
1. INTRODUCTION
The Nampu plant (Homalomena rostrata Griff) is a member of the Araceae family, renowned for its attractive leaf shape (attractive size and color) and ability to thrive in various environmental conditions. Its morphological description includes upright stems and broad and oval-shaped leaves. This plant is generally found in tropical rainforests, especially in moist and shaded areas [1]. Its natural habitat contributes to the diversity of flavonoid compounds produced.
The traditional uses of the Nampu plant are highly varied. In addition to being an ornamental plant, parts of this plant are utilized in traditional medicine for various health issues, including digestive disorders and inflammation. The economic value of this plant is also increasing, coinciding with the growing public interest in ornamental plants and herbal medicines [2]. Further research into the secondary metabolite production of this species could open new avenues for the development of herbal products.
Research can exhibit that the Nampu plant contains various bioactive compounds that may contribute to its medicinal properties. For instance, studies have indicated that flavonoids and other phenolic compounds present in the plant can exhibit antioxidant and anti-inflammatory activities [3]. These properties are particularly relevant in the context of traditional medicine, where such compounds are often sought for their therapeutic benefits. The increasing interest in natural remedies has led to a resurgence in the study of plants like Nampu, which could serve as a source of new herbal products [4].
Moreover, the economic potential of the Nampu plant is underscored by its dual role as both an ornamental purpose and a medicinal plant. As consumer preferences shift toward natural and organic products, the demand for plants with both esthetic and health benefits is likely to grow. This trend presents opportunities for local farmers and businesses to cultivate and market Nampu, thereby enhancing its economic viability [5]. The Nampu plant (H. rostrata Griff) is not only valued for its ornamental appeal but also for its traditional medicinal uses. Exploring its chemical composition and pharmacological activity could enhance its economic value and lead to the development of innovative herbal products that cater to the growing demand for natural remedies. The pharmacological potential of this plant can be attributed to its secondary metabolite content, which serves as an active compound in plant defence and exhibits biological effects in humans.
Tissue culture is a sophisticated technique employed for the propagation of plants under sterile laboratory conditions, allowing researchers to isolate and culture plant cells, tissues, or organs in suitable media. This method is particularly beneficial for the conservation of rare species and the development of superior varieties, as it enables the production of genetically uniform plants. This is crucial in preserving genetic diversity and ensuring the availability of valuable plant species for future generations. The influence of culture conditions on the production of secondary metabolites is important. Factors such as media composition, light intensity, temperature, and the presence of growth hormones can markedly affect both the quantity and quality of secondary metabolites produced in tissue culture systems. For instance, the choice of culture medium can provide essential nutrients and growth factors that are crucial for the optimal growth of plant cells and tissues. In addition, light conditions can influence photosynthetic activity and, consequently, the biosynthesis of secondary metabolites, which are often derived from primary metabolic pathways [6].
Previous studies have demonstrated that the addition of specific hormones to culture media can enhance the production of flavonoids and alkaloids in several plant species. For example, the incorporation of auxins and cytokinins in varying concentrations has been shown to stimulate cell division and differentiation, leading to increased yields of valuable secondary metabolites [7,8]. Furthermore, the manipulation of environmental factors, such as temperature and light duration, can also lead to significant concentration variations in metabolite profiles. Highlighting the need for a thorough understanding of optimal culture conditions to maximize secondary metabolite production [9,10]. Tissue culture techniques can yield a wider variety and higher quantities of secondary metabolites compared to conventionally grown plants. This enhanced production potential presents exciting opportunities for developing new applications in pharmaceuticals, nutraceuticals, and other industries. Secondary metabolites, such as flavonoids, alkaloids, and terpenoids, are known for their bioactive properties and have been widely studied for applications in pharmaceuticals, nutraceuticals, and cosmetics. The ability to produce these compounds in a controlled environment through tissue culture not only ensures a consistent supply but also enables the exploration of novel compounds that may have been previously difficult to obtain from traditional cultivation methods [11]. Tissue culture is a powerful tool for both plant propagation and the production of secondary metabolites. By carefully manipulating culture conditions, the yield and diversity of valuable compounds can be enhanced, opening new possibilities in medicine, agriculture, and biotechnology. The careful manipulation of culture conditions can lead to significant enhancements in the yield and diversity of valuable compounds, paving the way for innovative applications in various fields, including medicine, agriculture, and biotechnology.
Preliminary studies on the in vitro propagation of Homalomena aromatica Schott., an endangered aromatic medicinal herb native to Northeast India, have demonstrated its potential to enhance plant availability and conservation efforts. Tissue culture techniques provide a controlled environment for the propagation of this species, which is crucial given its endangered status and the increasing demand for its medicinal properties. One significant study by Raomai et al. demonstrated successful in vitro propagation of H. aromatica through rhizome axillary bud multiplication [12]. The researchers cultured rhizome bud explants on Murashige and Skoog (MS) medium supplemented with various concentrations of cytokinins to induce multiple shoot formation for micropropagation. The highest number of shoots was achieved in MS medium supplemented with 2.0 mg/L 6-benzylaminopurine (BAP) [12].
There have been no documented instances of callus induction in H. rostrata Griff plants using the growth regulators 2,4-D and kinetin. This research explores the potential for in vitro propagation and the production of valuable secondary metabolites. Tissue culture techniques enable the controlled growth of plant cells, tissues, or organs under sterile conditions, which is particularly beneficial for species like H. rostrata that are difficult to propagate through conventional methods. This approach not only facilitates the rapid production of plants but also ensures genetic uniformity, which is essential for conservation and commercial applications.
2. MATERIALS AND METHODS
2.1. Materials
MS medium (Himedia, India), Indole-3-butyric acid (IBA) (Himedia, India), 2,4-Dichlorophenoxyacetic acid (2,4-D) (Himedia, India), 1-Naphthylacetic acid (NAA) (Himedia, India), pineapple extract, kinetin (Himedia, India), BAP (Himedia, India), silica gel 60 F254 (Merck, Germany), 10% H2SO4 in methanol (Merck, Germany), and Ferric chloride (FeCl3) (Merck, Germany).
2.2. Preparation of Nampu Leaves
2.2.1. Determination of nampu plant
The determination of the plant was conducted at the Herbarium Bogoriense, Botany Division, LIPI Research Centre, Bogor, confirming that the specimen is indeed the Nampu plant (H. rostrata Griff).
2.3. Tissue Culture of Nampu Leaves
2.3.1. Sterilization of space and instrument
The culture space was cleaned by spraying the working surface with 70% alcohol. The tools to be used were sterilized by washing them thoroughly with detergent and then air-drying. Once dry, they were wrapped in paper (except for culture bottles) and placed in an autoclave at 121°C for 15 min. Planting tools such as tweezers and scalpels could be sterilized again by heating over a spirit flame after being dipped in 70% alcohol before planting (the process is repeated between each use).
2.3.2. Sterilization of distilled water and culture media
The media and distilled water used were first sterilized in an autoclave. Distilled water was sterilized in culture bottles sealed with plastic and autoclaved for 15 min at 121°C. The culture media to be used were sterilized in the autoclave at the same temperature and pressure for 15 min. The basic medium used was the MS medium.
2.3.3. Preparation of media
The preparation of the MS medium involved weighing the MS components, placing them into a measuring flask, and adding distilled water to the appropriate level.
2.3.4. Optimization of sterilization of nampu rhizome explants
The nampu rhizome was washed with running water and then soaked in a 70% alcohol solution for 1 minute to clean its outer surface. Subsequently, it was immersed in a mixture of three drops of Tween and various soaking times in 2% hypochlorite, specifically for 5 min, 7 min, and 10 min. To cleanse the internal tissues of the plant, it was then rinsed with sterile distilled water for 1 min in three portions.
2.3.5. Plant growth regulators (PGRs) to MS media
The growth hormones (PGRs) used in this study included NAA and BAP [Tables 1-4].
Table 1: Concentration of NAA: BAP used on Nampu leaves.
| NAA: BAP concentration (mg/L) | |||
|---|---|---|---|
| 0:1 | 0.5:0 | 1:0 | 1.5:0 |
| 0:5 | 0.5:1 | 1:1 | 1.5:1 |
| 0:10 | 0.5:5 | 1:5 | 1.5:5 |
| 0.5:10 | 1:10 | 1.5:10 | |
NAA: 1-Naphthylacetic acid, BAP: 6-benzylaminopurine.
Table 2: Concentration of IBA: Kinetin used on Nampu leaves.
| Concentration IBA: Kinetin (mg/L) | |
|---|---|
| 0.5:1 | 0.75:1.5 |
| 1:1 | 1.5:1.5 |
| 1:0.5 | 1.5:0.75 |
IBA: Indole-3-butyric acid.
Table 3: Concentration of 2,4-D: Pineapple extract used on Nampu leaves.
| 2,4D (mg/L): Pineapple extract (%) |
|---|
| 0.5:15 |
| 1:15 |
| 1.5:15 |
Table 4: Concentration of 2,4-D: Kinetin used on Nampu leaves 2,4-d: Kinetin (mg/L).
| 2,4 D: Kinetin (mg/L) | |||
|---|---|---|---|
| 0:1 | 1:0 | 1:2 | 2:1 |
| 0:2 | 1:1 | 2:0 | 2:2 |
The MS media solution, once thoroughly mixed, was assessed for acidity using a pH meter. If the acidity of the medium was found to be <5.4, a few drops of 1 N NaOH solution were added; conversely, if the medium exhibited an acidity >5.8, several drops of 1 N HCl were added to achieve the desired acidity.
A gelling agent was added once the pH of the media solution was adjusted to the desired level. The solution was heated until it came to a boil. The heated media solution was then transferred into culture bottles and sealed with aluminium foil. The filled bottles were autoclaved for 15 min at 121°C. The autoclaved media were stored in a cool place for a period before being utilized for planting. This storage aimed to ascertain the presence or absence of contamination within the culture media before the planting of explants.
2.3.6. Sterilization of nampu leaf explants
Nampu leaves were washed with running water, soaked in a 70% alcohol solution for 1 min to clean the outer surface of the plant, followed by immersion in a mixture of three drops of Tween and 2% hypochlorite for 5 min to clean the internal tissues of the plant, and then rinsed with sterile distilled water for 1 min in three portions.
2.3.7. Culture of nampu leaves
Explants were cut to a size of 1 × 1 cm (each growth regulator substance was repeated 3 times), with the edges of each piece wound before planting them into MS media containing various growth hormones. The bottles were then tightly sealed and placed in an incubator at a temperature of 25°C.
2.4. Preparation of Nampu Leaf Extract from Tissue Culture
The preparation of Nampu leaf extract was performed using a cold extraction method, specifically maceration with sonication using a series of solvents, including n-hexane, ethyl acetate, and methanol. A quantity of dried simplicia from the tissue culture was placed into a vial, and n-hexane solvent was added until fully submerged. This was left for 20 min. The filtrate was then filtered, and the residue was re-macerated with ethyl acetate until submerged and treated similarly to the n-hexane. The resulting filtrate was re-macerated with methanol until it was fully submerged, and then it was filtered.
2.5. Analysis of Secondary Metabolites
The concentrated extracts of Nampu leave from tissue culture and simplicia were dissolved in their respective solvents and then spotted on a GF254 silica gel plate measuring 5 × 7 cm. A developing solvent of n-hexane and ethyl acetate (7:3) was used and saturated in a chamber for elution with the specified developing solvent on the silica plate. Subsequently, spraying was performed using 10% H2SO4 in methanol, Lieberman–Burchard, FeCl3, and citroborate. The Thin Layer Chromatography (TLC) profile of the extract from the tissue culture of Nampu leaves was compared with that of the extract from Nampu leaves growing in their natural habitat.
2.6. Isolation of Secondary Metabolites Using Preparative TLC
A silica gel slurry was prepared by mixing 25 g of silica gel 60 F254 with 50 mL of distilled water (1:2). This mixture was vigorously shaken in a stoppered Erlenmeyer flask for approximately 90 s until it became homogeneous. The silica slurry was then poured and spread onto a glass plate, followed by a 24-h resting period in an oven at 106°C for 30–60 min. The saturation of the developing solvent was performed using a 7:3 ratio of n-hexane and ethyl acetate, which was placed in a chamber and allowed to saturate for 60 min. Subsequently, the n-hexane fraction extract was spotted onto the silica plate in a series, forming a band as the starting line for development (1–2 cm from the bottom edge). During the elution phase, the plate containing the sample spots was placed in the saturated developing solvent chamber and left until the elution process reached the edge of the plate (1–2 cm from the top edge). Afterwards, it was removed and allowed to dry.
2.7. Identification of Isolate Structure
The identification of the isolated structure was conducted using UV-Vis spectrophotometry to determine the wavelength and infrared (IR) spectroscopy to ascertain the main functional groups present in the sample.
