
1. INTRODUCTION
Herpes simplex virus type 1 (HSV-1) is classified under the Herpesviridae family, Alphaherpesvirinae subfamily, and Simplexvirus genus [1]. HSV-1 is highly contagious and still endemic in various parts of the world [2]. Primary HSV-1 infection mainly affects the oral mucosal epithelium and is clinically manifested as herpes labialis or herpes gingivostomatitis [3]. The initial infection can become a latency stage, permanently affecting human hosts’ peripheral nervous system [4]. Commercially available antiherpetic drugs have a narrow mechanism of action. For instance, nucleoside and nucleotide analogs only target the viral DNA replication stages; therefore, a complete prevention or cure is not achieved [5]. Hence, it is vital to conduct research on natural products to discover their potential as therapeutic agents against HSV-1 infection. Natural products have distinct structure and chemical compounds which could be an ideal source of new antiherpetic agents [6]. In recent times, bioactive compounds derived from natural products such as plants, microorganisms, marine organisms, fungi, and animal are being explored with great interest as potential antiherpetic agents [7]. Plant-derived bioactive compounds have been shown to be able to elicit potent antiviral effects against HSV at multiple phases of viral replication including latency stage [8-11]. Besides that, natural botanical products are less cytotoxic toward mammalian cells, produce less undesired secondary effects, and are cheap in terms of production [12].
Kyllinga nemoralis, also known as whitehead spike sedge, poverty grass, white globose spike, or white Kyllinga is a monocotyledonous flowering graminoid plant generally similar to grasses, herbs, or rushes [13,14]. The medicinal values of K. nemoralis are often attributed to bioactive compounds in the plant. Various plant morphological sections and extracts contain secondary bioactive compounds and metabolites. The chemical compounds are usually identified through a phytochemical screening. A whole plant can be converted into several decoctions through a solvent mixing process to produce various aqueous, chloroform, ethanol, methanol, and hexane extracts [15]. The aerial divisions of K. nemoralis aqueous extract contain chemicals such as carbohydrates, phenolic compounds, protein, amino acids, flavonoids, and glycosides [16]. The whole K. nemoralis plant aqueous extract possesses potent secondary metabolites such as carbohydrates, saponin, anthocyanin, and coumarins [17]. A combination of roots and leaves sections of K. nemoralis has active chemical substances such as phenolic compounds, terpenoids, saponin, glycosides, triterpenoids, flavonoids, and essential oils [18]. The distillate of K. nemoralis ethanolic rhizome roots is positive for volatile oil, carbohydrates, and alkaloids [19].
Polyphenols are secondary metabolites derived from plants, often distinguished by more than 1 hydroxyl (-OH) group on complex aromatic ring or benzene ring formation. Phenolic acids, tannins, and flavonoids are polyphenols that exhibit antiviral, antibacterial, hepatoprotective, and anti-tumor activities [20]. Phenolic compounds alter the conformation and function of HSV-1 envelope proteins, which further prevent viral fusion and penetration [12]. Furthermore, the phenolic compound also inhibits the expression of the viral gene [7]. Flavonoids are diphenyl propane-structured secondary metabolites found in plants. Flavonoids have been known to prevent microbes from invading and infecting hosts. Therefore, plants containing flavonoids are widely used as traditional therapy for treating various communicable diseases caused by microbes [21]. Research in the past has shown evidence of flavonoids exhibiting antiviral activity. Flavonoid compounds interrupt HSV-1 viral entry into host cells by directly inhibiting viral particles [22]. It is a vital discovery because the current antiviral drug, acyclovir (ACV), only inhibits viral DNA replication [23].
