathi C. Strategies for fermentation medium optimization: an in-depth review. Frontn Microbiol 2017;7:2087.
32. Hu J, Wang Z, Xu W. Production-optimized fermentation of antifungal compounds by Bacillus velezensis LZN01 and transcriptome analysis. Microb Biotechnol 2024;17:e70026.
33. Ak AU, Demirkan E, Cengiz M, Sevgi T, Zeren B, Abdou M. Optimization of culture medium for the production and partial purification and characterization of an antibacterial activity from Brevibacillus laterosporus strain EA62. Rom Biotechnol Lett 2019;24:705–13.
34. Pan X, Huang T, Fang Y, Rao W, Guo X, Nie D, et al. Effect of Bacillus thuringiensis biomass and insecticidal activity by cultivation with vegetable wastes. Royal Soc Open Sci 2021;8:201564.
35. Yao M, Ma C, Bian X, Yang Y, Xu Y, Wu Q, et al. Isolation and optimal fermentation conditions of Bacillus licheniformis SFD-Y5 for a New Douchi fibrinolytic enzyme producer. Fermentation 2023;9:668.
36. Nayarisseri A, Singh P, Singh SK. Screening, isolation and characterization of biosurfactant producing Bacillus subtilis strain ANSKLAB03. Bioinformation 2018;14:304.
37. Pournejati R, Karbalaei-Heidari HR. Optimization of fermentation conditions to enhance cytotoxic metabolites production by Bacillus velezensis strain RP137 from the Persian Gulf. Avicenna J Med Biotechnol 2020;12:116.
38. Cappelletti M, Presentato A, Piacenza E, Firrincieli A, Turner RJ, Zannoni D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl Microbiol Biotechnol 2020;104:8567–94.
39. Tan ZQ, Leow HY, Lee DC, Karisnan K, Song AA, Mai CW, et al. Co-culture systems for the production of secondary metabolites: current and future prospects. Open Biotechnol J 2019;13:13–8.
40. Ka AK, Keerthi TR. Co-culture as the novel approach for drug discovery from marine environment. Nov Appro Drug Des Dev 2017;2:1–4.
41. Vinale F, Nicoletti R, Borrelli F, Mangoni A, Parisi OA, Marra R, et al. Co-culture of plant beneficial microbes as source of bioactive metabolites. Sci Rep 2017;7:14330.
42. Moody SC. Microbial co-culture: harnessing intermicrobial signaling for the production of novel antimicrobials. Fut Microbiol 2014;9:575–8.
43. Ueda K, Beppu T. Antibiotics in microbial coculture. J Antibiot 2017;70:361–5.
44. Seyedsayamdost MR. High-throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc Natl Acad Sci 2014;111:7266–71.
45. Afolayan AO, Ayeni FA.Antagonistic effects of three lactic acid bacterial strains isolated from Nigerian indigenous fermented Ogi on E. coli EKT004 in co-culture. Acta Aliment 2017;46:1–8.
46. Kathayat D, Closs G, Helmy YA, Deblais L, Srivastava V, Rajashekara In vitro and in vivo evaluation of Lacticaseibacillus rhamnosus GG and Bifidobacterium lactis Bb12 against avian pathogenic Escherichia coli and identification of novel probiotic-derived bioactive peptides. Probiotics Antimicrob Proteins 2022;14:1–17.
47. Bhaskar R, Xavier LS, Udayakumaran G, Kumar DS, Venkatesh R, Nagella P. Biotic elicitors: a boon for the in-vitro production of plant secondary metabolites. Plant Cell Tissue Organ Cul 2022;149:7–24.
48. Maulidia V, Sriwati R, Soesanto L, Syamsuddin, Hamaguchi T, Hasegawa K. Endophytic bacteria isolated from higher plant in Aceh, Indonesia, and their chemical compounds activity against Fusarium oxysporum f. sp. lycopersici. Egypt J Biol Pest Control 2021;31:1–7.
49. Karabanovich G, Fabiánová V, Vocat A, Dusek J, Valaskova L, Stolarikova J, et al. Both nitro groups are essential for high antitubercular activity of 3, 5-dinitrobenzylsulfanyl tetrazoles and 1, 3, 4-oxadiazoles through the deazaflavin-dependent nitroreductase activation pathway. J Med Chem 2023;67:81–109.
50. Yan D, Li Y, Liu Y, Li N, Zhang X, Yan C. Antimicrobial properties of chitosan and chitosan derivatives in the treatment of enteric infections. Molecules 2021;26:7136.
51. Freestone TS, Ju KS, Wang B, Zhao H. Discovery of a phosphonoacetic acid derived natural product by pathway refactoring. ACS Synth Biol 2017;6:217–23.
52. Totaro G, Sisti L, Cionci NB, Martinez GA, Di Gioia D, Celli A. Elastomeric/antibacterial properties in novel random Ricinus communis based-copolyesters. Polym Test 2020;90:106719.
53. Premjanu N, Jaynthy C. Antimicrobial activity of diethyl phthalate: an in silico approach. Asian J Pharm Clin Res 2014;7:141–2.
54. Mostafiz MM, Hassan E, Lee KY. Methyl benzoate as a promising, environmentally safe insecticide: current status and future perspectives. Agriculture 2022;12:378.
55. Mohanapriya S, Muthukumaran M, Vairam S. Synthesis, characterization, thermal behavior and antimicrobial activity of 3-methyl benzoate complexes of transition metal with hydrazine. Bull Chem Soc Ethiopia 2016;30:241–52.
