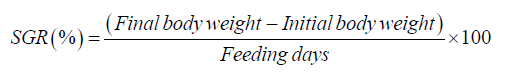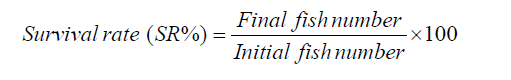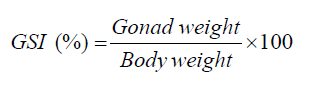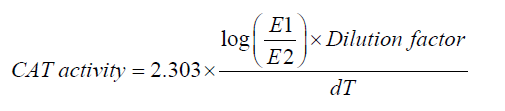Bisphenol A (BPA) is a well-known endocrine disruptor that negatively impacts reproductive physiology, barrier function, and overall metabolic health in aquatic organisms. Melatonin, a hormone secreted by the pineal gland, possesses antioxidant and chronobiotic properties and has been proposed as a potential protective agent against BPA-induced toxicity. However, evidence supporting efficacy in teleost models remains limited. In this study, zebrafish (Danio rerio) were exposed to BPA and melatonin for 21 days to assess alterations in circadian rhythm-associated genes (Clock1a, Cry3a, Per1b, Mt1), tight junction proteins (Claudin, Occludin), inflammatory cytokines (Interleukin-6, Tumor Necrosis Factor-alpha), antioxidant enzymes, and histopathological changes in the gut and ovary. BPA exposure resulted in significant downregulation of circadian genes and tight junction markers, accompanied by elevated levels of inflammatory mediators and oxidative stress. In contrast, melatonin administration alone enhanced gene expression, antioxidant capacity, and tissue integrity. Moreover, co-treatment with melatonin effectively mitigated the adverse effects of BPA, restoring gene expression and reducing inflammation and oxidative damage. These findings highlight the role of melatonin in counteracting BPA-induced toxicity and support its potential application as a therapeutic agent in aquatic toxicology.
Patel Y, Das B, Ramachandran AV, Gajaria T, Baxi D. Melatonin attenuates bisphenol A-induced circadian disruption, gut barrier damage, and ovarian dysfunction in zebrafish. J Appl Biol Biotech 2026;14(2):145-156. http://doi.org/10.7324/JABB.2026.282178
Bisphenol A (BPA) is extensively used in polycarbonate plastic and epoxy resin production [1,2]. Prevalent usage of BPA in consumer materials has resulted in widespread environmental contamination and consequent chronic exposure to humans and animals. BPA is an endocrine-disrupting compound (EDC) with the capability of interfering with hormonal signaling pathways and impairing reproductive function, homeostatic disruption of metabolism, and developmental abnormalities [3]. Reproductive toxicity, oxidative stress, inflammatory reactions, and gut microbiota dysbiosis have been recognized in aquatic animals, including zebrafish (Danio rerio), due to BPA exposure [4,5].
Melatonin, which is an indoleamine produced mainly by the pineal gland, has multiple physiological roles, such as circadian rhythm regulation, antioxidant defense, and modulation of the immune response [6,7]. As an amphiphilic molecule, melatonin easily permeates cell membranes and the blood–brain barrier and thus easily scavenges reactive nitrogen and oxygen species and upregulates endogenous antioxidant enzymes [8]. When melatonin was recently shown to reduce inflammatory cascades and protect mitochondria under toxicological stress, a potential EDC-protecting agent was identified [9].
Despite abundant documentation of BPA’s toxicity and the protectant effects of melatonin, there are very few studies exploring the role of melatonin among aquatic vertebrates. Zebrafish is a favorable model system for endocrine disruption research due to the conserved nature of its hormonal pathways, translucent embryonic development, and the convenience of exposure assessment [10]. In this study, the focus is on exploring the capability of melatonin in mitigating the impact of BPA exposure to restore zebrafish ovarian-circadian homeostasis, alongside maintaining gut integrity. Melatonin supplementation has been efficacious in offsetting BPA-caused endocrine, inflammatory, and gut microbiota disruptions.