2.7.1. Absorption of isolate in ultraviolet visible (UV-Vis) spectrophotometry
The sample was dissolved in a solvent that does not absorb in the UV-Vis spectrophotometry range. Identification was performed using UV-Vis spectrophotometry to determine the wavelength of the sample exposed to light, thereby identifying the functional groups present within the sample.
2.7.2. Method for determining functional groups using IR spectrophotometry
The sample was placed in a KBr pellet, mixed with KBr at a concentration of 0.1–2.0% by weight of the mixture, and then ground in a mortar and pressed to eliminate any air trapped within the mixture. The instrument employed for this analysis was Fourier Transform Infrared Spectroscopy (FTIR).
3. RESULTS
3.1. Optimization of Sterilization
The optimization of sterilization for the rhizome of the Nampu plant was conducted by varying the soaking time in sodium hypochlorite at a concentration of 2% v/v for durations of 5, 7, and 10 min. However, this optimization did not yield optimal sterilization results due to the high water content in the rhizome, which led to browning.
The sterilization of nampu leaves was performed using sodium hypochlorite at a concentration of 2% v/v for 5 min. This condition was deemed relatively ideal for sterilizing the Nampu leaves, as excessively high concentrations of hypochlorite and prolonged soaking times could damage the plant cell tissues. Sterilization of leaf explants was achieved by soaking them in 70% alcohol for 1 min.
3.2. Callus Induction
The induction of the explants was carried out by planting the Nampu leaf explants in a medium supplemented with various types of PGRs at different concentrations. The PGRs utilized in this study included NAA-BAP, 2,4-D-Kinetin, 2,4-D-pineapple extract, IBA-Kinetin, and a control medium without PGR supplementation.
The variation in PGRs that provided the best development of the explants was found to be NAA: BAP (1.5:10 mg/L), 2,4-D-Kinetin (2:2 mg/L), 2,4-D: Pineapple extract (0.5:15000 mg/L), IBA: Kinetin (0.5:1 mg/L), and the control without PGRs. The leaf explants without PGR treatment, serving as the control, on day 33 displayed curling and a yellowish-green color [Figure 1]. The development of the explants using the PGR NAA: BAP (1.5:10 mg/L) on day 98 showed that the leaves were nearly curled, and the upper surface of the leaves appeared white, with no callus formation observed [Figure 2]. In the case of the leaf explants treated with the 2,4-D-Pineapple extract (0.5 mg/L:15%) on day 68, the campus leaves had not undergone any morphological changes, although the leaf color had faded [Figure 3]. The results of tissue culture using the PGR 2,4-D-Kinetin (2:2 mg/L) on day 33 indicated that the leaf explants were curling and the callus was brown [Figure 4]. The leaf explants treated with the medium containing IBA-kinetin (0.5: 1 mg/L) on day 41 exhibited curling, accompanied by brown callus [Figure 5].
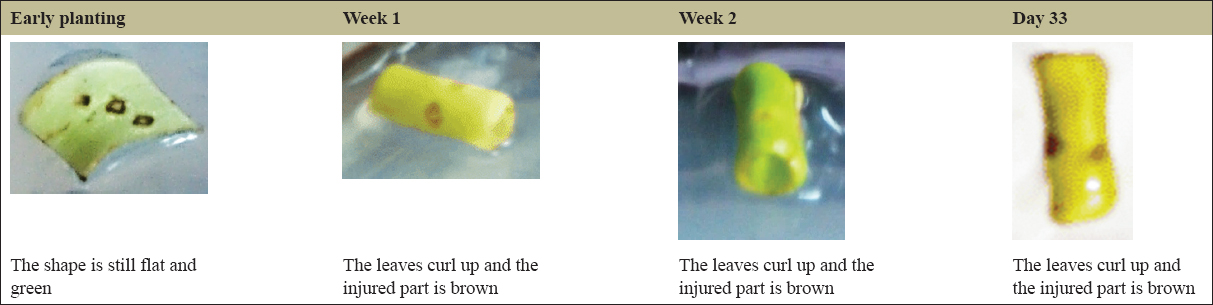 | Figure 1: The growth of Nampu leaf explants on Murashige and Skoog medium. [Click here to view] |
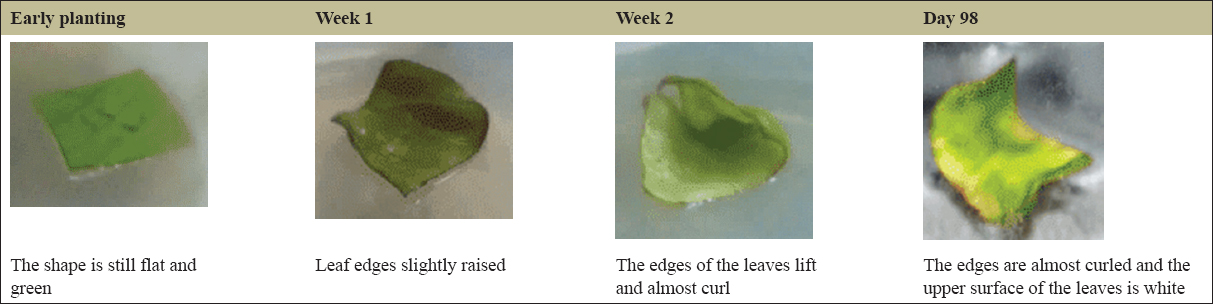 | Figure 2: The growth of Nampu leaf explants on Murashige and Skoog medium + NAA: BAP (1.5:10). [Click here to view] |
 | Figure 3: The growth of Nampu leaf explants on Murashige and Skoog medium + 2.4 D: Pineapple extract (0.5:15000 mg/L). [Click here to view] |
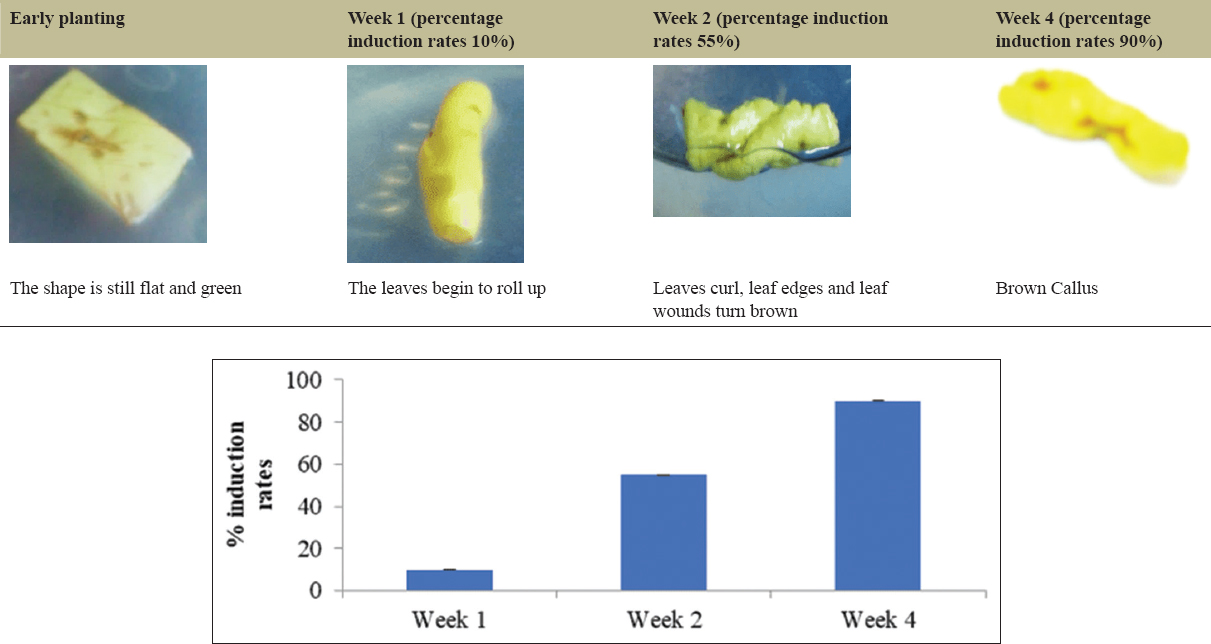 | Figure 4: The growth of nampu leaf explants on MS medium + 2,4D: Kinetin (2:2). [Click here to view] |
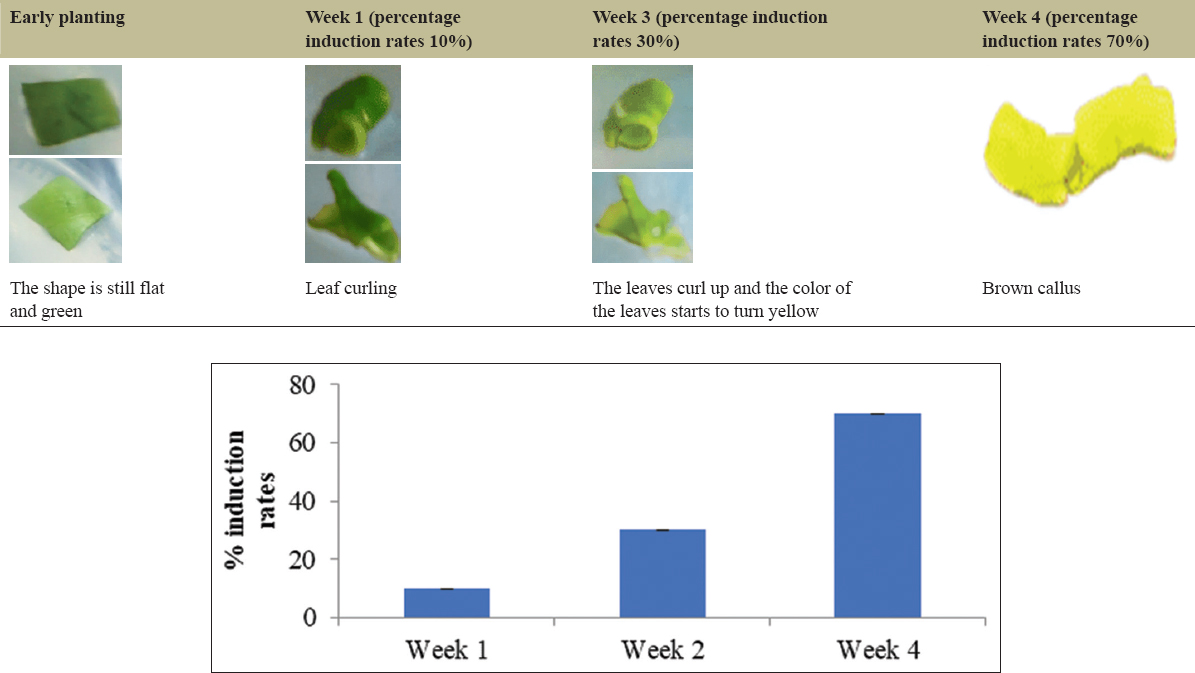 | Figure 5: The growth of nampu leaf explants on MS medium + IBA: Kinetin (0.5:1). [Click here to view] |
The optimal ratio of kinetin to 2,4-D can vary based on the species being cultured. Kinetin, a type of cytokinin, plays a vital role in cell division, while 2,4-D, an auxin, facilitates the initial formation of callus from explants. Numerous studies support the significance of these growth regulators in enhancing morphogenetic responses in tissue culture systems. Research has shown that varying concentrations and combinations of PGRs have a significant influence on callus induction. The percentage of callus growth in the medium supplemented with growth regulators is 90%, indicating a potential presence of steroid compounds within the callus.
According to Masekesa et al., reducing the ratio of 2,4-D to kinetin has been linked to a shift in developmental response from callus formation to direct organogenesis, particularly in species like sweet potato and wheat [13]. Similarly, various combinations, such as those tested by Rodiansah et al. for garlic, confirm that specific concentrations of auxins and cytokinins are essential for effective callus induction [14]. These findings are echoed by slazak et al., which highlight the role of indirect organogenesis through callus formation, emphasizing the importance of appropriate PGR ratios in achieving high regeneration rates [15].
3.3. Analysis of Secondary Metabolites from callus
The extraction method employed for the leaves of Nampu, derived from tissue culture, utilized the maceration extraction technique combined with sonication, owing to the minimal weight of the tissue-cultured leaves. In contrast, the extraction of Nampu leaves without tissue culture was conducted using a maceration method without the use of sonication. This study employed the maceration technique, a cold extraction process, which is anticipated to preserve the integrity of the compounds by avoiding heat exposure. Both types of leaves were subjected to maceration in succession. N-hexane was employed to extract non-polar compounds, followed by ethyl acetate for semi-polar compounds, and finally methanol for polar compounds.
Dried simplicia were submerged in n-hexane until fully immersed. For the tissue-cultured Nampu leaves, the immersion was maintained for 24 h, while for the Nampu leaves without tissue culture, the duration was extended to 24 h. Subsequently, the filtrate was filtered, and the residue was re-macerated with ethyl acetate until it was fully submerged, then treated in the same manner as with n-hexane. The resultant filtrate was then re-macerated with methanol until fully immersed and subsequently filtered.
