
1. INTRODUCTION
Production of copolymers of 3-hydroxybutyrate and 3-hydroxyvalerate [P(3HB-co-3HV)], one of the most common representatives of polyhydroxyalkanoates (PHAs) by a wide diversity of microorganisms have attracted significant attention for its improved material properties. These microbial polyester exhibits low crystallinity, increased impact strength and flexibility in addition to its biodegradability and biocompatibility [1]. However, despite its huge potential for eco-benign applications, it could not gain substantial consideration, so far, due to its low yield and expensive production cost. To circumvent these limitations, efforts are being made to find newer high-yielding microbial strains, relatively cheaper substrates for production and development of low-cost as well as easier down-stream processing strategies. Since, the carbon feedstock used in fermentation accounts for more than 50% of production cost [2], development of an optimized up-stream bioprocessing utilizing inexpensive and renewable carbon sources has become an important issue. Exploitation of agro-industrial residues and industrial wastes are beneficial options from ecological as well as economic perspectives [3].
Sugarcane molasses, the nutrient and mineral rich by-product of the sugar industry is inexpensive, available in abundance and has been exploited for the production of wide variety of industrial products including the biopolyester PHAs [4,5]. Beside sucrose, the molasses also contain other sugars, vitamins, and variety of minerals which promote microbial growth. Utilization of this sugar refinery by-product as a substrate for PHA production was initially reported by Beaulieu et al. [4] with the strain Alcaligenes eutrophus. Following this, a variety of microbial species such as Bacillus sp. [6], Bacillus megaterium [5,7], Bacillus cereus [8], Pseudomonas aeruginosa [9] and recombinant Escherichia coli [10] have been reported to produce homopolymers of 3-hydroxybutyrate [P(3HB)] from molasses. A mixed microbial culture also produced PHA containing 3-hydroxybutyrate (3HB), 3-hydroxy-2-methylbutyrate (3H2MB), 3-hydroxyvalerate (3HV), 3-hydroxy-2-methylvalerate (3H2MV) and 3-hydroxyhexanoate (3HHx) when sugarcane molasses was supplemented with synthetic feeds containing volatile fatty acids (VFAs) in the fermentation media [11]. Recently, Zafar et al. [12] has documented P(3HB-co-3HV) accumulation by Azohydromonas lata in propionic acid supplemented molasses medium. These reports focussed primarily on the process optimization for PHA production using sugarcane molasses with very little emphasis on the material properties of the accumulated biopolyesters.
Bacillus cereus RCL 02, an endophytic bacterium of oleaginous plant Ricinus communis L. has already been reported to accumulate P(3HB) in mineral salts medium containing glucose as the sole source of carbon [13]. The present study demonstrates the bioprocessing as well as partial characterization of P(3HB-co-3HV) copolyesters produced by the endophytic isolate RCL 02 following bioconversion of sugarcane molasses as the sole source of carbon in the growth medium.
2. MATERIALS AND METHODS
2.1. Bacterial Strain and Maintenance
Bacillus cereus RCL 02 (MCC 3436; GenBank accession no. KX458035), a leaf endophyte of oil-yielding plant R. communis L. was isolated in this laboratory and used throughout this study. The culture was grown on slopes of tryptic soy agar by regular sub-culturing.
2.2. Growth and Polymer Production
Microbial growth and polymer production by the endophytic isolate RCL 02 were evaluated in modified mineral salts (MS) medium [14] under batch cultivation. The MS medium contained (g/L) Na2HPO4.7H2O, 6.7; KH2PO4, 2.0; (NH4)2SO4, 1.0 and MgSO4.7H2O 1.0 (pH-7.2). The commercially available sugarcane molasses was separately sterilized and added to the medium as the sole source of carbon. The medium (25 per 100 mL Erlenmeyer flask) was inoculated with a freshly prepared inoculum of B. cereus RCL 02 and incubated at 32 °C on a rotary shaker at 120 rpm.
2.3. Measurement of Growth
Growth was determined by measuring the dry weight of the biomass. Cells were harvested by centrifugation (10,000 rpm × 10 min) after a definite period of incubation, washed thoroughly with deionized water as well as acetone and the biomass was determined after drying to a constant weight at 80 °C.
2.4. Extraction and Purification of the Polymer
The polymer accumulated in the dried cell mass was extracted thrice in warm chloroform (45-50 °C), filtered through glass wool, concentrated and precipitated with double volumes of pre-chilled diethyl ether [14]. The precipitated polymer was recovered by centrifugation (12,000 rpm × 12 min) and dried to obtain the powdered polymer.