2. MATERIALS AND METHODS
2.1. Experimental Design and Husbandry of Animals
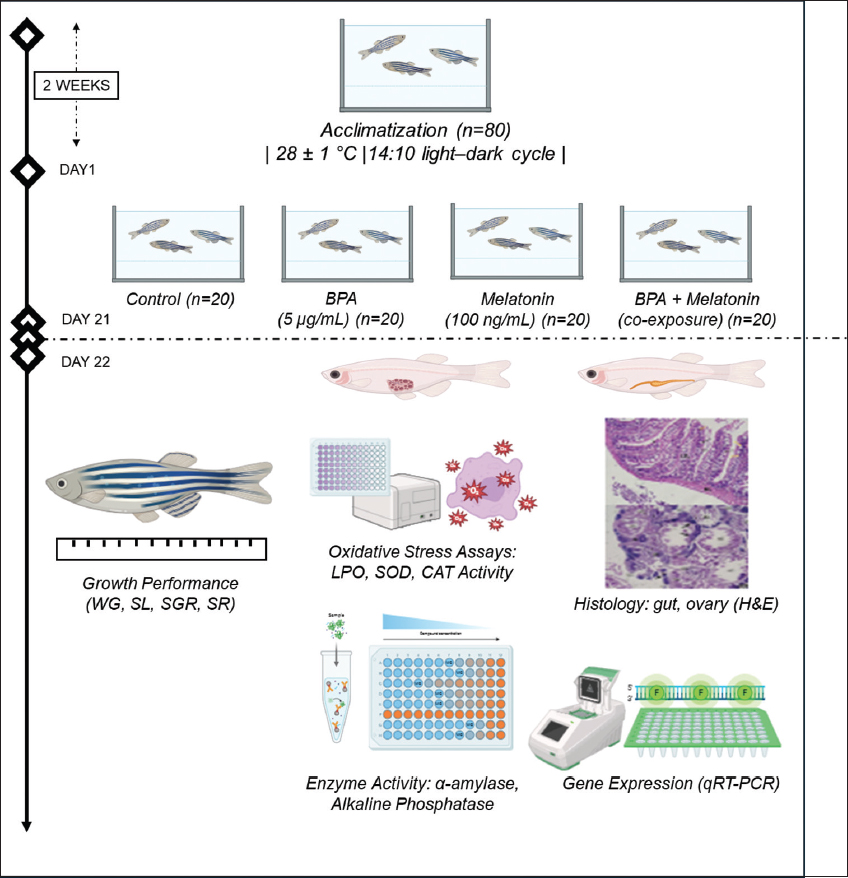
Adult wild-type female zebrafish (n = 20 in a 20 L tank) were procured from an accredited breeder and acclimatized under regular lab conditions for 2 weeks before experimentation commenced. Fish were maintained in aerated freshwater at 28 ± 1°C under a 14:10 h light–dark cycle and with constant filtration and water quality checks. They were fed a commercial dried worm diet twice a day, as shown in Figure 1. All experimental procedures were conducted in accordance with the guidelines of the Institutional Ethics Committee and followed all the guidelines of ARRIVE 2.0. The experimental set was carried out in triplicate.
Following acclimatization, fish were randomized using a computer-generated sequence (Excel RAND), blocked by body weight (BW) (±10%), and distributed across the three replicate tanks per group to avoid tank effects, then allocated to one of four experimental groups (n = 20 per group):
Group 1 (Control): Maintained in clean system water without any additives
Group 2 (BPA): Exposed to BPA at a concentration of 5 μg/mL
Group 3 (Melatonin): Treated with melatonin at a concentration of 100 ng/mL
Group 4 (BPA + Melatonin): Co-exposed to BPA (5 μg/mL) and melatonin (100 ng/mL) simultaneously.
2.2. Chemical Preparation, Exposure Protocol, and Sample Collection
BPA (Sigma-Aldrich, Mumbai, India. ≥99% purity) was diluted in absolute ethanol and added to water. To note that ethanol was used at a minimal concentration of 2–3 drops of ethanol per 20 L of tank water, resulting in a final solvent concentration of approximately 0.0006% v/v, which accounts for final dilution <1:10000, while melatonin (Sigma-Aldrich, Mumbai, India) was prepared similarly to a working concentration of 100 ng/mL and administered via water. The selection of these doses was based on earlier reports demonstrating that BPA at environmentally relevant concentrations in the low μg/mL range can induce measurable biochemical and molecular alterations in zebrafish, while melatonin at nanogram levels has been shown to exert protective and modulatory effects against oxidative and endocrine disruption [4,11]. Zebrafish were exposed for 21 days under static immersion with daily partial renewal of water, alongside routine feeding. Water quality parameters were monitored daily. At the end of exposure, fish were anesthetized with buffered tricaine methane sulfonate (MS-222, 150 mg/L; pH 7.0) until loss of reflex, and samples were collected for biochemical, histological, and gene expression studies, with tissues preserved in TRIzol™ Reagent (Himedia, Mumbai, India) at −80°C (for gene expression) and fixed in 4% paraformaldehyde (for histology).