The antiviral activity of K. nemoralis extract against HSV-1 has been scarcely explored. However, other researchers have investigated this plant’s broad curative properties. K. nemoralis ethanol extract exhibited intense larvicidal activity against three species of larvae that transmit dengue, malaria, and Japanese encephalitis; the exposure of larvae to plant extract resulted in pathological damages such as cell degradation and lysis [17]. Methanol extract of K. nemoralis exhibits antibacterial properties; it showed the highest inhibition against Streptococcus pneumoniae growth on the culture medium [24]. Another study reported the anthelminthic activity of the plant’s root ethanol extract. These scientific research show that K. nemoralis can destroy several types of organisms. However, the antiviral effect of K. nemoralis extract has not been investigated. Therefore, in this present study, we explored the possible action of K. nemoralis stem with leaves aqueous extract against HSV-1 replication in vitro.
2. MATERIALS AND METHODS
2.1. Plant Material
The plants were collected in Kuala Nerus, Terengganu, Malaysia. The stem with leaf of K. nemoralis plants (500 g) was cleaned with water and then dried at 27°C or 80 F until it dried thoroughly. The plant parts were then oven-dried at 65–70°C for 48 h. After that, the plant parts were cut into small fragments and ground using electric food processor (Kenwood Ltd., Havant, United Kingdom). The coarse powder was preserved in air-tight container.
2.2. Extraction Method
For preparing aqueous extract of K. nemoralis, 100 g powder of the stem with leaf was mixed into deionized water with a ratio of powder to water of 10:1 according to a published method with some modifications [25]. The mixture was boiled for 2 h. The mixture was then allowed to cool down to room temperature before being filtered to collect the extract. After that, the extract was frozen at −20°C for 7 h. Then, freeze-dry technique was performed to lyophilize the extract.
2.3. Cells and Virus
The Vero cells were cultivated in Dulbecco’s Modified Eagle Medium (DMEM; Sigma, USA) supplemented with 5% fetal bovine serum (FBS; JR Scientific, USA) and penicillin/streptomycin (100 IU) (Gibco-Life Technologies, USA). Clinical strain of HSV-1 propagation was done in Vero cells cultured in DMEM containing 5% FBS and incubated at 37°C, 5% CO2 until cytopathic was observed. After that, plaque assay has been used in determining the titer of virus stock.
2.4. Cytotoxicity Test
Colorimetric analysis in 96-well plate utilizing 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was carried out to determine the cytotoxicity level of K. nemoralis stem with leaves aqueous extract as previously described [26]. The 50% cytotoxicity concentration (CC50) was characterized as the sample concentration that is able to reduce 50% of cell viability compared to the untreated cells. Briefly, monolayer of Vero cells was seeded in 96-well microplates (2.0 × 104 cells/well) and exposed with several concentrations of extract (31–1000 μg/mL) in triplicates. MTT was added into each well after 48 h of incubation. The plate was further incubated for at least 2 h and dimethyl sulfoxide was added to each well to dissolve the formazan crystals. Optical density of each well was measured using spectrophotometer at 540 nm and the dose-response curve was plotted using GraphPad Prism 5.0 (GraphPad Software Inc., San Diego, CA).
2.5. Post-Treatment of HSV-1-Infected Cells with Plant Extract
Antiviral activity of K. nemoralis stem with leaves aqueous extract against intracellular replication of HSV-1 was carried out by exposing the Vero cells (2.0 × 106 cells/well) with extract after adsorption with 50 plaque-forming unit (PFU) of virus for 2 h at 37°C to permit virus adsorption. The different concentrations of extract (1–31 μg/mL) were mixed with DMEM + methylcellulose immediately after the adsorption period and added into all test wells. The cells were then incubated for 48 h. After incubation, the cells were stained using crystal violet and plaques were counted according to published protocol [27].
2.6. Pre-Treatment of Plant Extract on HSV-1-Infected Cells
The assay was conducted as reported by a previously published method [28]. Initially, Vero cells (2.5× 106 cells/well) were seeded into 24-well plate and incubated for 24 h. Cells were pre-treated with different concentrations of extract (1–31 μg/mL) for 24 h before being withdrawn and infected with 300 μL of virus at 50 PFU. The cells were incubated at 37°C in 5% CO2 for 1 h 30 min to ensure the adsorption of virus into cells. After incubation, the cells were stained using crystal violet and plaques were counted. Selective index, SI (CC50/effective concentration [EC50] ratio, where EC50 is 50% EC50), was used to evaluate K. nemoralis effectiveness as a prospective antiviral agent.
