
1. INTRODUCTION
Thermophiles have unique metabolic capabilities and enzymes adapted to function in extreme conditions [1]. Therefore, these organisms serve as a valuable source of stable proteins, enabling microorganisms to conserve energy and nutrient resources that would otherwise be expended on protein degradation and synthesis [2]. Subsequently, an extensive exploration of Geobacillus and Parageobacillus transcriptomes and secretomes has shown many proteins with either proven or potential industrial and medicinal applications [3,4], such as bacteriocins or BLIS [5,6]. Bacteriocins are ribosomally synthesized antimicrobial peptides (AMPs) produced by bacteria to inhibit other bacteria from establishing predominance in environmental microbial communities [7]. AMPs have emerged as highly promising alternatives to antibiotics due to their potential in treating bacterial infections, including those caused by multidrug-resistant pathogens [8]. They have also been shown to have low or no cytotoxicity on eukaryotic cell lines even when used at very high doses [9].
Numerous AMPs with distinct activity spectra and mechanisms of action have been characterized [8], such as toebicin 218, thermoleovorins, and termosin, which are produced by Geobacillus toebii [10], Geobacillus thermoleovorans [11], and Geobacillus stearothermophilus [12], respectively. In the study by Alkhalili [13], Geobacillus sp. has a source of novel antibacterial peptides different from various other types. The AMPs secreted by Geobacillus thermodenitrificans, Geobacillus ZGt-1, and Geobacillus spp. are known to effectively inhibit pathogenic bacteria, Staphylococcus aureus, Salmonella enterica serovar Typhimurium, and Klebsiella pneumoniae, respectively [2,14,15].
Currently, immobilized microorganism technology is increasingly used for producing AMP. This technique is a broad term used to describe the physical trapment of viable microbial cells within a specific, defined area (polymer). This confinement restricts their free movement and imparts hydrodynamic properties that are distinct from those of the surrounding environment, thereby increasing fermentor cell density and stabilizing continuous or repeated cultures [16]. This, in turn, enhances the productivity of various bacteriocins [17,18]. It is also susceptible to the effects of inhibitory compounds, which reduce the risk of cell damage and contamination [19]. Naghmouchi et al. [20] investigated the highest production of pediocin PA-1 obtained during repeated-cycle batch cultures using Pediococcus acidatili UL5 immobilized in k-carrageenan beads. This repeated-cycle batch culture process was stable and resulted in high pediocin PA-1 production (4096 AU/mL) and volumetric productivity (342 AU/mL/h) at the end of 0.75- and 2-h incubation cycles.
The thermophilic bacterial isolate, Geobacillus subterraneus Tm6Sp1, from Mount Kamojang Crater, West Java, Indonesia, seems to be a promising source of AMPs, which was confirmed by the coding gene’s availability [21]. The focus of this study is on G. subterraneus and its potential for producing antibacterial substances against various pathogenic bacteria utilizing an immobilized cell system.
2. MATERIALS AND METHODS
2.1. Microorganisms and Maintenance of Culture
G. subterraneus Tm6Sp1 isolate was obtained from Mount Kamojang Crater, West Java, Indonesia. In a previous study, Tm6Sp1 was identified and found to have more than 99% similarity to G. subterraneus (accession no. CP051162.1) [21]. Furthermore, it was maintained from glycerol at 4°C to Mueller-Hinton Agar (MHA) at 55°C for 24-48 h. The test indicator organisms were Streptococcus mutans ATCC 700610, S. aureus ATCC 29213, S. enterica serovar Typhimurium ATCC 49416, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, and K. pneumoniae ATCC 700603. They were grown on Tryptic Soya Agar at 37°C for 24 h.
2.2. Inoculum Preparation and Cell Count
G. subterraneus Tm6Sp1 culture was grown in Luria Bertani (LB) broth at 55°C for 24–48 h in an incubator shaker at 150 rpm [22]. The growth phase of bacteria was determined with two replications by measuring the number of cells in McFarland turbidity value converted to Colony form unit (CFU)/mL (this conversion referred to Dalynn Biological [23]). This observation of the cell number of bacteria was carried out every 2 h for 36 h. Furthermore, the supernatant was taken after reaching the late logarithmic phase as the initial inoculum for producing bacteriocins [15].