2.3. Growth Performance and Survival Rate (SR)
Growth performance of zebrafish was assessed at the end of the experimental period by calculating weight gain (WG), specific growth rate (SGR), standard length (SL), and SR. Individual BWs and SLs were recorded at the beginning and conclusion of the 21-day feeding trial. WG (g) was determined as the difference between final and initial BWs, while SGR (%) was calculated according to the formula:
SL (cm) was calculated by subtracting the initial body length from the final measurement.
The SR% was calculated as:
All calculations were performed following established protocols [12,13], and results were expressed as mean ± standard error (SE).
2.4. Gonadosomatic Index (GSI)
GSI was calculated to evaluate reproductive status. The GSI was calculated by relating gonad weight to total BW [14]. Following euthanasia, individual fish were blotted dry, and total BW was recorded. Gonads were carefully excised, weighed (GW), and the GSI was calculated using the formula:
Values were expressed as percentages for statistical analysis.
2.5. Lipid Peroxidation (LPO)
LPO was estimated by quantifying malondialdehyde (MDA), a stable end product of LPO, using the thiobarbituric acid reactive substances method with a commercial kit (EZAssay™ CCK023, Himedia, India). Tissue homogenates (100 mg) were prepared in phosphate-buffered saline (PBS), and 25 μL of homogenate was mixed with 500 μL of color development reagent. Samples were incubated in a boiling water bath for 60 min, cooled on ice, and centrifuged at 3,000 rpm for 10 min. The supernatant (150 μL) was transferred to a 96-well plate, and absorbance was measured at 530–540 nm. MDA concentrations were determined from a standard curve (2–10 μM) and expressed as mmol/g tissue, normalized to total protein content determined by Bradford’s method [15].
2.6. Superoxide Dismutase (SOD)
SOD activity was determined by its ability to inhibit the reduction of nitro blue tetrazolium (NBT) in the presence of superoxide radicals, following the method based on phenazine methosulfate (PMS)–NADH–NBT reaction [16]. Tissue homogenates (100 mg) were prepared in PBS, and the assay mixture contained sodium pyrophosphate buffer (0.052 mM, pH 8.3), PMS (186 μM), NADH (780 μM), and NBT (300 μM). The reaction was initiated by NADH addition, incubated at 37°C for 90 s, and terminated with acetic acid, followed by extraction in n-butanol. Absorbance was measured at 560 nm, and activity was expressed as units per gram of tissue, where one unit corresponds to 50% inhibition of NBT reduction. Results were normalized to total protein content determined by Bradford’s method [15].
2.7. Catalase (CAT)
CAT activity was measured by monitoring the rate of hydrogen peroxide (H2O2) decomposition at 240 nm [17]. Tissue homogenates (10% w/v) were prepared in PBS (0.1 M, pH 7.4) and centrifuged at 1,000 rpm to remove debris. The supernatant was treated with ethanol and incubated in an ice water bath for 30 min, followed by the addition of Triton X-100 (10%) and phosphate buffer immediately before the assay. The reaction mixture contained phosphate buffer (50 mM, pH 7.0) and freshly prepared H2O2 (30 mM), and the reaction was initiated by enzyme addition. The decrease in absorbance at 240 nm was recorded at 5-s intervals for 15 s. CAT activity was calculated using the formula:
Expressed as mmol H2O2 decomposed per second per gram of tissue. Results were normalized to total protein content determined by Bradford’s method [15].
2.8. Gene Expression
Total RNA was extracted from the gut and ovary using TRIzol™ Reagent (Himedia, Mumbai, India) following the manufacturer’s protocol [18]. RNA concentration and purity were assessed using NanoDrop™ 2000 spectrophotometer (Thermo Scientific) at 260/280 nm, and samples with ratios between 1.8 and 2.0 were further considered for expression studies. Complementary DNA (cDNA) was synthesized from 1 μg of total RNA using the High-Capacity cDNA Reverse Transcription Kit (ThermoFisher Scientific) with random hexamer primers.
Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYBR Green PCR Master Mix (ThermoFisher Scientific) on a StepOne Real-Time PCR System (Thermofisher) to quantify the expression of targeted genes related to circadian rhythm (Clock1a, Cry3a, Per1b), melatonin signaling (MT1), intestinal barrier integrity (occludin, claudin), and inflammatory cytokines (Interleukin-6 [IL-6], tumor necrosis factor-alpha [TNF-α]). The housekeeping gene β-actin was used for normalization. Primer sequences were designed using NCBI Primer-BLAST to produce amplicons of 50–150 bp and verified for their specificity. All reactions were performed in triplicate to ensure reproducibility, and a no-template control was included for each primer set to monitor contamination or primer-dimer formation. PCR cycling conditions included an initial denaturation at 95°C for 2 min, followed by 40 cycles of 95°C for 15 s, 57–60°C for 15 s, and 72°C for 1 min, with melt curve analysis to confirm amplification specificity. Relative expression was calculated using the 2–ΔΔCt method [19]. qRT-PCR plates were prepared and analyzed using anonymized sample codes; group identities were revealed only after statistical analysis was completed. The list of primers is stated in Table 1.