2.7. Direct Virucidal Assay
Extracellular anti-HSV-1 activity of K. nemoralis stem with leaves aqueous extract was determined by incubating 50 PFU HSV-1 suspension with equal volume of the various concentrations of extract (1–31 μg/mL) for 30 min at 37°C. Then, viral suspension was infected to the Vero cells for 1 h at 37°C. The cells were washed with phosphate-buffered saline to remove the unabsorbed viruses. Then, the cells were incubated at 37°C for 48 h. Antiviral activity was evaluated by viral plaques reduction assay [29].
3. RESULTS AND DISCUSSION
3.1. Cytotoxicity of K. nemoralis Stem with Leaves Aqueous Extract
Cytotoxicity assay was first done ahead of antiviral screening to determine K. nemoralis stem with leaves aqueous extract concentration which is cytotoxic to 50% of cell population (CC50). From this CC50 value, lower concentrations which were least toxic or non-toxic to the cells were used in antiviral screening. To evaluate the non-toxic dose, Vero cells were treated with 2-fold serially diluted extract at concentrations ranging from 31 to 1000 μg/mL. In this assay, the CC50 value of K. nemoralis stem with leaves aqueous extract was determined at 56 μg/mL [Figure 1]. No signs of toxicity were remarked in Vero cells when the extract was added at concentrations ranging from 2 to 31 μg/mL. The cells exhibited a normal monolayer similar to the healthy control cells. No loss of monolayer, granulation, rounding, or dwindling of cells was seen. K. nemoralis stem with leaves aqueous extract CC50 was considered to be not in the range of cytotoxic compound since the value of CC50 of the extract was determined to be higher than minimum value for compound with active cytotoxicity. Any CC50 or IC50 (50% inhibitory concentration) value of a compound that was <4 μg/mL was considered to have active cytotoxic effect [30].
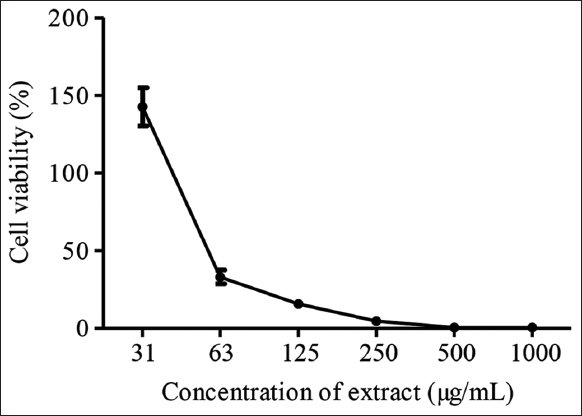 | Figure 1: Cytotoxicity analysis of Kyllinga nemoralis stem with leaves aqueous extract. [Click here to view] |
3.2. Antiviral Assays with K. nemoralis Against HSV-1
The extract was further evaluated for their ability to inhibit replication after infection of the cells with the virus (post-treatment), the effect of the extract as prophylactic agent in protecting the cell from virus adsorption and penetration (pre-treatment), and the ability of the extract to act directly against virus virion particle (virucidal). Plaque reduction assays were done to screen for anti-HSV-1 activity using K. nemoralis stem with leaves aqueous extract with different concentrations. Extract was added at the different stages of viral replication cycle: (i) treatment for 48 h after 2 h of post-adsorption, (ii) pre-treatment for 24 h after infection for studying, the prophylactic activity, and (iii) direct treatment to cell with virus suspension to evaluate direct virucidal effect.