The viscous extracts from both types of Nampu leaves underwent secondary metabolite analysis via TLC using the same developing solvent. The developing solvent employed for eluting both extracts was a mixture of n-hexane and ethyl acetate in a ratio of 7:3. The TLC profiles of both extracts were compared and sprayed with Lieberman–Burchard reagent, citroborate, and FeC3. The TLC profiles obtained from the tissue-cultured Nampu leaves and those without tissue culture exhibited distinct chromatographic patterns.
Visually, the tissue-cultured Nampu Callus was not detected due to its low metabolite content; however, spots were observed under UV light at 365 nm [Figure 6]. The visualization agent used was 10% H2SO4 in methanol, followed by heating on the plate. The heating process evaporated the solvent and facilitated the detection of organic compounds, as indicated by a color change after heating. In contrast, the Nampu leaves without tissue culture displayed a slight brown color change, whereas several tissue-cultured Nampu leaves were not visually detectable due to the low concentration of the extract. In addition to the universal visualization agent, analyses were conducted using specific visualization agents, including Lieberman–Burchard, Anisaldehyde, corroborate, and FeCl3 [
 | Figure 6: Thin layer chromatography results using the mobile phase n-hexane: ethyl acetate (7:3) in the control (MS medium). 1: N-hexane extract, 2: Ethyl acetate extract, 3: Methanol extract, 4: N-hexane extract of nampu leaves from tissue culture, 5: Ethyl acetate extract of nampu leaves from tissue culture, 6: Methanol extract of nampu leaves from tissue culture. (a) Visual, (b) ultraviolet (UV) 365 nm, (c) UV 254 nm, (d) H2SO4 spot appearance (Visual), (e) H2SO4 spot appearance (UV 365 nm), (f) Lieberman–Burchard spot appearance (Visual), (g) Lieberman–Burchard spot appearance (UV 365 nm), (h) Citroborate spot appearance (Visual), (i) Citroborate spot appearance (UV 365 nm), (j) Citroborate spot appearance (UV 254 nm), (k) FeCl3 spot appearance (Visual). [Click here to view] |
 | Figure 7: Thin layer chromatography using the mobile phase n-hexane: ethyl acetate (7:3) in NAA: BAP. 1: N-hexane extract, 2: Ethyl acetate extract, 3: Methanol extract, 4: N-hexane extract of nampu leaves from tissue culture, 5: Ethyl acetate extract of nampu leaves from tissue culture, 6: Methanol extract of nampu leaves from tissue culture, (a) Visual, (b) ultraviolet (UV) 365 nm, (c) UV 254 nm, (d) H2SO4 spot appearance (Visual), (e) H2SO4 spot appearance (UV 365 nm), (f) Lieberman–Burchard spot appearance (Visual), (g) Lieberman–Burchard spot appearance (UV 365 nm), (h) Citroborate spot appearance (Visual), (i) Citroborate spot appearance (UV 365 nm), (j) Citroborate spot appearance (UV 254 nm), (k) FeCl3 spot appearance (Visual). [Click here to view] |
 | Figure 8: Thin layer chromatography using the mobile phase n-hexane: ethyl acetate (7:3) on PGR 2.4D: Pineapple extract. 1: N-hexane extract, 2: Ethyl acetate extract, 3: Methanol extract, 4: N-hexane extract of nampu leaves from tissue culture, 5: Ethyl acetate extract of nampu leaves from tissue culture, 6: Methanol extract of nampu leaves from tissue culture, (a) Visual, (b) ultraviolet (UV) 365 nm, (c) UV 254 nm, (d) H2SO4 spot appearance (Visual), (e) H2SO4 spot appearance (UV 365 nm), (f) Lieberman–Burchard spot appearance (Visual), (g) Lieberman–Burchard spot appearance (UV 365 nm), (h) Citroborate spot appearance (Visual), (i) Citroborate spot appearance (UV 365 nm), (j) Citroborate spot appearance (UV 254 nm), (k) FeCl3 spot appearance (Visual). [Click here to view] |
Detection using Lieberman–Burchard resulted in a visual color change to pink, suggesting the presence of steroid compounds in both the Nampu leaves and those derived from tissue culture. The presence of polyphenolic compounds in Nampu leaves was indicated by the visualization agent FeCl3, which produced a black color on heating, with polyphenols in the Nampu leaves detected in the ethyl acetate and methanol extracts. Furthermore, citroborate was used to detect the presence of flavonoids, as evidenced by bright blue fluorescence under UV light at 365 nm [Figures 9 and 10].
 | Figure 9: Thin layer chromatography using the mobile phase n-hexane: ethyl acetate (7:3) on PGR 2,4D: Kinetin. 1: N-hexane extract, 2: Ethyl acetate extract, 3: Methanol extract, 4: N-hexane extract of nampu leaves from tissue culture, 5: Ethyl acetate extract of nampu leaves from tissue culture, 6: Methanol extract of nampu leaves from tissue culture, (a) Visual, (b) ultraviolet (UV) 365 nm, (c) UV 254 nm, (d) H2SO4 spot appearance (Visual), (e) H2SO4 spot appearance (UV 365 nm), (f) Lieberman–Burchard spot appearance (Visual), (g) Lieberman–Burchard spot appearance (UV 365 nm), (h) Citroborate spot appearance (Visual), (i) Citroborate spot appearance (UV 365 nm), (j) Citroborate spot appearance (UV 254 nm), (k) FeCl3 spot appearance (Visual). [Click here to view] |
 | Figure 10: Thin layer chromatography using the mobile phase n-hexane: ethyl acetate (7:3) on the ZPT IBA: Kinetin. 1: N-hexane extract, 2: Ethyl acetate extract, 3: Methanol extract, 4: N-hexane extract of nampu leaves from tissue culture, 5: Ethyl acetate extract of nampu leaves from tissue culture, 6: Methanol extract of nampu leaves from tissue culture, (a) Visual, (b) ultraviolet (UV) 365 nm, (c) UV 254 nm, (d) H2SO4 spot appearance (Visual), (e) H2SO4 spot appearance (UV 365 nm), (f) Lieberman–Burchard spot appearance (Visual), (g) Lieberman–Burchard spot appearance (UV 365 nm), (h) Citroborate spot appearance (Visual), (i) Citroborate spot appearance (UV 365 nm), (j) Citroborate spot appearance (UV 254 nm), (k) FeCl3 spot appearance (Visual). [Click here to view] |
In the n-hexane fraction of the callus obtained from the kinetin-2,4-D medium (2:2 mg/L), a subsequent separation stage was conducted using preparative TLC, employing a method for the separation and adsorption of compounds on silica gel measuring 20 × 20 cm (total plate size). The application of the sample was performed along the length of the plate, resulting in a band-like appearance upon elution. TLC was undertaken to isolate and amplify the yield of the produced compounds. The results indicated a band with an Rf value of 0.72, which exhibited blue fluorescence under UV light at 365 nm; the band is faintly visible under visual inspection or at UV 254 nm. The product obtained from the TLC was scraped off and dissolved using the developing solvent, specifically a mixture of n-hexane and ethyl acetate (7:3), followed by filtration and subsequent TLC testing [Figure 11].
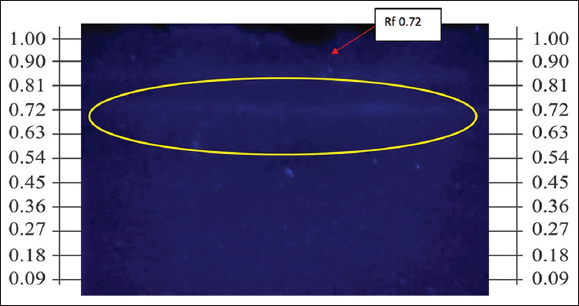 | Figure 11: Profile of the chromatogram for preparative thin layer chromatography of nampu leaf callus from kinetin - 2,4-d medium. [Click here to view] |
3.4. Identification of Isolate Structure
The identification of the initial stages of the isolate was conducted using UV-Vis spectrophotometry at wavelengths ranging from 200 to 400 nm, employing a solvent analogous to the developing agent, specifically n-hexane: ethyl acetate (7:3). The examination results indicated that the isolate exhibited absorbance at wavelengths of 255 nm and 292 nm. This absorbance suggests the presence of chromophoric groups within the isolate that undergo π-to-π* transitions, which also contribute to absorbance in the K band or conjugated bonds. The K band arises from compounds containing a conjugated system. All types of conjugated components exhibit strong absorption of UV radiation in the wavelength range of 220–350 nm [Figure 12].
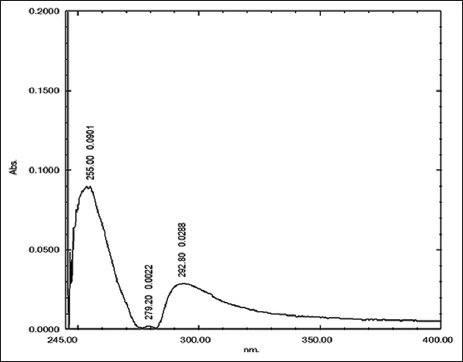 | Figure 12: Ultraviolet-visible spectrum of the isolate. [Click here to view] |
The identification of advanced stages using IR spectroscopy involves analyzing the functional groups of a compound within the range of 400–4000 cm−1. The spectrum obtained is a result of transitions between different vibrational energy levels when the sample is absorbed. The resulting data are derived from a graph of wavenumber, which continuously varies across a narrow region of the electromagnetic spectrum plotted against percentage transmittance (T%). The identification using this instrument reveals the presence of C=C bending and Ar-H at a wavenumber of 802.39 cm−1. The presence of C– C bending in aromatic compounds is observed at a wavenumber of 1226.73 cm−1. The C-H stretching in aliphatic -CH2- appears at 1365.60 cm−1. The C-H stretching in aliphatic -CH3- is indicated at a wavenumber of 1465.90 cm−1. The presence of carbonyl in ketones is demonstrated at a wavenumber of 1739.79 cm−1 [Figure 13 and Table 5].
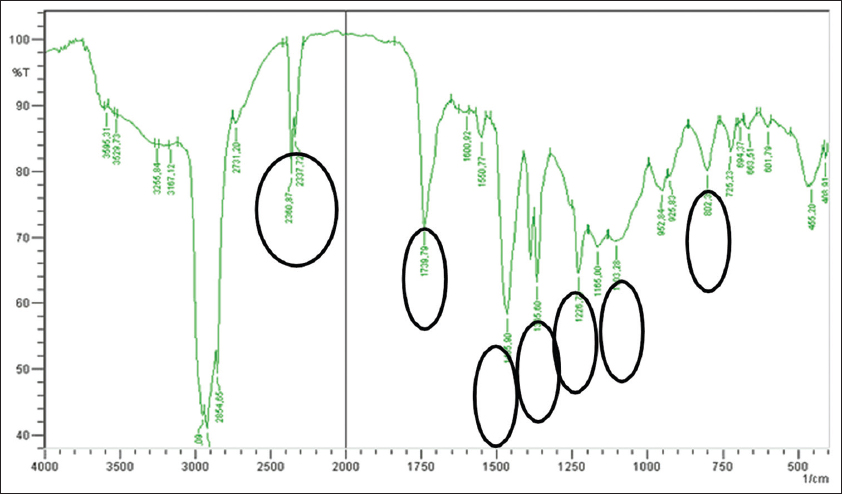 | Figure 13: Infrared spectroscopy spectrum of the isolate. [Click here to view] |
Table 5: Frequency range of nampu plant callus isolates.
| No | Frequency range (cm-1) | Functional group |
|---|---|---|
| 1 | 802.39 cm−1 | C=C bending and Ar-H |
| 2 | 1226.73 cm−1 | C–bending in aromatic compounds |
| 3 | 1365.60 cm−1 | -CH2- |
| 4 | 1465.90 cm−1 | -CH3- |
| 5 | 1739.79 cm−1 | carbonyl in ketones |
4. DISCUSSION
4.1. Optimization of Sterilization
The optimization of sterilization for the rhizome of the Nampu plant was conducted by varying the soaking time in a 2% sodium hypochlorite solution for durations of 5, 7, and 10 min. Despite these efforts, the results indicated that the sterilization process was not optimal, primarily due to the high water content in the rhizome, which resulted in browning and compromised tissue integrity. The browning observed in the rhizomes can be attributed to the enzymatic activity of polyphenol oxidases (PPO), which are activated in the presence of oxygen and can lead to the oxidation of phenolic compounds, resulting in discoloration [14]. In the context of plant tissue culture, it is crucial to strike a balance between sterilization efficacy and tissue quality preservation. Prolonged exposure to sodium hypochlorite 2% (durations 5 min) can exacerbate tissue damage, as observed in other studies where extended treatment durations led to increased browning and reduced viability of explants [16]. This suggests that while sodium hypochlorite 2% (durations 5 min) is effective for surface sterilization, its penetration into high-water-content tissues may be insufficient, leading to incomplete sterilization and subsequent browning. Furthermore, the choice of sterilization agent and its concentration play a significant role in the outcomes of the sterilization process.