Table 1: Primer list.
Gene name
Accession No. (NCBI)
Sequence
Tm (oC)
Amplicon length
β-actin
NM_181601.5
Forward Primer (5′ → 3′)
ACAGGGAAAAGATGACACAGATCA
68
72
Reverse Primer (5′ → 3′)
CAGCCTGGATGGCAACGTA
60
Clock1a
XM_073932134.1
Forward Primer (5′ → 3′)
CATCCTACAGAAGAGCATCGACTT
70
73
Reverse Primer (5′ → 3′)
GATTTCACTCGACTCCGACTGT
66
Per1b
NM_212439.3
Forward Primer (5′ → 3′)
ATCCAGACCCCAATACAAC
56
99
Reverse Primer (5′ → 3′)
GGGAGACTCTGCTCCTTCT
60
Cry3a
NM_001313822.1
Forward Primer (5′ → 3′)
GGACCAATACACCAGCACCAG
62
245
Reverse Primer (5′ → 3′)
CAGCAAGTGTCCTGCCATGTC
62
Ocln
NM_001008618.1
Forward Primer (5′ → 3′)
CAAAATCAGGCAAAGGCTTC
56
176
Reverse Primer (5′ → 3′)
AACAATAGTGGCGATGAGCA
60
Caln
NM_131764.1
Forward Primer (5′ → 3′)
GTACCCTCCGCAAAGTCGTA
60
130
Reverse Primer (5′ → 3′)
CTTTCAAGGAAAGACTGACAGC
62
IL-6
NM_001261449.1
Forward Primer (5′ → 3′)
TCAACTTCTCCAGCGTGATG
58
Reverse Primer (5′ → 3′)
TCTTTCCCTCTTTTCCTCCTG
62
TNF-α
NM_212859.2
Forward Primer (5′ → 3′)
GCTGGATCTTCAAAGTCGGGTGTA
72
138
Reverse Primer (5′ → 3′)
TGTGAGTCTCAGCACACTTCCATC
72
MT1
NM_131393.1
Forward Primer (5′ → 3′)
AAAAGCAAACCTTCCCCACT
58
185
Reverse Primer (5′ → 3′)
TTGCTTCGATTGTGTTGAGG
57
2.9. Digestive Enzyme Activities
2.9.1. α-amylase activity
α-Amylase activity in intestinal tissue was measured using a commercial assay kit (Reckon Diagnostic, India) following the manufacturer’s instructions. Briefly, 10% tissue homogenate was prepared in ice-cold distilled water, incubated at room temperature for 15 min, and centrifuged at 3,000 × g for 10 min. The supernatant was then incubated with the substrate, and reducing sugars produced were detected by reaction with 3,5-dinitrosalicylic acid, yielding a brown-red complex that was measured at 540 nm using a microplate reader. Enzyme activity was calculated from a standard curve and expressed as U/g tissue, where one unit represents the production of 1 mg of reducing sugar per minute. Values were normalized to total protein content determined by the Bradford method [15].
2.9.2. Alkaline phosphatase (ALP)
ALP activity was quantified using a commercial ELISA kit specific for fish ALP (MyBioSource), following the manufacturer’s instructions. Tissue homogenates were prepared in PBS (0.1 M, pH 7.4) and centrifuged at 3000 rpm for 20 min. The clear supernatant was collected for analysis.
For each assay, 50 μL of sample or standard was added to wells of a pre-coated 96-well plate, followed by 100 μL of Horseradish Peroxidase-conjugate reagent. Plates were incubated at 37°C for 60 min, washed 4 times, and sequentially treated with 50 μL of Chromogen Solution A and 50 μL of Chromogen Solution B. After a 15-min incubation at 37°C in the dark, 50 μL of stop solution was added, and absorbance was measured at 450 nm using a microplate reader.
A standard curve was generated from serial dilutions of the provided standards, and ALP activity in samples was calculated in accordance. Assays were run in triplicate, and enzyme activity was expressed as U/L, normalized to protein content estimated by Bradford’s method [15].