2.3. Repeated Batch Production of Antibacterial Substances by Free Cells (F)
The inoculum from a fresh culture of G. subterraneus Tm6Sp1 was prepared by adjusting to the 0,5 McFarland turbidity standard (equivalent to 1×108 CFU/mL) and then diluting it 1/10 (v/v). The resulting suspension was used as the starter inoculum, which was transferred to 40 mL of modified Mueller-Hinton Broth (MHB) containing additional salts such as MgSO4 and CaCl2 as the production medium [16]. According to the growth curves, this suspension was incubated with batch fermentation in an incubator shaker at 55°C and 150 rpm until it reached the late logarithmic phase, which occurred at the 18th h. The culture broth was then centrifuged at 10000× g for 20 min at 4°C (Eppendorf centrifuge 5424 R). This centrifugation step separated the cells in the pellet for further treatment.
For the free cells (F) treatment, the pellet was resuspended in 75 mL of fresh MHB medium and subjected to repeated fermentation. As for the immobilized cell treatment, the pellet was used to prepare immobilized cells as described in Section 2.5. The supernatant obtained from the centrifugation step was filtered using 0.22 μm syringe filters to remove bacterial cells. The fourth batch of fermentation was performed to obtain data at four different observation times.
F1: First batch filtrate of G. subterraneus Tm6Sp1 free cells
F2: Second batch filtrate of G. subterraneus Tm6Sp1 free cells
F3: Third batch filtrate of G. subterraneus Tm6Sp1 free cells
F4: Fourth batch filtrate of G. subterraneus Tm6Sp1 free cells
All filtrates were tested for antibacterial activity by disk diffusion method.
2.4. Repeated Batch Production of Antibacterial Substances by Immobilized Cells (I) In Agar Beads
The method of cell immobilization used in this study was based on the procedure described by Alkhalili et al. [16], with some modifications. The cell pellet obtained from centrifugation was resuspended in 5 mL of sterile MHB and mixed with 25 mL of 3,5% (w/v) sterile molten agar solution that had been pre-warmed to 60°C. This mixture was dropped into cold mineral oil using a syringe needle to form agar beads with an average diameter of 5 mm. Furthermore, the agar beads were washed thoroughly with sterile phosphate buffer saline and distilled water to remove the oil before being transferring to 75 mL of fresh, sterile MHB. The bead suspension was incubated at 55°C and 150 rpm for 18 h. After incubation, the culture broth was carefully separated from the beads by pouring it into sterile bottles. The culture broth was then centrifuged at 10,000× g for 20 min at 4°C. The resulting supernatant was filtered using a 0.2 mm syringe filter. Finally, the filtered supernatant was stored for further analysis.
The beads were transferred back to the cultivation flask with 75 mL of fresh, sterile MHB, and another cultivation cycle was conducted. The beads were further recycled up to the fourth batch to obtain four observation times.
I1: First batch filtrate of G. subterraneus Tm6Sp1 immobilized cells
I2: Second batch filtrate of G. subterraneus Tm6Sp1 immobilized cells
I3: Third batch filtrate of G. subterraneus Tm6Sp1 immobilized cells
I4: Fourth batch filtrate of G. subterraneus Tm6Sp1 immobilized cells
All filtrates were tested for antibacterial activity by the disk diffusion method.
2.5. Antibacterial Activity Test
The filtrates (F1-F4 and I1-I4) were checked for pH, which must be neutral before use, and adjusted with 5 M NaOH [24]. The filtrate was put in a vial containing sterile blank disks of 6 mm diameter, and the disks were soaked for 1 h at room temperature. After soaking, the disks were placed on the MHA plate surface, inoculated with the test bacteria, and incubated at 37°C for 24 h. The production of antibacterial substances using free and immobilized cells was compared based on their antibacterial activity in the diameter (mm) of the inhibition zone. This was interpreted as follows: diameters of the inhibition zone >20 mm, 10–20 mm, 5–10 mm, and 0–5 mm represent very strong, strong, moderate, and no inhibitory activity or weak, respectively [25].