2.5. Assay of the Polymer
The extracted polymer was converted to crotonic acid following treatment with concentrated H2SO4 in a boiling water bath for 10 min [15], cooled to room temperature and the absorbance was recorded at 235 nm in a UV-VIS spectrophotometer (Jenway, Model 6505) using authentic P(3HB) from Sigma, USA as standard. The total PHA content of the dried cell mass was quantified gravimetrically.
2.6. Preparation of the Polymer Films
Films of PHA were prepared by casting 2% (w/v) chloroform solution of the polymer onto a glass Petriplate, following slow evaporation of the solvent under regular humidity at 50 °C. The remaining solvent was evaporated for two days.
2.7. Characterization of the Polymer
2.7.1. 1H NMR spectroscopic analysis
The monomer composition of the purified polymer was elucidated from proton nuclear magnetic resonance (1H NMR) spectroscopic analysis. The polymer dissolved in deuterated chloroform (CDCl3) were analyzed in a Bruker AV300 Supercon NMR (at 300 MHz) spectrophotometer. A multinucleate probe head at 30-degree flip angle was used. The chemical shift-scale was in parts per million and tetramethylsilane (Me4Si) was used as the internal standard.
2.7.2. Fourier-transform infrared spectroscopic analysis
For FTIR spectroscopy, the pellet was prepared by mixing the purified polymer and KBr and scanned in Bruker FTIR spectrophotometer in the range of 4000 to 400 cm-1.
2.7.3. X-ray diffraction analysis
The wide angle X-ray diffractograms of the purified polymer sheets were recorded in X’pert PRO PANalytical X-ray diffractometer. The nickel filtered Cuα radiation (λ = 0.154 mm, 40 kV, 30 mA) was used at room temperature. The scan speed was 2θ = 5°/min.
2.7.4. Thermogravimetric and differential scanning calorimetric analysis
Thermal characterization was done with TA STDQ 600 thermogravimetric analyzer and Perkin-Elmer Diamond DSC thermal analyzer operating with nitrogen flow of 100 cc/min and a heating rate of 10 °C/min. While the PHA sample (10 mg) was heated from room temperature to 500 °C in an alumina crucible under dinitrogen atmosphere in TGA analysis, the scanning temperature in DSC ranged from -50 to 200 °C.
3. RESULTS AND DISCUSSION
3.1. Growth Kinetics and Accumulation of PHA
The endophytic bacterium B. cereus RCL 02 accumulated PHA during growth in molasses medium and maximum intracellular polyester accumulation [85.4% of cell dry weight (CDW)] occurred after 48 h of growth under shake-flask condition. Compositional analysis of the PHAs isolated from B. cereus RCL 02 following gravimetric as well as proton nuclear magnetic resonance (1H NMR) spectral analysis revealed that the addition of sugarcane molasses (2%, w/v) in the growth medium resulted in production of copolymers of 3HB and 3HV [P(3HB-co-3HV)]. It was also evident that during the early stages of growth and polyester accumulation, there was no incorporation of 3HV monomers in the polymer, but the 3HV monomer fraction in PHA increased to 5.1 mol% at the highest polyester accumulation phase. Biosynthesis of P(3HB-co-3HV) was associated with a decline in pH of the growth medium (pH 5.2 after 96 h) which may possibly be due to the production of several metabolic intermediates during growth [16]. In addition, cells of B. cereus RCL 02 started sporulating in late stationary phase and about 78% of cells showed matured endospores at the end of 96 h of incubation (Figure 1). Such delayed sporogenesis, therefore, could not interfere in the biosynthesis and accumulation of the intracellular polymer during the exponential to early stationary phases of growth. Concomitant with the increase in the frequency of sporulation, there was an initiation of decline in the PHA content of the cells from the late stationary phase of growth. Such decline in intracellular PHA may be due to the possible utilization of the accumulated polyesters as carbon and energy sources for the process of sporogenesis.