2.10. Histology
Histological examination was performed to assess the structural organization and cytoarchitecture of the gut and ovary [20,21]. Dissected tissues were rinsed in cold (PBS, pH 7.4) to remove blood and debris, and immersion-fixed in 4% paraformaldehyde at 4°C for 24–48 h. Following fixation, samples were dehydrated in graded ethanol, cleared in xylene, and embedded in paraffin wax. Sections of 4 μm thickness were prepared using a rotary microtome, mounted on poly-L-lysine-coated slides, and stained with hematoxylin and eosin following standard protocols [22]. The slides were examined under a bright-field inverted microscope (12818065, Axio Vert A1; Zeiss, Oberkochen, Baden-Württemberg, Germany) for morphological changes, and representative images were captured. Structural alterations, including disruption of gut and ovarian architecture, were also observed.
2.11. Statistical Analysis
The statistical analysis was done using two-way analysis of variance (ANOVA) with Dunnett’s test for comparison of mean values for gene expression studies, and the antioxidant assay analysis was done using one-way ANOVA with Tukey’s post hoc test. The histological quantification data were analyzed using one-way ANOVA using GraphPad Prism 8.4.2. All the experimental data were carried out in triplicate, where *P < 0.05, **P < 0.01, and ns represents non-significant values.
3. RESULTS
3.1. Growth Performance, Survival, and Somatic Indices
Growth performance and GSI varied significantly among all experimental groups [Table 2]. BPA exposure significantly reduced WG, SL, SGR, and survival compared to its control (P < 0.05). In contrast, melatonin supplementation alone slightly enhanced growth and survival relative to control. Simultaneous administration of melatonin with BPA led to partial recovery of growth indices, with values intermediate between the BPA and control groups.
Table 2: Growth performances, survival, and somatic indices of the experimental study.
Parameters
Weight gain (g)
Standard length (cm)
SGR (%)
SR (%)
GSI (%)
Control
0.052±0.004
0.166±0.010
0.242±0.016
96.66±1.66
0.343±0.004
BPA
0.033±0.005*
0.124±0.011*
0.168±0.017*
83.33±2.88*
0.253±0.004
Melatonin
0.056±0.003
0.173±0.009*
0.251±0.015
98.33±1.66
0.348±0.005
BPA+Melatonin
0.046±0.004
0.154±0.010
0.216±0.016
91.66±2.88
0.412±0.005
SGR: Specific growth rate, SR: Survival rate, GSI: Gonadosomatic index. Statistical significance compared with the control is denoted as
P < 0.05
Similarly, BPA markedly suppressed GSI relative to control (P < 0.05), while melatonin alone maintained values comparable to its control. Melatonin improved GSI when given simultaneously to BPA.
3.2. Antioxidant Parameters
3.2.1. Lipid peroxidase (LPO)
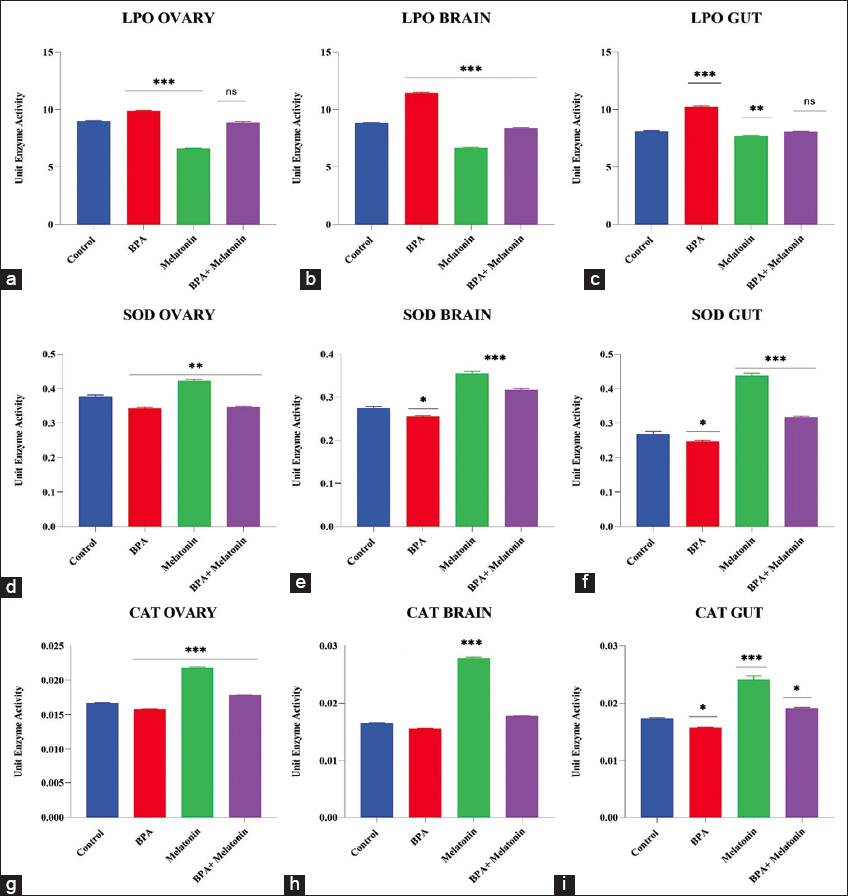
LPO, expressed as MDA equivalents, showed clear tissue-specific differences across treatment groups [Figure 2a-c]. In the ovary, BPA exposure significantly increased LPO compared to control (P < 0.001), while melatonin supplementation markedly reduced levels (P < 0.001). Melatonin restored values close to control when administered alongside BPA. A similar pattern was observed in the brain, where BPA enhanced oxidative stress (P < 0.001), melatonin alone suppressed peroxidation (P < 0.001), and the combined group showed intermediate values. In the gut, BPA again elevated LPO (P < 0.001), whereas melatonin reduced it (P < 0.01); BPA+Melatonin produced values comparable to control.