2.6. Statistical Analysis
The data obtained from this research regarding antibacterial activity were subjected to statistical analysis using the non-parametric Kruskal-Wallis test in the Statistical Package for the Social Sciences 25th version software. A significance level of 0.05 was employed for the analysis. All analyses were done with three replications.
3. RESULTS
3.1. Growth of G. subterraneus Tm6sp1
Bacterial growth was observed every 2 h during 36 h of incubation in an MHB medium, and CFU/mL determined the number of bacteria. A graph was made between the number of bacteria and the incubation time to identify the logarithmic phase, as shown in Figure 1.
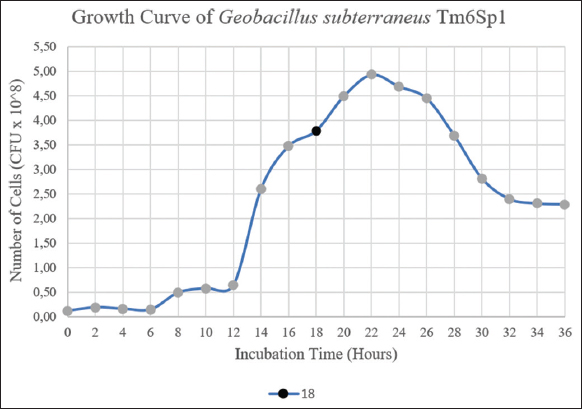 | Figure 1: Growth curves of G. subterraneus Tm6Sp1 at their optimal growth condition for 36 h. The black dot ( ) represent the time (h) of culture sampling for the first batch of free cells and the initial inoculum of immobilized cells. culture sampling for the first batch of free cells and the initial inoculum of immobilized cells. ) represent the time (h) of culture sampling for the first batch of free cells and the initial inoculum of immobilized cells. culture sampling for the first batch of free cells and the initial inoculum of immobilized cells.[Click here to view] |
Based on the analysis of growth curve parameters, it was determined that the generation time of Tm6Sp1 was relatively rapid compared to other Geobacillus strains, with an approximate value of 36.78 min. This finding is consistent with the observations made by Marchant et al. [26], who reported a doubling time of around 30 min for several Geobacillus strains isolated from soil samples in nutrient-rich media. For the production of the first batch of free cell filtrate (F1), the bacterial culture was harvested during the late logarithmic phase after 18 h of incubation.
3.2. Physical Properties of The Agar Beads With Embedded G. Subterraneus Tm6Sp1
Spherical particles immediately formed and hardened into beads when an agar solution containing embedded Tm6Sp1 was dropped into cold mineral oil. The final particles were pellucid white spheres of approximately 3–4 mm diameter, as shown in Figure 2.
 | Figure 2: Agar beads embedded with G. subterraneus Tm6Sp1 formed with 3% w/v agar bacteriological and cold mineral oil. [Click here to view] |
3.3. Antibacterial Activity by Free Cells (F)
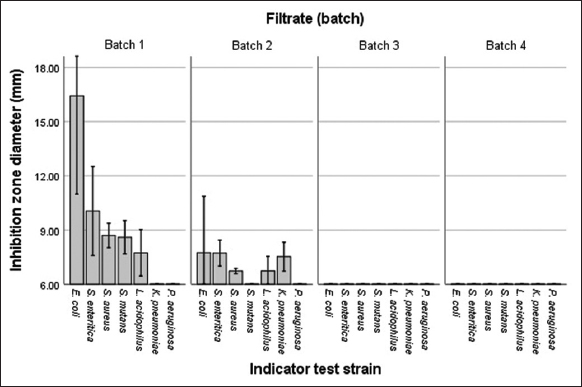
The inhibition zones can show antibacterial activity by free cells, as graphically presented in Figure 3.