The present study clearly indicated that the efficiency of utilizing sugarcane molasses for PHA production by the endophytic bacterium B. cereus RCL 02 was much better as compared to the purified sugars viz. glucose, sucrose, maltose etc. [13]. It may also be presumed that molasses served not only as a potential source of carbon but also provided vitamins and other essential growth factors for effective growth and sustainable production of the biopolyester by the isolate RCL 02. Survey of literature pertaining to the PHA production utilizing sugarcane molasses clearly indicated that most of the bacterial strains were explored mainly for synthesis and accumulation of the homopolyester, P(3HB) [4-10,17]. While, supplementation of different volatile fatty acids [11] and propionic acid [12] along with sugarcane molasses have led to the production of PHA copolymers, here we report the accumulation of P(3HB-co-3HV) utilizing molasses as the sole source of carbon by the endophytic bacterium B. cereus RCL 02. Production of intermediate acids during growth and polymer accumulation by RCL 02 in molasses medium might contribute to the incorporation of 3HV monomers in the polymer chain.
 | Figure 1: Time course of growth, PHA production, sporulation, changes in pH of the medium and 3HV monomer incorporation in the copolymer produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
3.2. Effect of Molasses Concentration
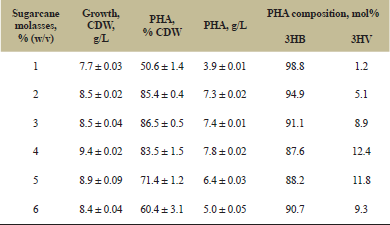
Influence of different concentration of sugarcane molasses on growth, production of the copolymer and 3HV monomer (mol%) incorporation was elucidated. Addition of molasses at 4% (w/v) level in the growth medium showed maximum production of P(3HB-co-3HV) (7.8 g/L) with the incorporation of 12.4 mol% 3HV monomers (Table 1). However, the PHA content of the cell mass (83.5%, CDW) was lower than those grown in 3% (w/v) molasses. Further increase in molasses concentration showed a negative effect on biomass formation, polymer accumulation as well as 3HV monomer incorporation in the copolymer and corroborated well with the findings of Zafar et al. [12].
3.3. Characterization of the Polymer
The copolymer so produced with 12.4 mol% 3HV was isolated and purified from the cell mass and partially characterized.
3.3.1. 1H NMR spectroscopic analysis
The 1H NMR spectrum (at 300 MHz) (Figure 2) of the copolyester displayed chemical shifts (δ) at 1.26 and 0.89 ppm indicating the presence of methyl (CH3) group of 3HB (87.6 mol%) and 3HV (12.4 mol%) monomer units, respectively. Moreover, the characteristic resonance absorption at 2.4-2.6 and 5.2 ppm were assignable to the methylene group (CH2) and methyne group (CH), respectively. The characteristic signals were consistent with the previous findings of Masood et al. [18].
 | [Click here to view] |
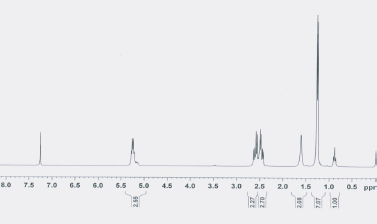 | Figure 2: 1H NMR spectra of purified copolyester P(3HB-co-3HV) produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
3.3.2. Fourier-transform infrared spectroscopic analysis
The FTIR spectra (Figure 3) of the copolyester showed absorption bands at 1727 cm-1 and 1278 cm-1 corresponding to ester C=O and C-O groups, respectively. The other characteristic bands at 3439 cm-1, 2933-2979 cm-1 and 1225-1382 cm-1 represented the -OH bending, C-H stretching and -CH group, respectively. These absorption peaks showed close proximity with the peaks of P(3HB-co-10 mol% 3HV) from non-endophytic B. cereus SPV [19] and provided supportive evidence in favor of the accumulation of similar copolyester by the endophyte B. cereus RCL 02.
3.3.3. X-ray diffraction analysis
The wide angle X-ray diffractograms of the purified copolymer were represented as peak positions at 2θ and X-ray counts (intensity) in a xy plot (Figure 4). The sharp and intense peak at 13° was attributed to high crystallinity. The XRD pattern of the copolymer was consistent when compared with previous crystallographic studies of P(3HB-co-3HV) [20].