Figure 2: Antioxidant enzyme activities and lipid peroxidation in zebrafish tissues following BPA and melatonin treatments. Lipid peroxidation (a-c), superoxide dismutase (d-f), and catalase (g-i) levels were measured in the ovary, brain, and gut tissues of zebrafish across experimental groups: Control, BPA, Melatonin, and BPA + Melatonin. Data are presented as mean ± standard error mean (n = 9). Statistical significance compared with control is denoted as *P < 0.05, **P < 0.01, and ***P < 0.001; ns = non-significant.
SOD activity was significantly altered by BPA and melatonin treatments in all examined tissues [Figure 2b-f]. In the brain, BPA exposure reduced SOD activity compared with control (P < 0.05), whereas melatonin markedly enhanced activity (P < 0.001). Melatonin resulted in partial restoration, with values higher than BPA alone but not fully reaching control when given simultaneously to BPA.
A similar trend was observed in the gut, where BPA significantly suppressed SOD activity (P < 0.05). Melatonin supplementation strongly increased activity (P < 0.001), and co-treatment again produced intermediate values.
In the ovary, BPA exposure reduced SOD activity relative to control, while melatonin treatment elevated it significantly (P < 0.001). The combined group suggested moderate recovery compared with BPA alone but did not reach control levels.
3.2.3. CAT
CAT activity showed distinct responses across tissues [Figure 2g-i]. In the ovary, BPA exposure produced only minor changes compared with control, whereas melatonin treatment significantly increased activity (P < 0.001). Simultaneous treatment with BPA and melatonin resulted in a modest elevation above BPA alone.
In the brain, CAT activity remained unchanged by BPA but was strongly enhanced by melatonin supplementation (P < 0.001). The BPA + melatonin group displayed intermediate levels, higher than BPA alone but lower than melatonin treatment.
In the gut, BPA slightly reduced CAT activity (P < 0.05), while melatonin markedly increased it (P < 0.001). Combined treatment partially restored activity, yielding values closer to control.
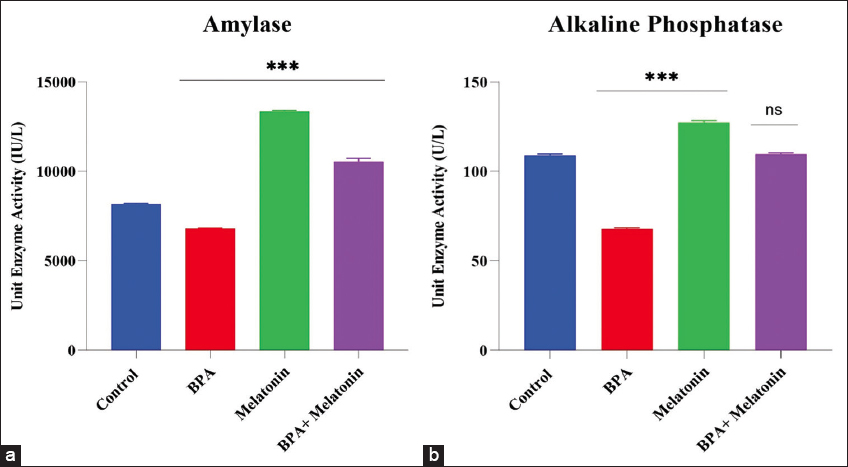
3.3 α-amylase activity
α-Amylase activity varied significantly across treatment groups [Figure 3a]. BPA exposure caused a pronounced reduction compared with the control (P < 0.001), indicating impaired digestive function. In contrast, melatonin supplementation markedly enhanced enzyme activity relative to control (P < 0.001). Combined with BPA and melatonin, it partially restored activity.