56. Alemdar A, Tan B, Toksöz O, Kurtulu? G, Sesal C, Odaba? Z. Systematically investigation on the spectral, antioxidant and antibacterial properties of fragrant methyl benzoate esters containing electron withdrawing and electron releasing groups. J Mol Struct 2023;1291:136100.
57. Granato ET, Meiller-Legrand TA, Foster KR. The evolution and ecology of bacterial warfare. Curr Biol 2019;29:R521–37.
58. Granato ET, Foster KR. The evolution of mass cell suicide in bacterial warfare. Curr Biol 2020;30:2836–43.
59. Andri? S, Rigolet A, Argüelles Arias A, Steels S, Hoff G, Balleux G, et al. Plant-associated Bacillus mobilizes its secondary metabolites upon perception of the siderophore pyochelin produced by a Pseudomonas competitor. ISME J 2023;17:263–75.
60. Rajivgandhi GN, Ramachandran G, Kanisha CC, Li JL, Yin L, Manoharan N, et al. Anti-biofilm compound of 1, 4-diaza-2, 5-dioxo-3-isobutyl bicyclo [4.3. 0] nonane from marine Nocardiopsis sp. DMS 2 (MH900226) against biofilm forming K. pneumoniae. J King Saud Univ Sci 2020;32:3495–502.
61. Kiran GS, Priyadharsini S, Sajayan A, Ravindran A, Selvin J. An antibiotic agent pyrrolo [1, 2-a] pyrazine-1, 4-dione, hexahydro isolated from a marine bacteria Bacillus tequilensis MSI45 effectively controls multi-drug resistant Staphylococcus aureus. RSC Adv 2018;8:17837–46.
62. Konwar AN, Basak S, Gurumayum S, Borah JC, Thakur D. Exploring northeast India’s culturable soil Actinomycetia for potent antibacterial agents against gram-positive bacterial pathogens of clinical importance. Sci Rep 2024;14:28640.
63. Sahadevan R, devi Sureshan S, Paskaran P. Pyrrolo (1, 2-A) Pyrazine-1, 4-Dione, Hexahydro-3-(2-Methyl propyl)-and Phenol, 2, 4,-Bis (1, 1-Dimethyl ethyl) novel antibacterial metabolites from a marine Kocuria sp. SRS88: optimization and its application in medical cotton gauze cloth against bacterial wound pathogens. Inter J Pharm Res Dev 2014;6:44–55.
64. Aydi-Ben-Abdallah R, Jabnoun-Khiareddine H, Daami-Remadi M. Fusarium wilt biocontrol and tomato growth stimulation, using endophytic bacteria naturally associated with Solanum sodomaeum and S. bonariense plants. Egypt J Biol Pest Control 2020;30:1–13.
65. Agisha VN, Kumar A, Eapen SJ, Sheoran N, Suseelabhai R. Broad-spectrum antimicrobial activity of volatile organic compounds from endophytic Pseudomonas putida BP25 against diverse plant pathogens. Biocontrol Sci Technol 2019;29:1069–89.
66. Patel A, Kumar A, Sheoran N, Kumar M, Sahu KP, Ganeshan P, et al. Antifungal and defense elicitor activities of pyrazines identified in endophytic Pseudomonas putida BP25 against fungal blast incited by Magnaporthe oryzae in rice. J Plant Dis Protect 2021;128:261–72.
67. Audrain B, Farag MA, Ryu CM, Ghigo JM. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol Rev 2015;39:222–33.
68. Akbar A, Gul Z, Aziz S, Sadiq MB, Achakzai JK, Saeed S, Chein SH. Bio-functional potential and biochemical properties of propolis collected from different regions of Balochistan Province of Pakistan. Oxid Med Cell Longev 2022;2022:7585406.
69. Kesk?n D, Ceyhan N, U?ur A, Dbeys AD. Antimicrobial activity and chemical constitutions of West Anatolian olive (Olea europaea L.) leaves. J Food Agric Environ 2012;10:99–102.
70. Russell AD. Types of antibiotics and synthetic antimicrobial agents. Hugo Russell's Pharm Microbiol 2004;10:152–86.
71. Dib MA, Bendahou M, Bendiabdellah A, Djabou N, Allali H, Tabti B, et al. Partial chemical composition and antimicrobial activity of Daucus crinitus Desf. extracts. Grasas y Aceites 2010;61:271–8.
72. Khachatryan A, Vardanyan N, Vardanyan A, Zhang R, Castro L. The effect of metal ions on the growth and ferrous iron oxidation by Leptospirillum ferriphilum CC isolated from Armenia mine sites. Metals 2021;11:425.
73. Sun HQ, Lu XM, Gao PJ. The exploration of the antibacterial mechanism of Fe3+ against bacteria. Brazil J Microbiol 2011;42:410–4.
74. Hansen CT, Kleint C, Böhnke S, Klose L, Adam-Beyer N, Sass K, et al. Impact of high Fe-concentrations on microbial community structure and dissolved organics in hydrothermal plumes: an experimental study. Sci Rep 2022;12:20723.
75. Jomova K, Raptova R, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol 2023;97:2499–574.
76. Guo W, Xing Y, Luo X, Li F, Ren M, Liang Y. Reactive oxygen species: a crosslink between plant and human eukaryotic cell systems. Int J Mol Sci 2023;24:13052.