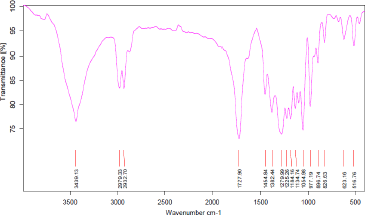 | Figure 3: FTIR spectra of purified copolyester P(3HB-co-3HV) produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
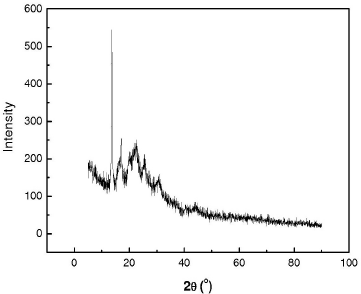 | Figure 4: X-ray diffraction pattern of purified copolyester P(3HB-co-3HV) produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
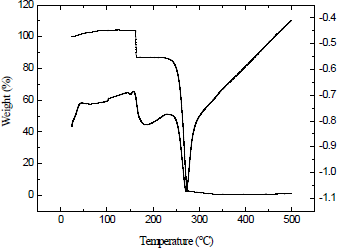
3.3.4. Thermogravimetric analysis
Thermogravimetric analysis (TGA) of P(3HB-co-3HV) from B. cereus RCL 02 showed that the major weight loss occurred between 230 to 300 °C as a single exotherm (Figure 5). The weight loss at the temperature <150 °C observed in the thermogram was due to loss of components like water, low molecular weight solvents or gas desorption [8]. The degradation temperature of the copolymer (in terms of 50% weight loss) was recorded at 263.14 °C and thus indicated its thermostability. The degradation was probably associated with ester cleavage of the copolyester chain by the β-elimination reaction.
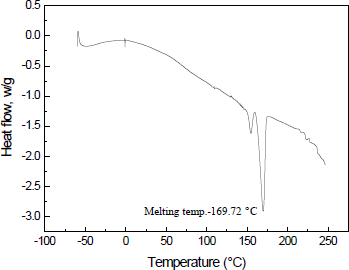
3.3.5. Differential scanning calorimetry
Determination of melting behavior of the polymer is crucial for its processing. Differential scanning calorimetry (DSC) study of the isolated and purified P(3HB-co-3HV) showed a melting temperature (Tm) of 169.72 °C and enthalpy of fusion (Δm) of 63.98 J/g (Figure 6). The values of Tm, as well as Δm, were consistent with findings of You et al. [21]. Incorporation of 3HV monomers into the 3HB polymer chain usually decreases the Tm as well as Δm of the copolymer matrix and thus reduces its crystallinity [22].
4. CONCLUSION
Novel bacterial strains with high production efficiency and cost-effective substrates are always essential for potential industrial usage to enhance process economy. The present study revealed that sugarcane molasses, an industrial by-product of sugar industries were efficiently utilized by the endophytic bacterial isolate B. cereus RCL 02 for the production of copolyesters of 3HB and 3HV. Supplementation of 4% (w/v) molasses as the sole carbon source in MS medium incorporated 12.4 mol% 3HV in the P(3HB-co-3HV) copolyester chain indicating its potential industrial application.
 | Figure 5: Thermogravimetric analysis of purified copolyester P(3HB-co-3HV) produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
 | Figure 6: DSC curve showing melting temperature (Tm) of purified copolyester P(3HB-co-3HV) produced by the endophytic isolate B. cereus RCL 02. [Click here to view] |
5. ACKNOWLEDGMENTS
Financial support from Department of Science and Technology, New Delhi, India in the form of INSPIRE Fellowship (Sanction No. DST/INSPIRE Fellowship/REL5/3013/C1) to Rituparna Das is duly acknowledged.
6. REFERENCES
1. Anjum A, Zuber M, Zia KM, Noreen A, Anjum MN, Tabasum S. Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: a review of recent advancements. International Journal of Biological Macromolecules 2016; 89:161-174.CrossRef
2. Sindhu R, Silviya N, Binod P, Pandey A. Pentose-rich hydrolysate from acid pretreated rice straw as a carbon source for the production of poly-3-hydroxybutyrate. Biochemical Engineering Journal 2013; 78:67-72.CrossRef
3. Koller M, Atlić A, Dias M, Reiterer A, Braunegg G. Microbial PHA production from waste raw materials. In: Microbiology monographs, 2010, p. 85-119.CrossRef
4. Beaulieu M, Beaulieu Y, Melinard J, Pandian S, Goulet J. Influence of ammonium salts and cane molasses on growth of Alcaligenes eutrophus and production of polyhydroxybutyrate. Applied and Environmental Microbiology 1995; 61(1):165-169.