Figure 3: Effects of bisphenol A (BPA) and melatonin on digestive enzyme activities in zebrafish. (a) α-Amylase activity (IU/L) across control, BPA, melatonin, and BPA + melatonin groups. BPA exposure significantly reduced amylase activity compared with control, while melatonin markedly enhanced it. Co-treatment with BPA + melatonin partially restored enzyme activity. (b) Alkaline phosphatase (ALP) activity (U/L) across experimental groups. BPA exposure decreased ALP activity, melatonin significantly increased it, and BPA + melatonin co-treatment restored values closer to control. Data are presented as mean ± standard error mean (n = 9 per group). Statistical significance: *P < 0.05, **P < 0.01, ***P < 0.001; ns, not significant.
ALP activity showed clear treatment-dependent variations [Figure 3b]. BPA exposure significantly suppressed enzyme activity compared with the control group (P < 0.001), indicating a marked sensitivity of ALP to xenobiotic stress. Melatonin supplementation alone, in contrast, produced a strong stimulatory effect, with activity values exceeding control levels (P < 0.001). In the combined group, ALP activity was partially restored, approaching control levels and supporting melatonin’s capacity to counteract BPA-induced inhibition.
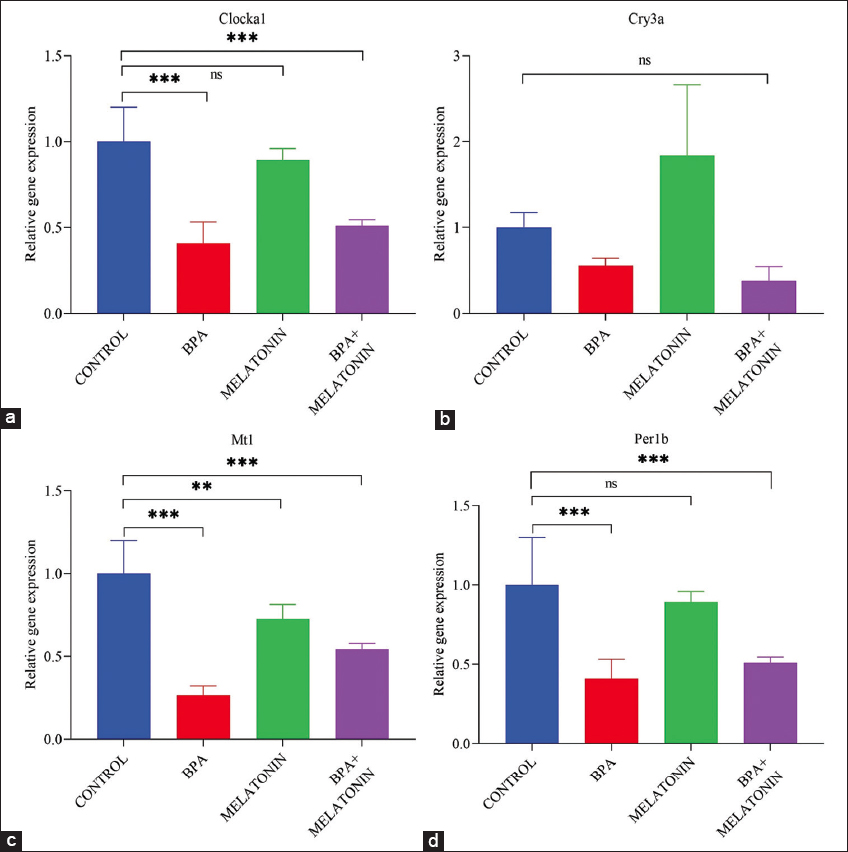
3.5. Gene Expression
Gene expression analysis revealed significant disruptions in circadian regulation, barrier integrity, and inflammatory signaling following BPA exposure, while melatonin supplementation exhibited both protective and restorative effects. Among circadian rhythm–associated genes, BPA markedly suppressed Clock1a, Cry3a, MT1, and Per1b expression relative to controls (P < 0.001) [Figure 4a-d]. Melatonin supplementation, in contrast, significantly upregulated these transcripts, suggesting a stimulatory influence on circadian regulation. Combined treatment with BPA and melatonin partially restored expression, with values higher than BPA alone but lower than melatonin-only treatment, indicating an intermediate protective effect.