The sterilization of nampu leaves was performed using sodium hypochlorite at a concentration of 2% v/v for 5 min, which was identified as a relatively ideal condition for effective sterilization. This specific concentration and duration were chosen to minimize potential damage to the plant cell tissues, which can occur with excessively high concentrations of hypochlorite or prolonged soaking times. In addition, a 70% alcohol solution was utilized for the sterilization of leaf explants for 1 min, a method that is well-established in plant tissue culture for its efficacy in reducing microbial contamination without significantly harming the plant tissues [17,18]. The choice of sodium hypochlorite 2% (durations 5 min) as a sterilizing agent is supported by its broad-spectrum antimicrobial properties, making it effective against a variety of pathogens. However, it is crucial to optimize the concentration and exposure time to avoid adverse effects on plant tissues. Studies have shown that higher concentrations or extended exposure can lead to cellular damage, browning, and reduced viability of explants [19,20]. Moreover, the use of 70% alcohol for a brief exposure time is consistent with established protocols in plant tissue culture. Alcohol acts by denaturing proteins and disrupting cell membranes, effectively reducing microbial load without the harsh effects associated with more potent disinfectants [21,22]. This method is effective in various plant species, providing a balance between sterilization efficacy and tissue preservation [23,24]. The findings from this study align with the literature, suggesting that the optimization of sterilization conditions is essential for maintaining the viability of plant tissues during in vitro culture. For example, Manea et al. noted that the effectiveness of sterilization methods can vary significantly based on the specific characteristics of the plant material, including moisture content and tissue structure [17]. This highlights the importance of tailoring sterilization protocols to the specific needs of different plant species. The sterilization of Nampu leaves using 2% sodium hypochlorite for 5 min, coupled with a 1-min exposure to 70% alcohol, represents a balanced approach to achieving effective sterilization while minimizing damage to plant tissues. Studies indicate that the soaking time can significantly influence the efficacy of sterilization; experimenting with durations of 5, 7, and 10 min in a 2% sodium hypochlorite solution demonstrated that increased exposure times can lead to browning and tissue integrity issues, particularly due to the high water content within the rhizome tissues The use of a 2% sodium hypochlorite solution for 5 min appears to be efficient for nampu leaves, variations in sterilization techniques are necessary for the rhizomes. These variations must accommodate the distinct properties of the rhizome to overcome the challenges of browning and integrity compromise.
4.2. Callus Induction
The observation that leaf explants without PGR treatment displayed curling and a yellowish-green color by day 33 highlights the critical role of hormonal regulation in plant tissue culture. The absence of PGRs can lead to suboptimal growth responses, as seen in various studies where explants exhibited poor morphogenic responses when not supplemented with appropriate growth regulators [25,26]. This aligns with findings that the source of the explant significantly influences in vitro responses, suggesting that endogenous hormone levels and their responsiveness can vary between different plant organs [25]. In the context of nampu leaf explants, curling and discoloration may indicate stress responses resulting from inadequate nutrient uptake or hormonal imbalance. Research has shown that leaf explants can regenerate more effectively when treated with a combination of auxins and cytokinins, which promote cell division and differentiation [26]. For instance, studies on other species have demonstrated that the addition of specific PGRs can enhance shoot regeneration and overall explant viability, thereby preventing the curling and yellowing observed in the control group [26]. Moreover, the use of sodium hypochlorite 2% as a sterilizing agent is crucial in preventing microbial contamination, which can further exacerbate stress responses in explants. Sodium hypochlorite 2% is known for its broad-spectrum antimicrobial activity, but its concentration and exposure time must be carefully optimized to avoid phytotoxic effects [27,28]. Inappropriate concentrations can lead to tissue damage, which may contribute to the curling and yellowing observed in the control explants [28]. Studies have indicated that lower concentrations of sodium hypochlorite 2% can be effective in maintaining tissue integrity while still providing adequate sterilization [28]. The physiological responses of plant tissues to sterilization and growth conditions are complex. They can be influenced by multiple factors, including the type of explant, the concentration of growth regulators, and the sterilization method employed [29]. For example, it has been reported that explants excised from younger tissues tend to regenerate more effectively due to higher levels of active growth hormones, which could mitigate the adverse effects observed in the control explants [29]. The curling and yellowish-green color of the nampu leaf explants without PGR treatment underscores the importance of hormonal regulation in plant tissue culture.
The development of nampu leaf explants using the PGRs NAA and BAP at a ratio of 1.5:10 mg/L on day 98 revealed significant morphological issues, including nearly curling leaves and a white appearance on the upper surface, with no callus formation observed. These observations suggest that the chosen concentrations and combinations of PGRs may not have been optimal for promoting healthy growth and regeneration in the explants. The curling of leaves can be indicative of several physiological stresses, often linked to hormonal imbalances or inadequate nutrient uptake. In plant tissue culture, the balance between auxins (like NAA) and cytokinins (like BAP) is crucial for regulating cell division and differentiation [30,31]. When the ratio of these hormones is not conducive to the specific needs of the plant tissue, it can lead to abnormal growth patterns such as curling and discoloration. For instance, studies have shown that excessive cytokinin levels relative to auxins can lead to uncontrolled cell division [32,33]. While excessive cytokinin levels can lead to issues such as shoot proliferation and related developmental disorders, the phenomenon of leaf curling has multifactorial origins that extend beyond hormonal influences. Nutrient balance and environmental factors play critical roles in determining leaf morphology and health, establishing a complex interplay that requires careful manipulation in agricultural practices to optimise both plant health and productivity [34]. High BAP concentrations can lead to excessive shoot formation, resulting in abnormal growth patterns, such as leaf curling. However, cytokinin-induced cell division typically leads to shoot formation rather than leaf curling. This phenomenon has been documented in various plant species where in vitro conditions lead to reduced photosynthetic efficiency due to low light levels or suboptimal nutrient conditions [35,36]. The absence of callus formation further indicates that the explants were not responding favorably to the applied PGRs, which is essential for successful regeneration in tissue culture [37,38]. In the context of Nampu leaf explants, the lack of callus formation could be attributed to the specific genotype’s response to the PGR combination used. Research has shown that the efficacy of PGRs can vary significantly among different plant species and even among cultivars within the same species [39]. For example, in Brassica species, the regeneration capacity is highly genotype-dependent, and specific combinations of BAP and NAA have been shown to yield better results in specific genotypes compared to others [30,40].
The timing of PGR application and the physiological state of the explants at the time of treatment can significantly influence the outcomes. Studies have indicated that younger tissues often respond better to PGR treatments compared to older tissues, which may have reduced regenerative capacity [41,42]. Therefore, the age and health of the explants used in this study could have played a significant role in the observed results. The development of nampu leaf explants under the specified PGR conditions highlights the complexity of plant tissue culture and the necessity for careful optimization of growth regulator concentrations and combinations. In plant tissue culture, the optimization of growth regulator combinations is crucial for effective regeneration and somatic embryogenesis. An essential aspect of this optimization process is the synergistic interaction between cytokinins and auxin hormones, particularly 2,4-D. Research indicates that the combination of cytokinins and 2,4-D plays a pivotal role in enhancing shoot regeneration efficiency in various plant species. For instance, Ivarson et al. report that auxins, including 2,4-D, when paired with cytokinins such as BAP or zeatin, lead to significantly higher shoot regeneration rates compared to the application of either hormone alone [43]. The interaction between high endogenous cytokinin levels and exogenous auxins is multifaceted. Elevated cytokinin levels can interfere with auxin function by modifying its biosynthesis, transport, and signaling pathways, leading to significant developmental implications [44]. The potential of fine-tuning PGR applications is supported by empirical evidence across various disciplines in plant sciences. As agricultural practices demand crops that can withstand climate variability and abiotic stresses, the nuanced application of PGRs specific to species requirements will be essential for sustainable cultivation practices [45].
The treatment of nampu leaf explants with 2,4-D and pineapple extract (0.5 mg/L: 15%) on day 68 resulted in no significant morphological changes, although a noticeable fading of leaf color was observed. This lack of morphological development, despite the presence of 2,4-D, suggests that the concentration and combination of growth regulators may not have been optimal for inducing callus formation or promoting growth in these explants. 2,4-D is a synthetic auxin commonly used in plant tissue culture to induce callus formation and promote cell division. However, its effectiveness can vary significantly depending on the concentration and the specific plant species being cultured [46,47]. In some cases, lower concentrations of 2,4-D are more effective for callus induction compared to higher concentrations, which can lead to tissue necrosis or other adverse effects [48,49]. The absence of callus formation in the current study may indicate that the concentration of 2,4-D used was insufficient to trigger the necessary physiological responses in the nampu leaf explants.
The fading of leaf color could be attributed to several factors, including the potential degradation of chlorophyll due to stress or inadequate nutrient uptake. Previous studies have indicated that prolonged exposure to certain growth regulators can lead to chlorosis, particularly when the explants are not receiving adequate light or nutrients [50,51]. The interaction between auxins and environmental conditions is critical, as the balance can significantly influence the physiological state of the explants [52]. Moreover, the combination of 2,4-D with other growth regulators, such as cytokinins, has been shown to enhance callus induction and growth in various plant species [53,54]. The lack of morphological changes in the nampu explants may suggest that the absence of cytokinin, which promotes cell division and shoot formation, could be a limiting factor in the regeneration process [55]. For instance, studies have demonstrated that the presence of cytokinins in conjunction with auxins can lead to improved callus formation and shoot regeneration, highlighting the importance of optimizing the hormonal balance in tissue culture media [56,57].
The endogenous levels of plant hormones in the explants themselves may also play a role in their response to exogenous growth regulators. High levels of endogenous auxins can inhibit the effectiveness of exogenous 2,4-D, potentially leading to a lack of response [58,59]. This phenomenon has been observed in other species, where the presence of high endogenous auxin levels correlated with reduced callus formation and regeneration capacity [60,61]. The lack of morphological changes and the fading of leaf color in nampu leaf explants treated with 2,4-D and pineapple extract suggest that the applied concentrations and combinations of growth regulators were not conducive to promoting growth or callus formation. Pineapple (Ananas comosus) extracts contain various bioactive compounds, such as bromelain and antioxidants, that may interact with PGRs. The antioxidant capacity of pineapple extracts is primarily attributed to the synergistic action of multiple compounds, which may amplify the action of PGRs by promoting metabolic pathways involved in plant growth [62]. For instance, studies have shown that PGRs, such as gibberellins, can enhance metabolic activities, including cellular respiration, which may be supported by the antioxidants present in pineapple extract, leading to improved growth performance under specific conditions [63]. Conversely, the interaction dynamics can be complex. High concentrations of pineapple extracts may lead to morphological changes and physiological disruptions caused by osmotic stress or nutrient imbalances. This phenomenon has been observed in various plant studies, suggesting that the balance between beneficial and inhibitory effects depends on the concentrations used, which may overwhelm the plant’s physiological resilience [64].
The results of tissue culture using the PGRs 2,4-D and Kinetin in a 2:2 mg/L ratio on day 33 indicated that the leaf explants exhibited curling, and the formed callus was brown. One of the primary studies worth noting is the work by Raomai et al., who successfully reported the in vitro propagation of H. aromatica via rhizome axillary bud multiplication. Utilizing MS medium supplemented with cytokinins, this research established multiple shoot formation, crucial for the micropropagation of this aromatic medicinal herb [12]. The use of PGR 2,4-D and Kinetin in a 2:2 mg/L in inducing Homalomena spp. callus. These observations highlight several critical aspects of plant tissue culture, particularly regarding the effects of hormonal balance on explant morphology and callus quality. The curling of the leaf explants can be attributed to the hormonal imbalance created by the specific concentrations of 2,4-D and Kinetin used. While 2,4-D is an auxin that promotes cell elongation and division, excessive levels can lead to abnormal growth patterns, including curling and necrosis [65,66].
Studies have shown that high concentrations of auxins can induce stress responses in plant tissues, resulting in morphological [67]. In contrast, Kinetin, a cytokinin, is known to promote cell division and shoot formation. However, when used in conjunction with high levels of auxins, it can lead to an imbalance that favors callus proliferation over normal shoot development, potentially resulting in the observed curling [13,68]. The browning of the callus is another significant concern, as it often indicates tissue stress or senescence. Browning in callus cultures can be caused by several factors, including phenolic compound accumulation, which is a typical response to wounding or tissue damage [45,69]. The presence of phenolic compounds can lead to oxidative stress, resulting in browning and reduced viability of the callus [70]. In this context, the combination of 2,4-D and Kinetin may have triggered such stress responses, leading to the observed browning of the callus tissue. Moreover, the choice of PGRs and their concentrations is crucial for successful callus induction and regeneration. Previous studies have indicated that a balanced ratio of auxins and cytokinins is essential for optimal callus formation and subsequent plant regeneration [71,72]. For instance, lower concentrations of 2,4-D combined with Kinetin have been shown to enhance callus quality and reduce browning compared to higher concentrations [73]. This suggests that the specific ratio of 2,4-D to Kinetin used in this study may need to be adjusted to improve the overall health and viability of the callus.