Potency of some extracts can be assessed using SI. Antiviral agent can be considered to have high potential to be developed as antiviral drug if their SI value is more than 10 [31]. Comparison of SI for the extract between post-treatment, pre-treatment, and virucidal assay is shown in Table 1. SI for post-treatment for the extract showed high values compared to those obtained from pre-treatment and virucidal assay. Therefore, the post-treatment was more efficient in suppressing viral replication compared to pre-treatment and virucidal. Results from our study suggest that K. nemoralis stem with leaves aqueous extract shows strong in vitro antiviral activity against all stages of the HSV-1 replication cycle such as intracellular virus replication (SI = 14.0). On the other hand, K. nemoralis stem with leaves aqueous extract showed mild direct virucidal activity (SI = 8.62), while its prophylactic effect against HSV-1 replication was weak (SI = 4.31).
Table 1: CC50, EC50, and SI values of Kyllinga nemoralis stem with leaves aqueous extract in post-treatment, pre-treatment, and virucidal assay.
| Assay | CC50 (µg/mL) | EC50 (µg/mL) | SI (CC50/EC50) |
|---|---|---|---|
| Post-treatment | 56 | 4 | 14.0 |
| Pre-treatment | 56 | 13 | 4.31 |
| Virucidal | 56 | 6.5 | 8.62 |
Post-treatment assay aims to investigate the intracellular HSV-1 inhibitory activity of K. nemoralis aqueous extract [32]. The highest plaque inhibition of 74% was observed in the post-treatment assay when HSV-1-infected Vero cells were treated with 31 μg/mL of K. nemoralis stem with leaves aqueous extract [Figure 2]. The effectiveness of the plant extract during post-treatment assay was attributed to its ability to inhibit intracellular virus replication upon entry into the cell. Viral gene replication of HSV-1 is a cascade event that involves the production of immediate-early, early, and late genes in sequential order [33]. In addition to the finding that K. nemoralis stem with leaves aqueous extracts block intracellular replication on viral entry into cells, the effectiveness of an antiviral agent during post-treatment step may indicate its ability to interfere with viral replication, DNA translation, assembly, and egress of progeny virions from cells [34]. Flavonoid subclasses, especially baicalin, fisetin, and quercetin, are classified as flavonols possessing anti-HSV-1 properties [35]. Flavonols such as quercetin have been found to be actively existing in K. nemoralis plant extract [36]. This was further supported by the observation in a study in which quercetin compounds vigorously hampered HSV-1 infectivity principally during post-treatment step [37]. Hence, the anti-herpetic activity of K. nemoralis aqueous extract could be related to the presence of flavonol sub-compounds such as quercetin. Besides that, there are mounting evidence of plant-derived polyphenols manifesting anti-herpetic efficacy through synergistic interaction during post-treatment stage [38].
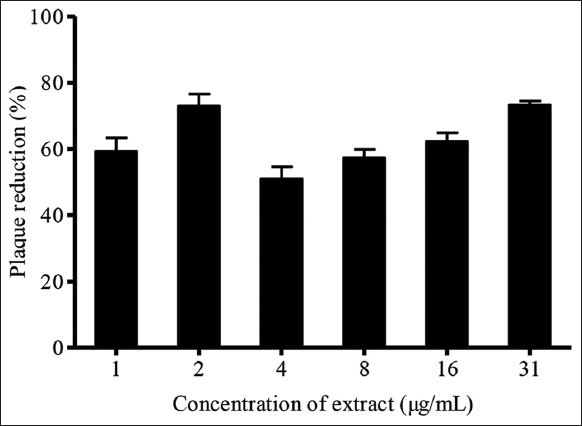 | Figure 2: Effects of post-treatment of Kyllinga nemoralis stems with leaves aqueous extract on HSV-1 plaque reduction. [Click here to view] |