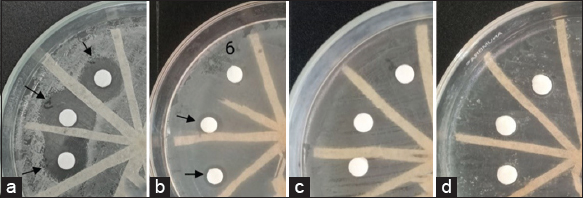 | Figure 3: Antibacterial activity by G. subterraneus Tm6Sp1 free cells against E. coli at (a) first, (b) second, (c) third, and (d) fourth batch. [Click here to view] |
The results showed that the production of antibacterial activity by free cells was at its maximum, with strong inhibition observed in the initial batch (F1) or during the final logarithmic phase before being repeated. However, upon reusing the cells, the antibacterial activity experienced a sharp decline and was not detectable in the subsequent batches (third to fourth). Overall, the inhibition zone size ranged from 8.63 mm to 16.43 mm against E. coli [Figure 4] under optimal conditions.
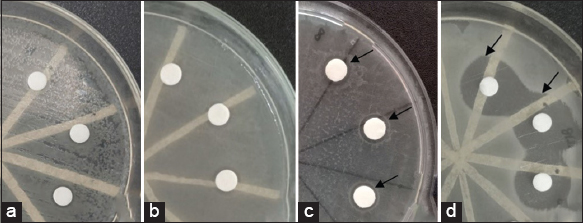 | Figure 4: Antibacterial activity by G. subterraneus Tm6Sp1 immobilized cells against E. coli at (a) first, (b) second, (c) third, and (d) fourth batch. [Click here to view] |
3.4. Antibacterial Activity by Immobilized Cells (I)
Antibacterial activity by immobilized cells can be seen by the inhibition zones formed, as shown in Figure 4.
Antibacterial production using immobilized cells exhibited maximum activity at the fourth batch (I4), inhibiting 18.04 mm against E. coli [Figure 4]. In the second batch (I2), a faint inhibition was observed against E. coli, S. mutans, and S. aureus, with inhibition zones measuring 7.05 mm, 7.23 mm, and 8.21 mm, respectively. Therefore, the production using immobilized cells increased along with the culture batch period until the fourth batch without declining [Figure 5].
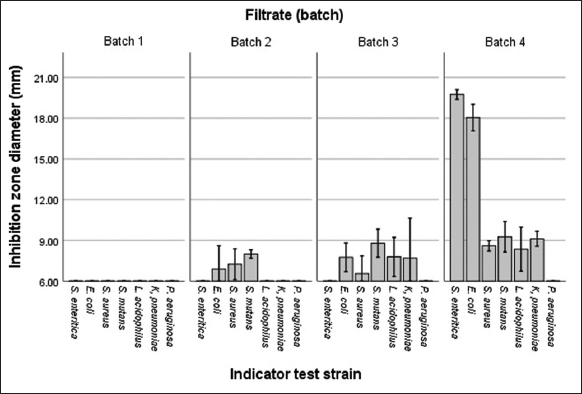 | Figure 5: Antibacterial activity by immobilized cells of Tm6Sp1 for four batches. (Very strong: >20 mm, strong: 10–20 mm, moderate: 6–10 mm, weak or not detected: <6 mm). [Click here to view] |
3.5. Antibacterial Spectrum Against Pathogenic Bacteria
Based on the optimization of batch results using both free and immobilized cells, the first and fourth batches demonstrated the most favorable conditions in terms of antimicrobial activity. Additionally, the spectrum activity was also examined against several pathogenic bacteria, as shown in Table 1.