The physiological state of the explants at the time of treatment can significantly influence their response to PGRs. Young, actively growing tissues generally respond better to hormonal treatments than older or stressed tissues [74,75]. Therefore, the age and health of the nampu leaf explants used in this study could have contributed to the adverse outcomes observed. The curling of the leaf explants and the browning of the callus in response to the 2,4-D and Kinetin treatment underscores the importance of optimizing PGR concentrations and ratios in plant tissue culture. Callus browning poses a significant challenge in plant tissue culture, adversely affecting the proliferation and viability of callus cultures, particularly in species that are rich in secondary metabolites. This browning can be attributed to various factors, including enzymatic reactions, oxidative stress, and the accumulation of phenolic compounds. The roles of PPO and peroxidase in this process have been extensively studied, highlighting their contributions to the browning phenomenon during in vitro culture [76]. In applying strategies to mitigate browning, such as pH adjustment and the inclusion of sucrose, researchers have found that optimizing culture conditions can enhance cell growth and reduce phenolic stress, thereby minimizing browning [77].
The treatment of nampu leaf explants with IBA and Kinetin at a ratio of 0.5:1 mg/L on day 41 resulted in curling of the leaves and the formation of brown callus. These observations raise important considerations regarding the effects of PGRs on the morphology and quality of callus tissue in plant tissue culture. The curling of the leaf explants can be attributed to the hormonal interactions between IBA and Kinetin. IBA is primarily an auxin that promotes root formation and cell elongation, while Kinetin, a cytokinin, is involved in cell division and shoot development [51,78]. When these two hormones are applied in an unbalanced ratio, as seen in this study, the resulting physiological stress can lead to abnormal growth patterns, such as leaf curling [79]. Previous research has indicated that a higher concentration of cytokinins relative to auxins can lead to excessive cell division without adequate elongation, resulting in distorted leaf morphology [79,80]. The presence of brown callus is another critical observation, often indicative of tissue stress or necrosis. Browning in callus cultures can occur due to the accumulation of phenolic compounds, which are typically produced in response to wounding or stress [26,81]. The combination of IBA and Kinetin may have triggered such stress responses, leading to oxidative damage and browning of the callus tissue [82]. This phenomenon has been documented in other studies where high auxin concentrations resulted in increased phenolic compound production, which in turn affected callus viability and morphology [83,84]. Moreover, the specific concentrations of IBA and Kinetin used in this study may not have been optimal for promoting healthy callus formation. Research has shown that the effectiveness of PGRs can vary significantly depending on the plant species and the specific tissue being cultured [57,85]. For example, in some species, lower concentrations of IBA combined with Kinetin have been found to enhance callus quality and reduce browning compared to higher concentrations [86,87]. This suggests that the current hormonal treatment may require optimization to improve the overall health and viability of the callus.
The age and health of the nampu leaf explants used in this study could have contributed to the negative outcomes observed. The curling of the leaf explants and the formation of brown callus in response to IBA and Kine might have been caused by the age and health of the explants [88,89]. NAA is an auxin that promotes cellular elongation and division, while BAP is a cytokinin that stimulates cell division and shoot formation. However, studies indicate that while NAA can work effectively in certain combinations, its concentration and pairing with compatible cytokinins like BAP can lead to suboptimal outcomes. Vahedi et al. found that a more synergistic effect on callus formation is generally obtained with 2,4-D combined with kinetin compared to the results obtained with NAA and BAP, which led to inferior callus formation [90].
4.3. Analysis of Secondary Metabolites from Callus
The observation was that tissue-cultured nampu leaves were not visually detectable due to their low metabolite content. At the same time, spots were observed under UV light at 365 nm, indicating the potential for secondary metabolite production in these cultures. The use of 10% H2SO4 in methanol as a visualization agent, followed by heating, facilitated the detection of organic compounds, as evidenced by the color change post-heating. This method is recognized for its effectiveness in revealing the presence of various secondary metabolites in plant tissues [91,92]. The low visibility of the tissue-cultured nampu leaves may suggest that the concentration of secondary metabolites was insufficient for detection without the aid of visualization techniques. This aligns with findings that tissue culture can sometimes lead to reduced metabolite production compared to field-grown plants, particularly when the culture conditions are not optimized [93,94]. The presence of spots under UV light indicates that some metabolites, possibly flavonoids or phenolic compounds, were produced, albeit in low concentrations. This is consistent with studies that have shown that tissue culture can yield secondary metabolites, but the levels can vary significantly based on the specific conditions and treatments applied [92]. The color change observed in the Nampu leaves after heating with H2SO4 is indicative of the presence of certain organic compounds, a standard method used to confirm the presence of secondary metabolites such as flavonoids and alkaloids [95]. The heating process not only evaporates the solvent but also facilitates the reaction of the visualization agent with the metabolites, leading to a color change that can be quantified or compared against standards [96].
In contrast, the slight brown color change in the nampu leaves without tissue culture suggests that these leaves may have undergone some degree of oxidative stress or degradation of chlorophyll, which can occur in non-cultured leaves due to environmental factors [97]. The differences in metabolite profiles between tissue-cultured and non-cultured leaves highlight the impact of in vitro conditions on plant physiology and secondary metabolite production. Furthermore, the use of specific visualization agents, such as Lieberman–Burchard, Anisaldehyde, citroborate, and FeCl3, in addition to the universal visualization agent, provides a comprehensive approach to analyzing the metabolite profile of the Nampu leaves. Each of these agents is known to react with specific classes of compounds, allowing for a more detailed understanding of the secondary metabolites present [98,99]. For example, Lieberman–Burchard is often used to detect sterols and terpenes, while Anisaldehyde is effective for identifying phenolic compounds [99].
The detection of steroid compounds in Nampu leaves and tissue-cultured samples using the Lieberman–Burchard reagent, which resulted in a visual color change to pink, indicates the presence of sterols. This color reaction is a well-established method for identifying steroids and terpenoids in plant extracts [100,101]. The Lieberman–Burchard test is particularly sensitive and can effectively differentiate between various classes of compounds, providing a reliable means of detecting sterols in both fresh and tissue-cultured plant materials [102,103]. The observation of brown callus in the tissue-cultured Nampu leaves suggests that the tissue may be undergoing oxidative stress or necrosis, which is often associated with high levels of phenolic compounds. The use of FeCl3 as a visualization agent confirmed the presence of polyphenolic compounds, as indicated by the formation of a black color on heating [104,105]. This reaction is characteristic of phenolic compounds, which are known to accumulate in response to stress and can contribute to browning in plant tissues [106]. The presence of these compounds is significant, as they are often associated with various biological activities, including antioxidant properties and potential health benefits [107,108].
Furthermore, the detection of flavonoids using citroborate, evidenced by bright blue fluorescence under UV light at 365 nm, highlights the diversity of secondary metabolites present in the Nampu leaves. Flavonoids are known for their antioxidant, anti-inflammatory, and antimicrobial properties, making them valuable components in medicinal plants [109]. The ability to visualize flavonoids through specific reagents underscores the importance of phytochemical screening in assessing the potential therapeutic applications of plant extracts [110]. The combination of different visualization agents, including Lieberman–Burchard, FeCl3, and citroborate, provides a comprehensive approach to analyzing the metabolite profile of nampu leaves. Each agent targets specific classes of compounds, allowing for a more detailed understanding of the secondary metabolites present in both tissue-cultured and non-cultured samples [111,112]. This multifaceted approach is essential for identifying bioactive compounds that may contribute to the medicinal properties of the nampu plant.
The application of preparative TLC to separate and analyze the n-hexane fraction of callus obtained from the kinetin; 2,4-D medium (2:2 mg/L) yielded significant insights into the composition of secondary metabolites in nampu callus. The successful separation of compounds, indicated by a band with an Rf value of 0.88 that exhibited blue fluorescence under UV light at 365 nm, suggests the presence of specific organic compounds, likely flavonoids or other phenolic derivatives, which are known for their bioactive properties [113,114]. The use of TLC as a separation technique is well-established due to its simplicity, cost-effectiveness, and ability to provide qualitative and quantitative data on complex mixtures [115,116]. The band-like appearance observed upon elution indicates effective separation, which is crucial for subsequent analyses and identification of bioactive compounds. The ability to visualize compounds under UV light further enhances the utility of TLC in phytochemical investigations, allowing researchers to identify specific classes of metabolites based on their fluorescence characteristics [113,114]. The observation that no band was detectable under visual inspection or at UV 254 nm highlights the importance of using appropriate visualization techniques tailored to the specific compounds of interest. This aligns with findings that certain metabolites may only be detectable under specific conditions or with specific reagents, emphasizing the need for a multifaceted approach to metabolite analysis [117,118]. The subsequent scraping of the product and dissolution in a solvent mixture of n-hexane and ethyl acetate (7:3) demonstrates a practical method for isolating compounds for further characterization, which is essential for understanding their potential biological activities [119].
4.4. Identification of Isolate Structure
The identification of the initial stages of the isolate using UV-Vis spectrophotometry, particularly at wavelengths 255 nm and 292 nm, provides valuable insights into the chemical composition of the nampu leaves and the callus derived from tissue culture. The absorbance at these wavelengths suggests the presence of chromophoric groups that undergo π-to-π* transitions, indicative of compounds with conjugated double bonds [120]. The K band, associated with conjugated systems, is known to exhibit strong absorption in the UV range of 220 to 350 nm, further supporting the presence of these metabolites [120]. The use of n-hexane and ethyl acetate (7:3) as a solvent for the separation of compounds is a common practice in phytochemical analysis, as this solvent mixture effectively extracts non-polar and moderately polar compounds, which are often responsible for the bioactive properties of plant extracts [121]. The subsequent TLC analysis, which resulted in a band with an Rf value of 0.72, indicates the successful separation and isolation of specific compounds from the complex mixture present in the callus. The ability to visualize these compounds under UV light at 365 nm is particularly significant, as it allows for the identification of compounds that may possess antioxidant or antimicrobial properties [122]. The observation that no band was visible under UV light at 255 nm further emphasizes the specificity of the compounds detected at 365 nm, suggesting that the isolated metabolites may have unique structural features that allow them to absorb UV light at this wavelength while being less detectable at lower wavelengths [123].
The identification of advanced stages of the isolate using IR spectroscopy provides critical insights into the functional groups present in the nampu leaves and callus derived from tissue culture. The analysis conducted within the range of 400–4000 cm−1 revealed significant absorbance peaks, indicating the presence of various functional groups that contribute to the chemical profile of the isolate. The absorbance at specific wavenumbers, such as 802.39 cm−1, suggests the presence of alkenes and aromatic compounds, which are known for their diverse biological activities [124]. The detection of C–H stretching in aliphatic -CH2 bonds further supports the identification of organic compounds. The detection of C–C bonds in aliphatic linkages at wavenumbers ranging from 2360.87 cm−1 to 2337.72 cm−1 emphasises the diversity of organic compounds present in the isolate. This diversity is crucial for understanding the potential health benefits of the nampu plant, as many of these compounds exhibit antioxidant, anti-inflammatory, and antimicrobial properties [125]. The use of IR spectroscopy as a tool for identifying functional groups is well-documented, and its application in this study highlights its importance in phytochemical analysis [126]. The methodology employed, including the use of n-hexane: ethyl acetate (7:3) as a solvent, is effective for extracting a wide range of organic compounds, particularly those with varying polarities. This solvent system facilitates the separation of both polar and non-polar compounds, allowing for a comprehensive analysis of the metabolite profile [127]. The subsequent analysis of the isolated compounds through TLC and UV-Vis spectrophotometry further enhances the understanding of the chemical composition and potential bioactivity of the Nampu extracts [128]. The absorbance at a wavenumber of 1739.79 cm−1 in the IR spectrum is indicative of a carbonyl (C=O) group. This characteristic is particularly relevant in the study of steroid compounds, as the presence of the C=O group is frequently associated with the structural attributes of steroids and other organic compounds such as ketones and aldehydes. The detection of this functional group can facilitate the identification and characterization of steroids through techniques such as FTIR, which is commonly employed to discern functional groups in various chemical compounds, including steroids [129].