Pre-treatment assay evaluated the potential of K. nemoralis stem with leaves and roots aqueous extract to interact with host cells to stop viral adsorption into cells. The cells were pre-incubated with plant extract for 24 h to ensure sufficient absorption of respective decoction into cells. In this assay, the highest plaque inhibition of 65% was examined when Vero cells were pre-treated with 1 μg/mL of K. nemoralis stem with leaves aqueous extract [Figure 3]. The EC50 of K. nemoralis stem with leaves aqueous extract in pre-treatment was higher, 13 μg/mL, than post-treatment, 4 μg/mL. The contrast in findings suggests that stem with leaves aqueous extract has impotent interactions with Vero cells. This ineffectual interactivity may have caused the weak prophylactic activity of stem with leaves extract on host cells. Furthermore, the higher EC50 corresponding to a lower SI of 4.31 shows that the extract is relatively toxic to cells when administered as pre-treatment. Therefore, this extract exhibits a weak direct protective effect on Vero cells. The 24 h incubation of cells extract might be inadequate to exert any protective effects on cells. However, this contradicts the above finding in which the extract is toxic to cells when given as pre-treatment, and more prolonged incubation would only contribute to more toxicity. Antiviral activity exhibited by extracts could be due to the inhibition of host cell glycoprotein receptors or prevention of HSV-1 virions binding to host cells. A study has shown that mutated cultured cells lacking glycosaminoglycans such as heparan sulfate reduced the binding efficiency of the virus to cells [39]. Therefore, the destruction of cellular receptors vital for membrane fusion could be the most plausible mechanism exerted by this extract.
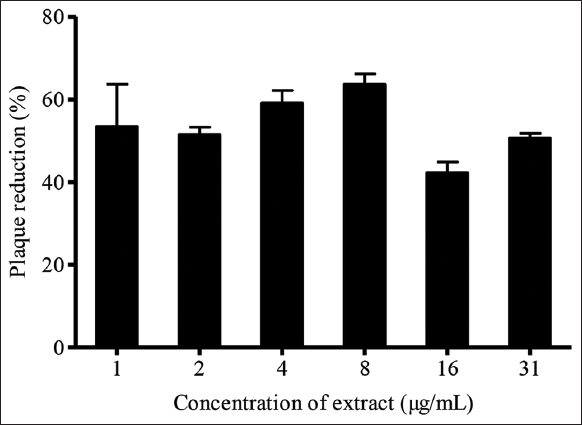 | Figure 3: Effects of pre-treatment of Kyllinga nemoralis stems with leaves aqueous extract on HSV-1 plaque reduction. [Click here to view] |
Virucidal assay determined the direct virucidal effect of plant extract on HSV-1. In this assay, K. nemoralis stem with leaves aqueous extract inhibited 66% of virus plaque at 31 μg/mL. As for EC50, K. nemoralis stem with leaves aqueous extract exhibited a lower value, 6.5 μg/mL [Figure 4]. This indicates that the extract can produce antiviral effects at a lower concentration within the safe cytotoxic span. The direct virucidal activity of K. nemoralis stem with leaves aqueous extract shows its ability to interact with and inhibit HSV-1 virions outside Vero cells [40]. An initial deactivation of HSV-1 particle could avert the establishment of primary viral infection [34]. In this study, the direct disruption of virus particles could be a target to suppress initial stages of infection such as virus adsorption, membrane fusion, and entry into host cells [41]. Apart from the whole virus destruction, K. nemoralis stem with leaves aqueous extract may have interfered with the viral envelope to conceal viral glycoproteins necessary for HSV-1 attachment or entry into cells. The virucidal effectiveness of natural compound during extract-virus interaction exhibits its potential to prevent virus attachment to host cells. At this stage of investigation, the specific viral glycoprotein/glycoproteins blocked by plant extract to mediate the anti-herpetic mechanism is yet to be identified. Nevertheless, the outcome of the present study confirms the virucidal potential of K. nemoralis stem with leaves aqueous extract which is most likely due to the suppression of HSV-1 surface glycoproteins and consequent inactivation of viral particles [42].