5. Gouda MK, Swellam AE, Omar SH. Production of PHB by a Bacillus megaterium strain using sugarcane molasses and corn steep liquor as sole carbon and nitrogen sources. Microbiological Research 2001; 156(3):201-207.CrossRef
6. Wu Q, Huang H, Hu G, Chen J, Ho KP, Chen GQ. Production of poly-3-hydroxybutyrate by Bacillus sp. JMa5 cultivated in molasses media. Antonie van Leeuwenhoek 2001; 80(2):111-118.CrossRef
7. Chaijamrus S, Udpuay N. Production and characterization of polyhydroxybutyrate from molasses and corn steep liquor produced by Bacillus megaterium ATCC 6748. Agricultural Engineering International: CIGR Ejournal. Manuscript FP 07 030, May, 2008, Vol. X: 1-12.
8. Sharma P, Bajaj BK. Production and characterization of poly-3-hydroxybutyrate from Bacillus cereus PS 10. International Journal of Biological Macromolecules 2015; 81:241-248.CrossRef
9. Tripathi AD, Yadav A, Jha A, Srivastava SK. Utilizing of sugar refinery waste (cane molasses) for production of bio-plastic under submerged fermentation process. Journal of Polymers and the Environment 2012; 20(2):446-453.CrossRef
10. Liu F, Li W, Ridgway D, Gu T, Shen Z. Production of poly-β-hydroxybutyrate on molasses by recombinant Escherichia coli. Biotechnology Letters 1998; 20(4):345-348.CrossRef
11. Bengtsson S, Pisco AR, Johansson P, Lemos PC, Reis MAM. Molecular weight and thermal properties of polyhydroxyalkanoates produced from fermented sugar molasses by open mixed cultures. Journal of Biotechnology 2010; 147(3-4):172-179.CrossRef
12. Zafar M, Kumar S, Dhiman AK. Artificial intelligence based modeling and optimization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) production process by using Azohydromonas lata MTCC 2311 from cane molasses supplemented with volatile fatty acids: a genetic algorithm paradigm. Bioresource Technology 2012; 104:631-641.CrossRef
13. Das R, Pal A, Paul AK. Production of biopolyester poly(3-hydroxybutyrate) by Bacillus cereus RCL 02, a leaf endophyte of Ricinus communis L. Journal of Microbiology and Biotechnology Research 2017; 7(4):32-41.CrossRef
14. Ramsay BA, Lomaliza K, Chavarie C, Dube B, Bataille P, Ramsay JA. Production of poly-(β-hydroxybutyric-co-β-valeric) acids. Applied and Environmental Microbiology 1990; 56(7):2093-2098.
15. Law JH, Slepeckey RA. Assay of polyhydroxybutyric acid. Journal of Bacteriology 1961; 82(1):32-36.
16. Benoit TG, Wilson GR, Baugh CL. Fermentation during growth and sporulation of Bacillus thuringiensis HD-1. Letters in Applied Microbiology 1990; 10(1):15-18.CrossRef
17. Kumar BS, Prabhakaran G. Production of PHB (bioplastic) using bio-effluent as substrate by Alcaligens eutrophus. Indian Journal of Biotechnology 2006; 5:76-79.
18. Masood F, Hasan F, Ahmed S, Chen P, Hameed A. Biosynthesis and characterization of poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) from Bacillus cereus S10. Journal of Polymers and the Environment 2012; 20(3):865-871. CrossRef
19. Valappil SP, Rai R, Bucke C, Roy I. Polyhydroxyalkanoate biosynthesis in Bacillus cereus SPV under varied limiting conditions and an insight into the biosynthetic genes involved. Journal of Applied Microbiology 2008; 104(6):1624-1635. CrossRef
20. Sato H, Ando Y, Mitomo H, Ozaki Y. Infrared spectroscopy and X-ray diffraction studies of thermal behavior and lamella structures of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (P(HB-co-HV)) with PHB-type crystal structure and PHV-type crystal structure. Macromolecules 2011; 44(8):2829-2837.CrossRef
21. You JW, Chiu HJ, Shu WJ, Don TM. Influence of hydroxyvalerate content on the crystallization kinetics of poly(hydroxybutyrate-co-hydroxyvalerate). Journal of Polymer Research 2003; 10(1):47-54. CrossRef
22. Chen LJ, Wang M. Production and evaluation of biodegradable composites based on PHB-PHV copolymer. Biomaterials 2002; 23(13):2631-2639.CrossRef