The identification of steroid compounds within the callus of nampu, as evidenced by the TLC profile and IR spectroscopy, provides significant insights into the phytochemical composition of this plant. The positive reaction observed with the Liebermann–Burchard reagent indicates the presence of sterols, which are important bioactive compounds known for their various health benefits, including anti-inflammatory and cholesterol-lowering effects [130,131]. The absorbance at a wavenumber of 1739.79 cm−1 in the IR spectrum suggests the presence of a carbonyl (C=O) group, which is characteristic of many steroid compounds [132]. This finding aligns with previous studies that have utilized IR spectroscopy to confirm the presence of functional groups associated with phytochemicals in various plant species [133,134]. The TLC analysis, which resulted in a distinct band corresponding to steroid compounds, further supports the notion that Nampu leaves and callus tissue contain valuable secondary metabolites. The Rf value obtained during the TLC process can provide insights into the polarity and structural characteristics of the isolated compounds [135]. The ability to visualize these compounds using specific reagents enhances the understanding of their potential applications in medicine and nutrition [136]. Moreover, the presence of other functional groups, such as those indicated by the C-O stretching at 1103.28 cm−1, suggests that the isolated compounds may also include alcohols or ethers, which can contribute to the overall bioactivity of the extracts [137]. The identification of these compounds is crucial, as they may play a role in the plant’s antioxidant properties, which are beneficial for human health [138]. The combination of TLC and IR spectroscopy provides a robust methodology for the identification and characterization of bioactive compounds in plant extracts. This approach has been widely utilized in phytochemical research to isolate and analyze secondary metabolites, facilitating the discovery of new therapeutic agents [139]. The identification of steroid compounds and other metabolites in the callus of nampu highlights the potential of this plant as a source of natural products with pharmacological significance.
5. CONCLUSION
The findings from this study demonstrate that the leaves of nampu (H. rostrata Schott) exhibit a positive growth response when induced with MS medium supplemented with Kinetin and 2,4-D at a concentration ratio of 2:2 mg/L. The successful induction of callus from nampu leaves signifies the potential for in vitro propagation and the enhancement of secondary metabolite production, which could be beneficial for both research and commercial applications. The distinct TLC profile of the callus derived from nampu leaves, compared to non-cultured leaves, indicates the production of unique compounds as a result of tissue culture. The pale purple coloration observed on visualization with the Liebermann–Burchard reagent suggests the presence of steroidal compounds, which are known for their various biological activities and therapeutic potential. This is further supported by the identification of chromophoric groups through UV-Vis spectrophotometry and the detection of carbonyl groups at a wavenumber of 1739.79 cm−1 through FTIR spectroscopy. Leveraging additional identification methods, such as mass spectrometry and nuclear magnetic resonance, ensures a higher degree of accuracy and confidence in compound identification within chemical analyses.
6. ACKNOWLEDGMENTS
Jenderal Achmad Yani University for providing the necessary support and facilities for this study.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. CONFLICT OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All the data are available with the authors and shall be provided upon request.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors confirm that there was no use of artificial intelligence (AI)-assisted technology for assisting in the writing or editing of the manuscript and no images were manipulated using AI.
REFERENCES
1. Faramayuda F. Identification of secondary metabolites compounds from nampu (Homalomena rosrata Griff.). Elkawnie. 2022;8(1):12.[CrossRef]
2. Aprilianingsih R, Wahidah BF, Hariri MR. DNA barcode of Homalomena pexa inferred from internal transcribed spacer region. J Ris Biol Dan Apl. 2022;4(2):69-74.[CrossRef]
3. Ravipati AS, Zhang L, Koyyalamudi SR, Jeong SC, Reddy NV, Bartlett JR, et al. Antioxidant and anti-inflammatory activities of selected Chinese medicinal plants and their relation with antioxidant content. BMC Complement Altern Med. 2012;12(1):173.[CrossRef]
4. Ridley C, Day J, Smith AG. Cryopreservation studies of an artificial co-culture between the cobalamin-requiring green alga Lobomonas rostrata and the Bacterium Mesorhizobium loti. J Appl Phycol. 2017;30(2):995-1003.[CrossRef]
5. Miguez-Salas O, Vardaro M, Rodriguez-Tovar FJ, Pérez-Claros JA, Huffard CL. Deep-sea echinoid trails and seafloor nutrient distribution:Present and past implications. Front Mar Sci. 2022;9:903864.[CrossRef]
6. Faramayuda F, Mariani TS, Elfahmi E, Sukrasno S. Influence of elicitation and precursors on major secondary metabolite production in cultures of purple Orthosiphon aristatus Blume Miq. Biocatal Agric Biotechnol. 2022;42:102324.[CrossRef]
7. Skrzypczak-Pietraszek E, Urbanska A, Zmudzki P, Pietraszek J. Elicitation with methyl jasmonate combined with cultivation in the plantformTM temporary immersion bioreactor highly increases the accumulation of selected centellosides and phenolics in Centella asiatica (L.) Urban shoot culture. Eng Life Sci. 2019;19(12):931-43.[CrossRef]
8. Shoja HM, Shishavani H. Effects of different hormonal treatments on growth parameters and secondary metabolite production in organ culture of Hyssopus officinalisL. Biotechnologia. 2021;102(1):33-41.[CrossRef]
9. Wakefield JC, Hassan HM, Jaspars M, Ebel R, Rateb ME. Dual induction of new microbial secondary metabolites by fungal bacterial co-cultivation. Front Microbiol. 2017;8:1284.[CrossRef]
10. Khattab AI, Babiker EH, Saeed HA. Streptomyces:Isolation, optimization of culture conditions and extraction of secondary metabolites. Int Curr Pharm J. 2016;5(3):27-32.[CrossRef]
11. Yanhua T, Liu F, Guo D, Fan L, ZhenXian Z, Xue Y, et al. Molecular characterization of flavanone 3-hydroxylase gene and flavonoid accumulation in two chemotyped safflower lines in response to methyl jasmonate stimulation. BMC Plant Biol. 2016;16(1):132.[CrossRef]
12. Raomai S, Kumaria S, Tandon P. In vitro propagation of Homalomena aromaticaschott., endangered aromatic medicinal herb of Northeast India. Physiol Mol Biol Plants. 2013;19(2):297-300.[CrossRef]
13. Masekesa RT, Gasura E, Kujeke GT, Ngadze E, Chidzwondo F. Induction of somatic embryogenesis and organogenesis in zimbabwean sweet potato (Cv Brondal). Adv Agric. 2021;2021:1-8.[CrossRef]
14. Rodiansah A, Sinuraya M, Hanafiah DS, Butar-Butar SS, Pohan A. Optimalisation of in vitro sterilisation methods for north sumatra local garlic (Allium sativum L.). Iop Conf Ser Earth Environ Sci. 2024;1362(1):12007.[CrossRef]
15. Slazak B, Sliwinska E, Saluga M, Ronikier M, Bujak J, Slomka A, et al. Micropropagation of Viola uliginosa (Violaceae) for endangered species conservation and for somaclonal variation-enhanced cyclotide biosynthesis. Plant Cell Tissue Organ Cult. 2014;120(1):179-90.[CrossRef]
16. Shetty SC, Narasimhan S. Sodium dichloroisocyanurate:An eco-friendly chemical alternative for media autoclaving and explant sterilisation in plant tissue culture. Int J Res Pharm Sci. 2021;12(1):107-12.[CrossRef]
17. Manea A, Leishman MR, Geraghty DM, Perera SD, O'Hare JA, Gil-Fernández M, et al. The effects of sterilisation on abiotic soil properties and the challenge this poses to untangling the influence of abiotic and biotic soil variables on plant growth. J Soil Sci Plant Nutr. 2024;24(3):4929-34.[CrossRef]
18. Holland JD, Webster GE, Rooney P, Wilshaw S, Jennings LM, Berry H. Effects of chemical and radiation sterilisation on the biological and biomechanical properties of decellularised porcine peripheral nerves. Front Bioeng Biotechnol. 2021;9:660453.[CrossRef]
19. Raynaud T, Pivato B, Siol M, Spor A, Blouin M. Soil microbes drive the effect of plant species and genotypic diversity interaction on productivity. Plant Soil. 2021;467(1-2):165-80.[CrossRef]
20. Abdelfattah FA, Gaede W. Influences on the sensitivity of real-time PCR for the detection of bovine DNA in heat-sterilised feedstuffs. Arch Anim Nutr. 2011;65(3):175-85.[CrossRef]
21. Antons J, Marascio MG, Aeberhard P, Weißenberger G, Hirt-Burri N, Applegate LA, et al. Decellularised tissues obtained by a CO2-philic detergent and supercritical CO2. Eur Cell Mater. 2018;36:81-95.[CrossRef]
22. Keerthana SG, Gunnaiah R, Patil S, Pattepur S, Prakasha DP, Nanjappanavar A, et al. Studies on in vitro surface sterilisation and antioxidants on pomegranate (Punica granatum L.) Cv. Bhagwa. Plant Cell Biotechnol Mol Biol. 2024;25(3-4):72-80.[CrossRef]
23. Rynio P, Galant K, Wójcik L, Grygorcewicz B, Kazimierczak A, Falkowski A, et al. Effects of sterilization methods on different 3D printable materials for templates of physician-modified aortic stent grafts used in vascular surgery-a preliminary study. Int J Mol Sci. 2022;23(7):3539.[CrossRef]
24. Liang M, Liu X, Parker IM, Johnson D, Zheng Y, Luo S, et al. Soil microbes drive phylogenetic diversity-productivity relationships in a subtropical forest. Sci Adv. 2019;5(10):eaax5088.[CrossRef]
25. Piovan A, Cassina G, Filippini R. Crambe tataria:Actions for ex situconservation. Biodivers Conserv. 2010;20(2):359-71.[CrossRef]
26. Mahroofa J, Singh S, Farhana M, Ahmad I. Effect of explant source and different hormonal combinations on in vitro regeneration of Heracleum candicans wall:An important medicinal herb. Afr J Biotechnol. 2019;18(28):707-12.[CrossRef]
27. Hesami M, Naderi R, Yoosefzadeh-Najafabadi M. Optimizing sterilization conditions and growth regulator effects on in vitro shoot regeneration through direct organogenesis in Chenopodium quinoa. Biotechnologia. 2018;99(1):49-57.[CrossRef]
28. Dramane K, Samuel KO, Oumar S, N'Guessan AR, Yeo N, Kouakou TH. Optimization of bud disinfection technique and influence of growth regulators on micropropagation in ginger (Zingiber officinale Rosc.). Int J Biol Chem Sci. 2023;16(6):2892-904.[CrossRef]
29. Tymoszuk A, Antkowiak M. In vitro adventitious organogenesis in Ajania pacifica(Nakai) Bremer Et Humphries. Biotechnologia. 2018;99(4):335-43.[CrossRef]
30. Mollika SR, Sarker RH, Hoque MI. In vitro plant regeneration in BrassicaSpp. Plant Tissue Cult Biotechnol. 2011;21(2):127-34.[CrossRef]
31. Goswami B, Khan S, Banu TA, Akter S, Islam M, Habib A. In vitro mass propagation of Withania somnifera (L.) an important medicinal plant of Bangladesh. Bangladesh J Bot. 2022;51(2):191-7.[CrossRef]
32. Wani M, Pande S, More NV. Callus induction studies in Tridax procumbens L. Int J Biotechnol Appl. 2010;2(1):11-4.[CrossRef]
33. Banu TA, Goswami B, Akter S, Islam M, Tanjin T, Habib A, et al. High frequencyin vitro regeneration of Gynura procumbens (Lour.) Merr. Plant Tissue Cult Biotechnol. 2017;27(2):207-16.[CrossRef]