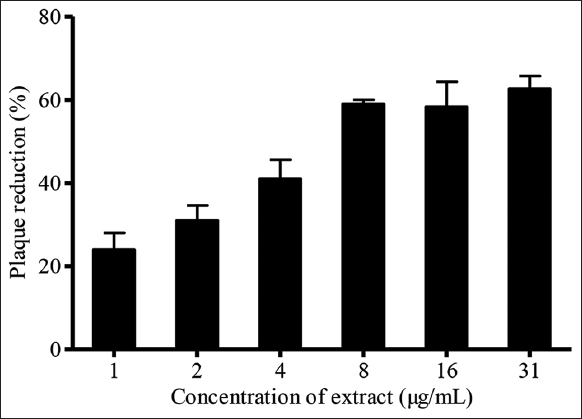 | Figure 4: Effects of virucidal assay of Kyllinga nemoralis stems with leaves aqueous extract on HSV-1 plaque reduction. [Click here to view] |
4. CONCLUSION
This study presented the potential of K. nemoralis aqueous extract as a new anti-HSV-1 agent. A more comprehensive study will be needed to explain the mechanisms involved and to determine whether the antiviral activity of K. nemoralis toward HSV-1 could be more potent when purified active compounds are used.
5. ACKNOWLEDGMENTS
The authors would also like to thank the management of Universiti Sultan Zainal Abidin, Terengganu, Malaysia, for facilitating and funding this study with research grant number UniSZA/2020/LABMAT/06.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. DATA AVAILABILITY
The data presented in this study are not available on request from the corresponding author.
10. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Dai X, Zhou ZH, Genetics M, Angeles L. Structure of the herpes simplex virus 1 capsid with associated tegument protein complexes. Science 2018;360:eaao7298. [CrossRef]
2. Nasrallah GK, Dargham SR, Abu-Raddad LJ. Negative epidemiological association between HSV-1 and HSV-2 infections. Heliyon 2019;5:e02549. [CrossRef]
3. Mody PH, Pathak S, Hanson LK, Spencer JV. Herpes simplex virus:A versatile tool for insights into evolution, gene delivery, and tumor immunotherapy. Virology (Auckl) 2020;11:1178122X20913274. [CrossRef]
4. Marcocci ME, Napoletani G, Protto V, Kolesova O, Piacentini R, Li Puma DD,
5. Ghosh A, Piazza P, Yee MB, Kumar S, Milosevic J, Bloom DC,
6. Treml J, GazdováM, Šmejkal K, ŠudomováM, Kubatka P, Hassan ST. Natural products-derived chemicals:Breaking barriers to novel anti-HSV drug development. Viruses 2020;12:154. [CrossRef]
7. Hassan ST, Masarcikova R, Berchova K. Bioactive natural products with anti-herpes simplex virus properties. J Pharm Pharmacol 2015;67:1325-36. [CrossRef]
8. Wahab NZ, Bunawan H, Ibrahim N. Cytotoxicity and antiviral activity of methanol extract from
9. Nazlina I, Norha S, Zarina NA, Ahmad IB. Cytotoxicity and antiviral activity of
10. Ripim NS, Fazil N, Ibrahim SN, Bahtiar AA, Wai YC, Ibrahim N,
11. Ibrahim N, Shahar S, Wahab NZ, Nor NS. Effect of styrylpyrone derivative (SPD) and SPD/foscarnet combination towards virus infected cell. AIP Conf Proc 2019;2111:040002. [CrossRef]
12. Gavanji S, Sayedipour SS, Larki B, Bakhtari A. Antiviral activity of some plant oils against herpes simplex virus Type 1 in vero cell culture. J Acute Med 2015;5:62-8. [CrossRef]
13. Mathew TK, Benny PJ. Flowers with antibacterial property-a survey in Meenachil Taluk. J Pharm Chem Biol Sci 2016;4:252-61.