Table 1: Antibacterial spectrum against pathogenic bacteria from an optimal batch of G. subterraneus Tm6Sp1.
| Test pathogen strain | Free cells at first batch (F1) | Immobilized cells at fourth batch (I4) |
|---|---|---|
| Gram-positive | ||
| Streptococcus mutans | - | + |
| Staphylococcus aureus | + | + |
| Lactobacillus acidophilus | + | + |
| Gram-negative | ||
| Pseudomonas aeruginosa | - | - |
| Klebsiella pneumoniae | - | + |
| Salmonella enteritica | ++ | ++ |
| Escherichia coli | ++ | ++ |
Classification of antibacterial activity, +++ (very strong): >20 mm, ++ (strong): 10–20 mm, + (moderate): 6–10 mm, - (weak or not detected): <6 mm
4. DISCUSSION
The growth phase of the G. subterraneus Tm6Sp1 isolate was determined by observing the growth curve. The results showed that the exponential phase of this strain occurred between 12 and 18 h of cultivation, as depicted in Figure 1. To obtain the first batch of free cell filtrate (F1), the culture was taken during the late logarithmic phase, specifically after 18 h of incubation. During this phase, the supernatant was filtered, resulting in the filtrate containing Tm6Sp1-free cells from the first batch. On the other hand, the centrifuged pellets were utilized in two different treatments, followed by repeated batch fermentation using the first batch as the initial inoculum to manufacture beads into immobilized cells.
The late logarithmic phase has been identified as the optimal time for the synthesis of antibacterial compounds in many bacteriocins produced by Bacilliaceae. Several studies reported that maximum production occurs during the mid-log phase in certain strains such as B. lentus, B. mycoides, and L. brevis [27]. Similarly, the production of thermosin 10, thuricin 439, and cerein 7 from G. stearothermophilus 32A [11], B. thuringiensis B439 [28], and B. cereus Bc7 [28], respectively, has been found to be most effective during the late log phase or early stationary phase. Ozdemir et al. [10] also discovered that bacteriocin-like inhibitory substances (BLIS) produced by G. toebii were maximally produced at the late logarithmic phase. Based on the promising results obtained in shake flasks, repeated batch fermentations were conducted using free and immobilized cells of G. subterraneus Tm6Sp1 to optimize the culture conditions, as presented in Table 2.
Table 2: Antibacterial activity in every batch against E. coli.
| Cells system preparation | Antibacterial activity in every batch against Escherichia coli | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Free cells | ++ | + | - | - |
| Immobilized cells | - | + | + | ++ |
The maximum antibacterial activity observed with free cells was 16.43 mm and 10.05 mm, respectively, that were detected in the first batch of filtrate against E. coli and S. enteritica, respectively. This finding is related to the previous study [29] on bacteriocin production. During batch fermentation, substances accumulate during the growth phase, when bacteria experience rapid growth and reach their peak. However, it was observed that the antibacterial activity sharply decreased to 8.63 mm against E. coli after the first batch of free cell culture. Furthermore, no antibacterial activity was detected in the third to fourth batches, as shown in Table 2. The statistical analysis revealed significant differences (P < 0.05) in the diameter of the zone of inhibition within every batch of fermentation [Table 3]. Additionally, the different batches of fermentation in each cell system preparation resulted in significant variations in the antibacterial activity, as evidenced by the formation of clear zones against the selected bacterial pathogens.
Table 3: Kruskal-Wallis test assessing the impact of batch fermentation on the antimicrobial activity exhibited.
| Variable | Cell system preparation | Batch 1 | Batch 2 | Batch 3 | Batch 4 | Kruskal-Wallis H | df | P-value |
|---|---|---|---|---|---|---|---|---|
| Diameter | Free cells | 62.07 | 51.93 | 28.00 | 28.00 | 43.789 | 3 | 0.000 |
| Immobilized cells | 22.50 | 34.31 | 46.79 | 66.40 | 43.564 | 3 | 0.000 |
The P-value represents the significance level of the Kruskal-Wallis test, where P≤0.05 indicate statistical significance differences
It has been similarly reported by Awais et al. [30] that during free cell synthesis of peptide antibiotics, maximum activity (14 mm) was seen at the first cycle of 24–48 h of incubation. Furthermore, the activity abruptly declined at 72 h due to the Tm6Sp1 cells entering the death phase. During batch fermentation experiments, Callewaert et al. [29] observed a significant decrease in amylovorin L471 titer after reaching peak activity during the active growth phase. This decrease in activity was accompanied by a decrease in the growth of Lactobacillus amylovorus DCE 471 in colony-forming units (CFU), indicating cell death. Moreover, it could be argued that the conditions for maximal bacteriocin production increase linearly with the cell density of the producer strain as a primary metabolite [12]. These results also showed that the free cell system was not suitable for repeated cycle culture.