34. Li Y, Liu F, Li P, Wang T, Zheng C, Hou B. An arabidopsis cytokinin-modifying glycosyltransferase UGT76C2 improves drought and salt tolerance in rice. Front Plant Sci. 2020;11:560696.[CrossRef]
35. Surgun-Acar Y, Ayaz ÖB, Bürün B. Relikt endemik liquidambar orientalis (Miller)'in hipokotil ve kotiledon eksplantlarindan adventif sürgün olusumu. Mugla J Sci Technol. 2018;4(2):137-42.[CrossRef]
36. Kazeroonian R, Mousavi A, Jari SK, Tohidfar M. Factors influencingin vitro organogenesis of Chrysanthemum morifolium Cv. “resomee splendid.“Iran J Biotechnol. 2018;16(2):132-9.[CrossRef]
37. Prasuna VN, Reddy CM, Lakshmi PV, Gopal MR, Purna GS, Srinivas B. Indirect shoot organogenesis from the leaf explants of Oxalis corniculata (L.) - an important medicinal plant. Plant Tissue Cult Biotechnol. 2022;32(2):181-91.[CrossRef]
38. Wahab U, Ashfaq M, Sajjad M, Shaheen S, Sadique R, Ali M, et al. Effects of different plant growth regulators on in vitro growth of Aloe vera L. Bangladesh J Bot. 2020;49(1):159-62.[CrossRef]
39. Kumar P, Srivastava DK. High frequency organogenesis in hypocotyl, cotyledon, leaf and petiole explants of broccoli (Brassica oleraceaL. Var. Italica), an important vegetable crop. Physiol Mol Biol Plants. 2015;21(2):279-85.[CrossRef]
40. Maloth GS, Marka R, Nanna RS. Effect of plant growth regulators on somatic embryogenesis and plantlet development of Turkey berry (Solanum torvum SW). European J Med Plants. 2021;32:1-8.[CrossRef]
41. Raju RI, Hashi AK, Jazib A, Hossain MT. Micropropagation of Alocasia amazonica through indirect shoot organogenesis. Plant Tissue Cult Biotechnol. 2022;32(1):13-20.[CrossRef]
42. Aasim M. In vitro shoot regeneration of NAA-pulse treated plumular leaf explants of cowpea. Not Sci Biol. 2010;2(2):60-3.[CrossRef]
43. Ivarson E, Ahlman A, Li X, Zhu L. Development of an efficient regeneration and transformation method for the new potential oilseed crop Lepidium campestre. BMC Plant Biol. 2013;13(1):115.[CrossRef]
44. Kurepa J, Shull TE, Smalle J. Antagonistic activity of auxin and cytokinin in shoot and root organs. Plant Direct. 2019;3(2):00121.[CrossRef]
45. Bashmakov DI, Lukatkin AS, MiliauskieneJ, Duchovskiene L, Duchovskis P. The efficiency of pre-treatment of maize seeds with plant growth regulators for resistance of maize seedlings to zinc ions. Zemdirbyste Agric. 2021;108(2):125-32.[CrossRef]
46. Jyothi KR, Josekutty PC, Shobha P, Mathew R, Kinza S, Subhashini C, et al. Callus induction and RAPD analysis of Simarouba glauca DC. Afr J Biotechnol. 2014;13(53):4766-74.[CrossRef]
47. Wahyuni DK, Purnobasuki H, Kuncoro EP, Ekasari W. Callus induction of Sonchus arvensis L. its antiplasmodial activity. Afr J Infect Dis. 2020;14(1):1-7.[CrossRef]
48. Ravikumar A, John J. In vitro antioxidant activity of Anisochilus carnosus leaf, stem and callus. Environ Exp Biol. 2022;20(3):165-70.[CrossRef]
49. Sakpere AM, Ayisire ER, Abioye O. Potential of Launea taraxacifolia (Willd) Amin Ex. C. Jeffrey for in vitro regeneration. Not Sci Biol. 2011;3(3):93-6.[CrossRef]
50. Tarrahi R, Rezanejad F. Callogenesis and production of anthocyanin and chlorophyll in callus cultures of vegetative and floral explants in Rosa gallica and Rosa hybrida(Rosaceae). Turk J Botany. 2013;37:1145-54.[CrossRef]
51. Junairiah J, Purnomo P, Utami ES, Ni'matuzahroh N, Sulistyorini L. Callus induction of Piper Betle L. nigra using 2,4-dichlorofenoxyacetic acidand 6-benzil aminopurin. Biosaintifika J Biol Biol Educ. 2018;10(3):588-96.[CrossRef]
52. Castro AH, Braga KQ, Sousa FM, Coimbra MC, Chagas RC. Callus induction and bioactive phenolic compounds production from Byrsonima verbascifolia (L.) DC. (Malpighiaceae). Rev Ciên Agron. 2016;47(1):143-151.[CrossRef]
53. Shoja AA, Çirak C, Ganjeali A, Cheniany M. Stimulation of phenolic compounds accumulation and antioxidant activity in in vitro culture of Salvia tebesana bunge in response to nano-TiO2 and methyl jasmonate elicitors. Plant Cell Tissue Organ Cult. 2022;149(1-2): 423-40.[CrossRef]
54. Sakpere AM, Ajayi SA, Adelusi AA. Effect of growth regulators and explant types on callus induction in Telfairia occidentalis Hook F. Afr J Biotechnol. 2014;13(20):2015-21.[CrossRef]
55. Shrestha U, Pant B. Production of bergenin, an active chemical constituent in the callus of Bergenia ciliata (Haw.) Sternb. Bot Orient J Plant Sci. 2012;8:40-4.[CrossRef]
56. Enrique AC, Lecona-Guzm CA, Antonio GM, Manuel ME, Carmen SG, Fima ML, et al. Effect of water stress induced by polyethylene glycol on growth, proline accumulation in Agave americana L. Phyton (B Aires). 2023;92(2):629-43.[CrossRef]
57. Kumari R. Induction of callus from different explants of Bacopa monnieri and effect of adjuvant on the growth rate of the calli. Indian J Sci Res. 2019;10(1):113.[CrossRef]
58. Inderiati S, Yanti FN, Mentari ER. Induction of morphogenic callus and regeneration of sugarcane (Saccharum officinarum L.) in vitro. Agriprima J Appl Agric Sci. 2021;5(1):61-7.[CrossRef]
59. Dangol SD, Yel I, Çaliskan ME, Bakhsh A. Manipulating genome of diploid potato inbred line Solanum chacoense M6 using selectable marker gene. Turk J Agric For. 2020;44(4):399-407.[CrossRef]
60. Budisantoso I, Amalia N, Kamsinah K.In vitro callus induction from leaf explants of VandaSp Stimulated by 2,4-D. Biosaintifika J Biol Biol Educ. 2017;9(3):492.[CrossRef]
61. Saptari RT, Sinta MM, Budiani A. In vitropropagation of Anthurium adreanumCv. Nitta through organogenesis. Agrivita J Agric Sci. 2017;39(2):192-200.[CrossRef]
62. Jovanovic M, Milutinovic M, Kostic M, Miladinovic B, Kitic N, Brankovic S, et al. Antioxidant capacity of pineapple (Ananas comosus (L.) Merr.) extracts and juice. Lek Sirovine. 2018;(38):27-30.[CrossRef]
63. Huang X, Rao G, Peng X, Xue Y, Hu H, Feng N, et al. Effect of plant growth regulators DA-6 and COS on drought tolerance of pineapple through bromelain and oxidative stress. BMC Plant Biol. 2023;23(1):180.[CrossRef]
64. Ilmiah M, Anggarani MA. The effect of PGR red onion extract on the growth and yield of green mustard plants (Brassica juncea L.). J Pijar Mipa. 2023;18(6):1006-11.[CrossRef]
65. Iriawati I, Puspita MI, Rodiansyah A. In vitro regeneration of foxtail millet (Setaria italica (L.) Beauv.) Cv. Buru Hotong. J Math Fundam Sci. 2017;49(2):171.[CrossRef]
66. Zuldin NN, Said IM, Noor NM, Zainal Z, Kiat CJ, Ismail I. Induction and analysis of the alkaloid mitragynine content of a Mitragyna speciosa suspension culture system upon elicitation and precursor feeding. Sci World J. 2013;2013(1):209434.[CrossRef]
67. Li Y, Zhao J, Chen H, Xia Y, Li H, Zhang Y, et al. Plant regeneration via callus-mediated organogenesis in commercial variety of Chuanbeichai No. 1 in Bupleurum chinense DC. Plant Biotechnol Rep. 2022;17(2):159-69.[CrossRef]
68. Ajijah N, Hartati RS, Rubiyo R, Sukma D, Sudarsono S. Effective cacao somatic embryo regeneration on kinetin supplemented DKW medium dnd somaclonal variation assessment using SSRs markers. Agrivita J Agric Sci. 2016;38(1):80-92.[CrossRef]
69. Raj D, Kokotkiewicz A, Drys A, Luczkiewicz M. Effect of plant growth regulators on the accumulation of indolizidine alkaloids in Securinega suffruticosa callus cultures. Plant Cell Tissue Organ Cult. 2015;123(1):39-45.[CrossRef]
70. Rout P, Naik N, Ngangkham U, Verma R, Katara JL, Singh O, et al. Doubled haploids generated through anther culture from an elite long duration rice hybrid, CRHR32:Method optimization and molecular characterization. Plant Biotechnol. 2016;33(3):177-86.[CrossRef]
71. Hooghvorst I, Ramos-Fuentes E, López-Cristoffanini C, Ortega M, Vidal R, Serrat X, et al. Antimitotic and hormone effects on green double haploid plant production through anther culture of mediterranean Japonica Rice. Plant Cell Tissue Organ Cult. 2018;134(2):205-15.[CrossRef]
72. Habibah NA, Lutfiah A, Liana A, Tunjung WA, Indrowati M, Pa'ee F. Callogenesis of Dayak onion (Eleutherine palmifolia) bulb in response of picloram, 2,4-D, and kinetin. Biosaintifika J Biol Biol Educ. 2023;15(2):270-80.[CrossRef]
73. Sharma M, Sharma M, Singh AK, Salgotra RK, Sharma M, Sharma BK,et al. In vitrodoubled haploid production of bacterial blight resistant plants from BC2F1 plants (Ranbir Basmati X Pau148) through anther culture. Int J Agric Syst. 2022;10(1):55.[CrossRef]
74. Muthu RK, Shanmugasundaram G, Karuppan T, Balakrishnan J, Ganesh D, Kathiresan S. Combined supplementation of 2, 4-D and kinetin elevates callus induction from cotyledon segments of different soybean cultivars [Glycine Max (L.) Merril]. Plant Arch. 2023;23(Suppl 2):25-31.[CrossRef]
75. Habibah NA, Yuniastuti A, Nugrahaning N, Safitri S, Puspitasari AD. Effects of low-dose kinetin, 2,4-D and monochromatic light conditions on flavonoid content in callus culture of Dioscorea esculenta. Trends Sci. 2024;21(2):7218.[CrossRef]
76. Wen X, Xu W, Zhang L, Shi X, Zhou J, Liu H, et al. Integrated metabolomic and transcriptomic analyses reveal the potential molecular mechanism underlying callus browning in Paeonia ostii. Plants. 2025;14(4):560.[CrossRef]
77. Handayani A, Hidayati S.Staurogyne elongata (nees) untze acanthaceae. Ethnobotany of the Mountain Regions of Southeast Asia. Ethnobotany of Mountain Regions. Springer. 2021;1015-20.[CrossRef]
78. Normasari R, Arumingtyas EL, Retnowati R, Widoretno W. The combination effect of auxin and cytokinin on callus induction of patchouli (Pogostemon cablinBenth.) leaf explants. Atlantis Press. 2023;551-7.[CrossRef]
79. Thaniarasu R, Kumar TS, Rao MV. Mass propagation of Plectranthus bourneaegamble through indirect organogenesis from leaf and internode explants. Physiol Mol Biol Plants. 2016;22(1):143-51.[CrossRef]
80. Sarropoulou V, Paloukopoulou C, Karioti A, Maloupa E, Grigoriadou K. Rosmarinic acid production fromOriganum dictamnus L. liquid cultures in vitro. Plants. 2023;12(2):299.[CrossRef]
81. Sayadi V, Mehrabi AA, Saidi M, Nourollahi K.In vitroculture and callus induction of chamomile (Matricaria chamomilla L.) under different concentrations of plant growth regulators. Int J Biosci. 2014;4:206-11.[CrossRef]
82. Padmanabhan P, Murch SJ, Sullivan JA, Saxena PK. Development of an efficient protocol for high frequency in vitro regeneration of a horticultural plant Primulina tamiana (B.L. Burtt) Mich. Möller A. Webber. Can J Plant Sci. 2014;94(7):1281-7.[CrossRef]
83. Mallón R, Rodríguez-Oubiña J, González ML. Shoot regeneration fromin vitro-derived leaf and root explants of Centaurea ultreiae. Plant Cell Tissue Organ Cult. 2011;106(3):523-30.[CrossRef]
84. Danial GH, Ibrahim DA. Organogenesis of tomato (Lycopersicon esculentum Mill.) Cv. sandra and rocky by hypocotyl, cotyledon and leaf explants. Arab J Agric Sci. 2020;3(5):31-54.[CrossRef]
85. Zeya S, Trivedi MP. Effect of rhustox on micropropagation of Scoparia dulcis L. leaf explants culture. Eur J Med Plants. 2019;29:1-14.[CrossRef]
86. Farzinebrahimi R, Taha RM, Rashid K, Yaacob JS. The effect of various media and hormones via suspension culture on secondary metabolic activities of (Cape Jasmine) Gardenia jasminoidesEllis. Sci World J. 2014;2014:1-7.[CrossRef]
87. Mokhtari A, Zarei M, Samsamzadeh B, Moradi K. Interactive effects of plant growth regulators and explants on direct shoot regeneration of Viola odorata. Biotechnologia. 2016;1:33-9.[CrossRef]