14. Odedeji ET, Adedeji O. Comparative foliar epidermal studies of twelve species in the family
15. Abubakar AR, Haque M. Preparation of medicinal plants:Basic extraction and fractionation procedures for experimental purposes. J Pharm Bioallied Sci 2020;12:1-10. [CrossRef]
16. Rajagopal PL, Kumar PN, Sreejith KR, Premaletha K. Phytochemical and antioxidant screening of the aerial parts of
17. Babu M, Kaleena PK, Janaki A, Velu K, Ravi S. Larvicidal activity and histopathological alterations effected by
18. Picardal JP, Abella KJ, Abella R, Villamor MJ, Picardal MT. Toxicity evaluation of white spike head (
19. Majumder P. Investigation of anthelmintic activity of an ignored plant “
20. Kamboj A, Saluja AK, Kumar M, Atri P. Antiviral activity of plant polyphenols. J Pharm Res 2012;5:2402-12.
21. Abid NB, Rouis Z, Lassoued MA, Sfar S, Aouni M. Assessment of the cytotoxic effect and
22. Hung PY, Ho BC, Lee SY, Chang SY, Kao CL, Lee SS,
23. Li W, Wang XH, Luo Z, Liu LF, Yan C, Yan CY,
24. Sindhu T, Rajamanikandan S, Srinivasan P.
25. Wahab NZ, Ibrahim N, Kamarudin MK, Lananan F, Juahir H, Ghazali A,
26. Wahab NZ, Azizul A, Muhamad N, Ibrahim N. Cytotoxic and antiviral activities of
27. Moses M, Nor NS, Ibrahim N.
28. Wahab NZ, Azizul A, Badya N, Ibrahim N. Antiviral activity of an extract from leaves of the tropical plant
29. Wahab NZ, Azizul A, Ibrahim N. Phytochemistry, cytotoxicity and antiviral activity of
30. Lee CC, Houghton P. Cytotoxicity of plants from Malaysia and Thailand used traditionally to treat cancer. J Ethnopharmacol 2005;100:237-43. [CrossRef]
31. Dargan DJ. Methods in Molecular Medicine. Totowa, New Jersey:Humana Press;1998.
32. Zeng Z, Zhang R, Hong W, Cheng Y, Wang H, Lang Y,
33. Harkness JM, Kader M, DeLuca NA. Transcription of the herpes simplex virus 1 genome during productive and quiescent infection of neuronal and nonneuronal cells. J Virol 2014;88:6847-61. [CrossRef]
34. Aoki-Utsubo C, Chen M, Hotta H. Time-of-addition and temperature-shift assays to determine particular step(s) in the viral life cycle that is blocked by antiviral substance(s). Bio Protoc 2018;8:e2830. [CrossRef]
35. Lyu SY, Rhim JY, Park WB. Antiherpetic activities of flavonoids against herpes simplex virus Type 1 (HSV-1) and Type 2 (HSV-2)
36. Thangaraj S, Sundarraj R, Srinivasan P.
37. Lee S, Lee HH, Shin YS, Kang H, Cho H. The anti-HSV-1 effect of quercetin is dependent on the suppression of TLR-3 in raw 264.7 cells. Arch Pharm Res 2017;40:623-30. [CrossRef]
38. Musarra-Pizzo M, Pennisi R, Ben-Amor I, Smeriglio A, Mandalari G, Sciortino MT.
39. Shieh MT, WuDunn D, Montgomery RI, Esko JD, Spear PG. Cell surface receptors for herpes simplex virus are heparan sulfate proteoglycans. J Cell Biol 1992;116:1273-81. [CrossRef]
40. Jaime MF, Redko F, Muschietti LV, Campos RH, Martino VS, Cavallaro LV.
41. Li W, Xu C, Hao C, Zhang Y, Wang Z, Wang S,
42. Hayashi K, Hayashi T, Otsuka H, Takeda Y. Antiviral activity of 5,6,7-trimethoxyflavone and its potentiation of the antiherpes activity of acyclovir. J Antimicrob Chemother 1997;39:821-4. https://doi.org/10.1093/jac/39.6.821 PMid:9222055