In contrast, antibacterial peptides synthesis with an immobilized cell system were produced in the second batch (7.05 mm) and kept increasing during the third and fourth fermentation cycles. The highest antibacterial activity was obtained in the fourth batch, with strong inhibition against S. enterica and E. coli at 19.67 mm and 18.04 mm, respectively, as seen in Figures 5 and 6. The result showed that immobilized cell system preparation was advantageous for long-term bacteriocin production, and the encapsulated cells could be reused more than two times after filtration and resuspension in a new medium. Similarly, Hasan et al. [31] stated that the production of an antibacterial substance by immobilized B. pumilus SAF1 is higher than that of free cells. The results on the reusability of the entrapped cells by repeated batch fermentations showed that increased production was observed up to four cycles. This may be due to the proper adaptation in this microenvironment and the proper growth of cells in the beads. In the study by Alkhalili et al. [2], immobilized cells could be recovered and recycled for sequential batch fermentation with increasing antibacterial activity up to the fourteenth batch. Compared to free cells, entrapment inside the gel beads provides a protective environment for the cells that are more active at producing certain metabolites than free cells [32]. However, free and immobilized cells could produce AMPs and had strong inhibition against several bacteria.
 | Figure 6: Antibacterial activity by free cells of Tm6Sp1 for four batches. (Very strong: >20 mm, strong: 10–20 mm, moderate: 6–10 mm, weak or not detected: <6 mm). [Click here to view] |
The antibacterial compound produced by G. subterraneus Tm6Sp1 worked antagonistically against six test pathogens, with strong inhibition against S. enterica and E. coli and moderate inhibition against S. aureus, S. mutans, L. acidophilus, and K. pneumoniae. Furthermore, there was strong inhibition reported against Gram-negative bacteria. This spectrum is similar to that of the other bacteriocins, such as Geobacilin I from G. thermodenitrificans OH2-1 and Geobacillus sp. M10EXG, which could inhibit S. aureus [14], while Geobacillin I from Geobacillus ZGt-1 inhibits S. aureus and S. enterica [16]. Moreover, thermoleovorin-S2 and thermoleovorin-N9 from G. thermoleovorans inhibit the pathogenic bacteria S. enterica [11]. Antibacterial activity also occurs against K. pneumoniae by bacteriocins produced by Geobacillus spp. [16]. BLIS from G. toebii HBB-247 can inhibit the growth of several strains, including G. stearothermophilus, Listeria sp., Enterococcus faecalis, Enterococcus avium AS-3, Anoxybacillus sp. HBB-134, Geobacillus sp. HBB-269, Clostridium pasteurianum, and Cellulomonas fimi [10]. Therefore, the spectrum of bacteriocins produced by Geobacillus had a broader antibacterial activity to inhibit pathogenic bacteria, a suitable prospect for application in medicine.
5. CONCLUSION
The antibacterial substance of G. subterraneus Tm6Sp1 can be produced using free and immobilized cells in their optimum condition. Strong inhibition against pathogenic bacteria was obtained at 10.88 mm to 19.75 mm by immobilized cells in the fourth batch. The production of this substance was initiated in the second batch and increased with the incubation period cycle (repeated batch) up to the fourth batch without a decrease. In contrast, the antibacterial activity detected in the free cells system between 10.05 mm and 16.43 mm was only detected in the first batch, during the late logarithmic phase of fermentation. The result concluded that the free cell system was unsuitable for reuse in repeated batch fermentation and that the antibacterial spectrum effectively inhibited gram-negative bacteria, E. coli, and S. enteritica.