88. Daneshvar MH, Moallemi N, Zadeh NA. The effects of different media on shoot proliferation from the shoot tip of Aloe vera L. Jundishapur J Nat Pharm Prod. 2013;8(2):93-7.[CrossRef]
89. Singh M, Chaturvedi R. Improved clonal propagation of Spilanthes acmellaMurr. production of scopoletin. Plant Cell Tissue Organ Cult. 2010;103(2):243-53.[CrossRef]
90. Vahedi M, Kalantari S, Salami S. Effects of osmolytic agents on somatic embryogenesis of saffron. Notulae Sci Biol. 2015;7(1):57-61.[CrossRef]
91. Turgut BA, Bezirganoglu I. Callus induction and bioactive compounds production from various cultivars of Medicago sativa L. (Alfalfa). Igdir Üniv Bilim EnstitüsüDerg. 2023;13(3):1625-32.[CrossRef]
92. Junairiah J, Wulandari DA, Utami ES, Zuraidassanaaz NI. Callus induction and secondary metabolite profile from Elephantopus scaberL. J Trop Biodivers Biotechnol. 2021;6(1):59234.[CrossRef]
93. Gabbiesh AH, Ghabeish I, Ito H, Kleinwächter M, Selmar D. Plant regeneration through somatic embryogenesis from calli derived from leaf bases of Laurus nobilisL. (Lauraceae). Plant Tissue Cult Biotechnol. 2015;24(2):213-21.[CrossRef]
94. Nada MR, Wang L, ChunHua F, Xiangping X, Wenwen J, Li M, et al. Establishment and optimization of high efficiency embryogenic callus induction system in Dendrobium Candidum. Afr J Plant Sci. 2016;10(4):77-83.[CrossRef]
95. Zhang Z, Zhao H, Li W, Wu J, Zhou Z, Zhou F, et al. Genome-wide association study of callus induction variation to explore the callus formation mechanism of rice. J Integr Plant Biol. 2019;61(11):1134-50.[CrossRef]
96. Fehér A. A common molecular signature indicates the pre-meristematic state of plant calli. Int J Mol Sci. 2023;24(17):13122.[CrossRef]
97. Ikeuchi M, Favero DS, Sakamoto Y, Iwase A, Coleman D, Rymen B, et al. Molecular mechanisms of plant regeneration. Annu Rev Plant Biol. 2019;70(1):377-406.[CrossRef]
98. Uysal H, Sevindik E. Development of breeding lines by zigotic ovule culture in Nigella sativaL. Genetika. 2021;53(2):583-91.[CrossRef]
99. Valdiani A, Hansen OK, Nielsen UB, Johannsen VK, Shariat M, Georgiev MI, et al. Bioreactor-based advances in plant tissue and cell culture:Challenges and prospects. Crit Rev Biotechnol. 2018;39(1):20-34.[CrossRef]
100. Carp OE, Pinteala M, Arvinte A. Innovative non-enzymatic electrochemical quantification of cholesterol. Sensors. 2022;22(3):828.[CrossRef]
101. Ilyas AN, Rahmawati R, Widiastuti H. In vitro anticholesterol activity test of ethanol extract of gedi leaves (Abelmoschus manihot L. Medik). Wind Health J Kesehat. 2020;3:57-64. https://doi.org/10.33368/woh.v0i0.216 [CrossRef]
102. Parbuntari H, Prestica Y, Gunawan R, Nurman MN, Adella F. Preliminary phytochemical screening (qualitative analysis) of cacao leaves (Theobroma Cacao L.). Eksakta Berk Ilm Bid Mipa. 2018;19(2):40-5.[CrossRef]
103. Adu JK, Amengor CD, Kabiri NC, Orman E, Patamia SA, Okrah BK. Validation of a simple and robust liebermann-burchard colorimetric method for the assay of cholesterol in selected milk products in Ghana. Int J Food Sci. 2019;2019:9045938.[CrossRef]
104. Adamu A, Paul ED, Gimba CE, Ndukwe IG. Phytochemical and proximate analysis of Aspillia kotschyi(Sch. Bipex, Hochst) Oliv. Niger J Technol. 2018;36(4):1135.[CrossRef]
105. Tagousop CN, Tamokou J, Ekom SE, Ngnokam D, Voutquenne-Nazabadioko L. Antimicrobial activities of flavonoid glycosides from Graptophyllum grandulosum and their mechanism of antibacterial action. BMC Complement Altern Med. 2018;18(1):252.[CrossRef]
106. Garzón-Castaño SC, Lopera-Castrillón IA, Jiménez-González FJ, Siller-López F, Veloza LA, Arias JC. Nrf2-mediated antioxidant activity of the inner bark extracts obtained from Tabebuia rosea (Bertol) DC and Tabebuia chrysantha (JACQ) G. Nicholson. F1000research. 2019;7:1937.[CrossRef]
107. Nogueira FA, Nery PS, Morais-Costa F, Oliveira NJ, Martins ER, Duarte ER. Efficacy of aqueous extracts of Genipa americanaL. (Rubiaceae) in inhibiting larval development and eclosion of gastrointestinal nematodes of sheep. J Appl Anim Res. 2014;42(3):356-60.[CrossRef]
108. Olabinri BM, Adepoju EA, Zainab A, Ahmed A. Phytochemical profiling of phytoconstituents of grape, jatropha curcas and neem (Azadirachta indica) extracts. J Pharmacogn Phyther. 2014;6(2): 17-23.[CrossRef]
109. You Y, Choi N, Kim K. Ethanol extract of Ulmus pumilaroot bark inhibits clinically isolated antibiotic-resistant bacteria. Evid Based Complement Altern Med. 2013;2013:269874.[CrossRef]
110. Sam-Ang P, Phanumartwiwath A, Liana D, Sureram S, Hongmanee P, Kittakoop P. UHPLC-QQQ-MS and RP-HPLC detection of bioactive alizarin and scopoletin metabolites from Morinda citrifoliaroot extracts and their antitubercular, antibacterial, and antioxidant activities. Acs Omega. 2023;8(32):29615-24.[CrossRef]
111. Arora S, Lohiya GK, Moharir KS, Shah S, Yende SR. Identification of potential flavonoid inhibitors of the SARS-CoV-2 main protease 6YNQ:A molecular docking study. Digit Chinese Med. 2020;3(4):239-48.[CrossRef]
112. Najjaa H, Zouari S, Ammar E, Neffati M. Phytochemical screening and antibacterial properties of Allium roseum L., a wild edible species in North Africa. J Food Biochem. 2011;35(3):699-714.[CrossRef]
113. Li L, Liang X, Xu T, Xu F, Dong W. Rapid detection of six glucocorticoids added illegally to dietary supplements by combining TLC with spot-concentrated raman scattering. Molecules. 2018;23(7):1504.[CrossRef]
114. Poblocka-Olech L, Glód D, Jesionek A, Luczkiewicz M, Krauze-Baranowska M. Studies on the polyphenolic composition and the antioxidant properties of the leaves of poplar (Populusspp.) species and hybrids. Chem Biodivers. 2021;18(7):2100227.[CrossRef]
115. Cheng S, Huang M, Wu LC, Chou CC, Cheng C, Jhang S, et al. Building blocks for the development of an interface for high-throughput thin layer chromatography/ambient mass spectrometric analysis:A green methodology. Anal Chem. 2012;84(14):5864-8.[CrossRef]
116. Simlai A, Chatterjee K, Roy A. A comparative study on antioxidant potentials of some leafy vegetables consumed widely in India. J Food Biochem. 2013;38(3):365-73.[CrossRef]
117. Zhang F, Ren H, Wang Y, Wang R, Sheng J. Bioassay-guided isolation and identification of a acetylcholinesterase inhibitors from Calophyllum polyanthum. Adv Mater Res. 2013;634-638:1225-8.[CrossRef]
118. Abreu IN, Ahnlund M, Möritz T, Albrectsen BR. UHPLC-ESI/TOFMS determination of salicylate-like phenolic gycosides in Populus tremula leaves. J Chem Ecol. 2011;37(8):857-70.[CrossRef]
119. Ceglowski M, Smoluch M, Reszke E, Silberring J, Schroeder G. Flowing atmospheric pressure afterglow combined with laser ablation for direct analysis of compounds separated by thin-layer chromatography. Anal Bioanal Chem. 2015;408(3):815-23.[CrossRef]
120. Balogun OS, Oladosu IA, Liu Z. Isolation of 2, 5-dithia-3, 6-diazabicyclo [2.2.1] heptane and GC-MS analysis of silylated extract from Tragia benthamii. IFE J Sci. 2020;22(2):75-80.[CrossRef]
121. Mohapatra M, Basak UC. UHPLC assessment of embelin in specialized mangrove plant Aegiceras corniculatum (L) blanco. Bull Natl Res Cent. 2022;46(1):215.[CrossRef]
122. Stankevicius M, Akuneca I, Jakobsone I, Maruška A. Comparative analysis of radical scavenging and antioxidant activity of phenolic compounds present in everyday use spice plants by means of spectrophotometric and chromatographic methods. J Sep Sci. 2011;34(11):1261-7.[CrossRef]
123. Meletiadis J, Mortensen KL, Verweij PE, Mouton JW, Arendrup MC. Spectrophotometric reading of EUCAST antifungal susceptibility testing of Aspergillus fumigatus. Clin Microbiol Infect. 2017;23(2):98-103.[CrossRef]
124. Gharagheizi F, Eslamimanesh A, Mohammadi AH, Richon D. Group contribution-based method for determination of solubility parameter of nonelectrolyte organic compounds. Ind Eng Chem Res. 2011;50(17):10344-9.[CrossRef]
125. Michel AP, Morrison AE, Preston V, Marx CT, Colson BC, White HK. Rapid identification of marine plastic debris via spectroscopic techniques and machine learning classifiers. Environ Sci Technol. 2020;54(17):10630-7.[CrossRef]
126. Albrektiene R, Rimeika M, GrazénienéR. Organic Fractions and Metal-Organic Complexes in the Groundwater. Vilnius Gediminas Technical University Press Technika. 2014;1-7.
127. Anim-Eduful B, Adu-Gyamfi K. Functional groups detection:Do chemistry teachers demonstrate conceptual difficulties in teaching? Glob J Hum Soc Sci. 2021;21:47-60.[CrossRef]
128. Arisawa M. Transition-metal-catalyzed synthesis of organophosphorus compounds involving P-P bond cleavage. Synthesis (Stuttg). 2020;52(19):2795-806.[CrossRef]
129. Megawati ER, Bangun H, Putra IB, Rusda M, Syahrizal D, Jusuf NK, et al. Phytochemical analysis by FTIR of Zanthoxylum acanthopodium, DC fruit ethanol extract, N-hexan, ethyl acetate and water fraction. Med Arch. 2023;77(3):183.[CrossRef]
130. Liang M, Yu C, Dai S, Cheng H, Li W, Lai F, et al. Reactivity and kinetics of 1,3-butadiene under ultraviolet irradiation at 254 nm. BMC Chem. 2022;16(1):4.[CrossRef]
131. Hamka A, Fatimawali F, Datu OS, Tallei TE. Antibacterial activity of betel (Piper betle L.) against Pseudomonas aeruginosa. Acta Biochim Indones. 2021;4(1):10.[CrossRef]
132. Boulet J, Ducasse MA, Cheynier V. Ultraviolet spectroscopy study of phenolic substances and other major compounds in red wines:Relationship between astringency and the concentration of phenolic substances. Aust J Grape Wine Res. 2017;23(2):193-9.[CrossRef]
133. Jelic D, Fazlagic S, Antunovic V, Bubic-Pajic N, Racic A, Dermanovic M. Analysis of ambroxol hydrochloride in flavamed®tablets by means of spectroscopic absorption methods. Qual Life (Banja Luka) Apeiron. 2016;13(1-2):24-28.
134. Ghodsi F, Habibi-Khorassani SM, Shahraki M. Kinetic spectrophotometric method for the 1,4-diionic organophosphorus formation in the presence of meldrum's acid:Stopped-flow approach. Molecules. 2016;21(11):1514.[CrossRef]
135. Mutiarahma S, Putra VG, Chaniago W, Carrera C, Anggrahini S, Palma M, et al. UV-Vis spectrophotometry and UPLC-PDA combined with multivariate calibration for Kappaphycus alvarezii (Doty) doty ex silva standardization based on phenolic compounds. Sci Pharm. 2021;89(4):47.[CrossRef]
136. Paulino FW, Julião MS, Paiva AS, Rodrigues LG, Moreira FL, Teixeira AM, et al. Qualitative identification of synthesized isoniazid derivatives by atemporal technique:UV-visible spectrophotometry. Concilium. 2024;24(5):139-52.[CrossRef]
137. Riswahyuli Y, Rohman A, Setyabudi FM, Raharjo S. Indonesian wild honey authenticity analysis using attenuated total reflectance-fourier transform infrared (ATR-FTIR) spectroscopy combined with multivariate statistical techniques. Heliyon. 2020;6(4):03662.[CrossRef]
138. Pratiwi R, Rahmawaty A, Hasanah AN. Simple analytical method on the determination of dexamethasone in herbal medicine. J Spectrosc. 2022;2022:1-5.[CrossRef]
139. Dulanlebit YH, Hattu N, Sangadji RP, Mahulette R, Kelmanutu N, Asari S. Performance of caffeine content analysis in Robusta Sp and Theobroma Cacao L using iodometry, UV-vis spectrophotometry and high performance liquid chromatography. J Pure Appl Chem Res. 2020;9(3):212-21.[CrossRef]