6. ACKNOWLEDGMENTS
The authors are grateful to Professor Dr. Ratu Safitri, drg. Emma Rachmawati, and Prof. Dr. Ani Melani Maskoen, the research supervisors, for the patient guidance, enthusiastic encouragement, and useful critiques provided for this study. The authors are also grateful to Akeyla Tabina and Husnul Azzahra for sharing their knowledge and cooperation. Furthermore, the authors are grateful to Ms. Shinta Asarina, the laboratory technician of the Microbiology and Parasitology Laboratory of Padjadjaran University General Education Hospital, for the assistance provided in offering the resources to run the program, to the proofreaders and language providers in DPRM of Padjadjaran University, as well as to family and friends for their support and encouragement.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All data relating to the manuscript has been provided in script.
12. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Coker JA. Extremophiles and biotechnology:Current uses and prospects. F1000Res 2016;5:396. [CrossRef]
2. Alkhalili RN, Bernfur K, Dishisha T, Mamo G, Schelin J, Canbäck B, et al. Antimicrobial protein candidates from the thermophilic Geobacillus sp. Strain ZGt-1:Production, proteomics, and bioinformatics analysis. Int J Mol Sci 2016;17:1363. [CrossRef]
3. Novik G, Savich V, Meerovskaya O. Geobacillus bacteria:Potential commercial applications in industry, bioremediation, and bioenergy production. In:Growing and Handling of Bacterial Cultures. London, UK:IntechOpen;2019. [CrossRef]
4. Nadeem MS, Al-Ghamdi MA, Khan JA. Studies on the recombinant production in E. coli and characterization of pharmaceutically important thermostable L-asparaginase from Geobacillus thermodenitrificans. Pak J Zool 2019;51:1235-41. [CrossRef]
5. Zebrowska J, Witkowska M, Struck A, Laszuk PE, Raczuk E, Ponikowska M, et al. Antimicrobial potential of the genera Geobacillus and Parageobacillus, as well as endolysins biosynthesized by their bacteriophages. Antibiotics (Basel) 2022;11:242. [CrossRef]
6. Manoharan M, Balasubramanian TS. An extensive review on production, purification, and bioactive application of different classes of bacteriocin. J Trop Biodiver Biotechnol 2022;7:72735. [CrossRef]
7. Chikindas ML, Weeks R, Drider D, Chistyakov VA, Dicks LM. Functions and emerging applications of bacteriocins. Curr Opin Biotechnol 2018;49:23-8. [CrossRef]
8. Rima M, Rima M, Fajloun Z, Sabatier JM, Bechinger B, Naas T. Antimicrobial peptides:A potent alternative to antibiotics. Antibiotics (Basel) 2021;10:1095. [CrossRef]
9. Pircalabioru GG, Popa LI, Marutescu L, Gheorghe I, Popa M, Barbu IC, et al. Bacteriocins in the era of antibiotic resistance:Rising to the challenge. Pharmaceutics 2021;13:196. [CrossRef]
10. Özdemir GB, Biyik HH. Isolation and characterization of a bacteriocin-like substance produced by Geobacillus toebii strain HBB-247. Indian J Microbiol 2012;52:104-8. [CrossRef]
11. Novotny JF, Perry JJ. Characterization of bacteriocins from two strains of Bacillus thermoleovorans, a thermophilic hydrocarbon-utilizing species. Appl Environ Microbiol 1992;98:2393-6. [CrossRef]
12. Pokusaeva K, Kuisiene N, Jasinskyte D, Rutiene K, Saleikiene J, Chitavichius D. Novel bacteriocins produced by Geobacillus stearothermophilus. Cent Eur J Biol 2009;4:196-203. [CrossRef]
13. Alkhalili RN, Canbäck B. Identification of putative novel class-I lanthipeptides in firmicutes:A combinatorial in silico analysis approach performed on genome sequenced bacteria and a close inspection of Z-geobacillin lanthipeptide biosynthesis gene cluster of the thermophilic Geobacillus sp. strain ZGt-1. Int J Mol Sci 2018;19:2650. [CrossRef]
14. Garg N, Tang W, Goto Y, Nair SK, van der Donk WA. Lantibiotics from Geobacillus thermodenitrificans. Proc Natl Acad Sci U S A 2012;109:5241-6. [CrossRef]
15. Pranckut?R, Kaunietis A, Kananavi?i?t?R, Lebedeva J, Kuisien?N, Šaleikien?J, et al. Differences of antibacterial activity spectra and properties of bacteriocins, produced by Geobacillus sp. Bacteria isolated from different environments. J Microbiol Biotechnol Food Sci 2015;5:155-61. [CrossRef]
16. Alkhalili R. Antimicrobial Potential of Geobacillus sp. ZGt-1 Isolated from Zara Hot Spring in Jordan. Sweden:Lund University;2019.
17. Garsa AK, Kumariya R, Sood SK, Kumar A, Kapila S. Bacteriocin production and different strategies for their recovery and purification. Probiotics Antimicrob Proteins 2014;6:47-58. [CrossRef]
18. Zou J, Jiang H, Cheng H, Fang J, Huang G. Strategies for screening, purification and characterization of bacteriocins. Int J Biol Macromol 2018;117:781-9. [CrossRef]
19. Okafor N, Okeke BC. Modern Industrial Microbiology and Biotechnology. 2nd ed. Alabama:CRC Press;2018.
20. Naghmouchi K, Fliss I, Drider D, Lacroix C. Pediocin PA-1 production during repeated-cycle batch culture of immobilized Pediococcus acidilactici UL5 cells. J Biosci Bioeng 2008;105:513-7. [CrossRef]
21. Rachmawati E, Asarina S, Kennardi GB, Safitri R, Subroto T, Maskoen AM. Antimicrobial peptide coding gene of thermophilic bacteria isolated from crater hot spring in mountains around West Java. J Appl Biol Biotechnol 2023;11:220-5. [CrossRef]
22. Zeigler DR. The Genus Geobacillus:Introduction and Strain Catalog. 7th ed., Vol. 3. Columbus, Ohio: Bacillus Genetic Stock Center;2001.
23. Dalynn Biologicals. Mcfarland Standard for In Vitro Use. Catalogue No. TM50-TM60. Canada:Dalynn Biologicals Inc.;2014.
24. Kaunietis A, Pranckute R, Lastauskiene E, Citavicius DJ. Medium optimization for bacteriocin production and bacterial cell growth of Geobacillus sp. 15 strain. J Antimicrob Agents 2017;3:133.
25. Davis WW, Stout TR. Disc plate method of microbiological antibiotic assay. I. Factors influencing variability and error. Appl Microbiol 1971;22:659-65. [CrossRef]
26. Marchant R, Banat IM, Rahman TJ, Berzano M. The frequency and characteristics of highly thermophilic bacteria in cool soil environments. Environ Microbiol 2002;4:595-602. [CrossRef]
27. Gautam N, Sharma N. Bacteriocin:Safest approach to preserve food products. Indian J Microbiol 2009;49:204-11. [CrossRef]
28. Ahern M, Verschueren S, van Sinderen D. Isolation and characterisation of a novel bacteriocin produced by Bacillus thuringiensis strain B439. FEMS Microbiol Lett 2003;220:127-31. [CrossRef]
29. Callewaert R, De Vuyst L. Bacteriocin production with Lactobacillus amylovorus DCE 471 is improved and stabilized by fed-batch fermentation. Appl Environ Microbiol 2000;66:606-13. [CrossRef]
30. Awais M, Pervez A, Yaqub A, Shah MM. Production of antimicrobial metabolites by Bacillus subtilis immobilized in polyacrylamide gel. Pak J Zool 2010;42:267-75.
31. Hasan F, Khan S, Shah AA, Hameed A. Production of antibacterial compounds by free and immobilized Bacillus pumilus SAF1. Pak J Bot 2009;41:1499-510.
32. Zur J, Wojcieszy?ska D, Guzik U. Metabolic responses of bacterial cells to immobilization. Molecules 2016;21:958. [CrossRef]