Figure 4: Relative gene expression analysis of circadian rhythm–associated and tight junction genes in zebrafish exposed to different treatments. (a) Claudin, (b) Cry3a, (c) MT1, and (d) Per1b expression levels were quantified by quantitative polymerase chain reaction across Control, bisphenol A (BPA), Melatonin, and BPA + Melatonin groups. BPA exposure significantly downregulated Claudin, MT1, and Per1b, while melatonin supplementation upregulated these genes compared to control. Co-treatment with BPA + melatonin partially restored expression toward control levels. Cry3a expression showed a non-significant trend across groups. Data are presented as mean ± standard error mean (n = 9 per group). Statistical analysis was performed using one-way analysis of variance followed by Tukey’s post hoc test. ***P < 0.001, **P < 0.01, ns = not significant.
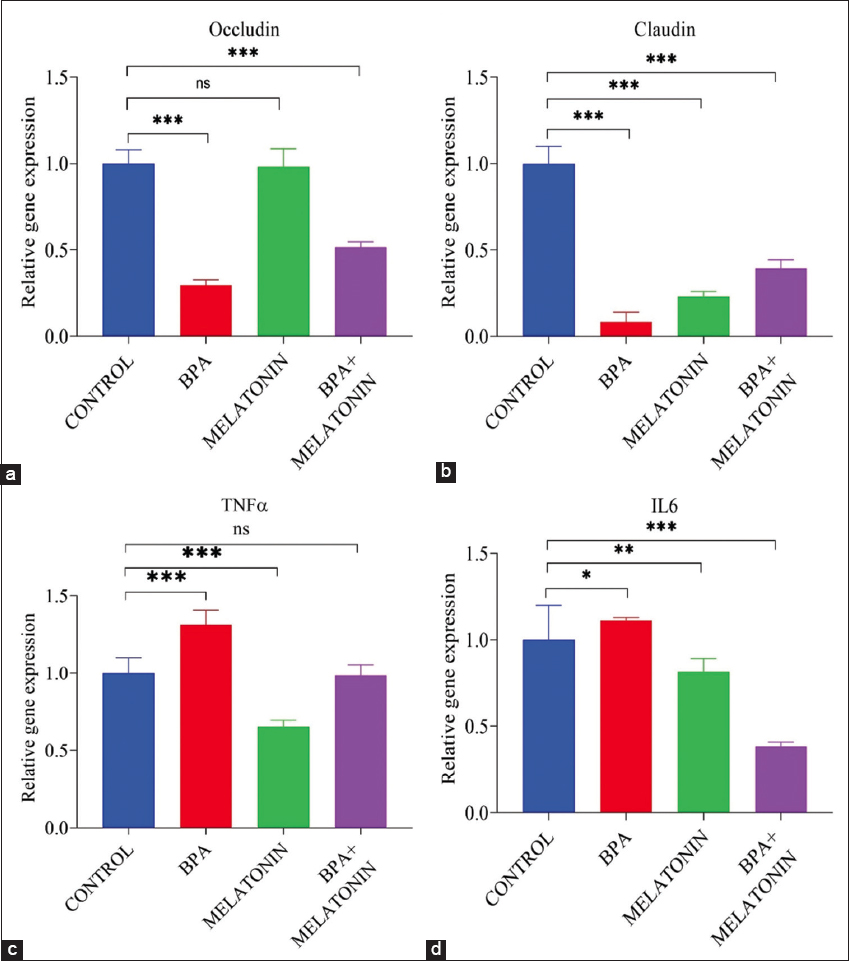
For intestinal barrier markers, BPA exposure significantly downregulated Claudin and Occludin (P < 0.001), reflecting impaired epithelial integrity. Melatonin supplementation enhanced the expression of both genes, while combined treatment indicated significant recovery, though expression levels did not fully return to control values [Figure 5a and b].
Figure 5: Effects of bisphenol A (BPA) and melatonin on the expression of intestinal barrier markers and inflammatory cytokines in zebrafish. (a) Occludin and (b) Claudin mRNA expression were significantly downregulated in the BPA group compared to the control, whereas melatonin treatment restored expression levels. Co-treatment (BPA + melatonin) partially ameliorated the suppression. (c) Tumour Necrosis Factor-alpha α and (d) Interleukin-6 expression levels were markedly upregulated in the BPA group, indicating a pro-inflammatory response. Melatonin alone reduced cytokine expression, while co-treatment significantly attenuated BPA-induced elevation. Data are represented as mean ± standard error mean (n = 9). Statistical significance: P < 0.05, P < 0.01, P < 0.001, ns = not significant.
With respect to inflammatory mediators, BPA markedly upregulated IL-6 and TNF-α, indicating a pronounced pro-inflammatory response. Melatonin supplementation counteracted these effects, significantly reducing expression of both cytokines. Importantly, BPA + melatonin substantially attenuated the BPA-induced elevation, with transcript levels approaching those observed in control [Figure 5c and d].
3.6. Ovarian Histology
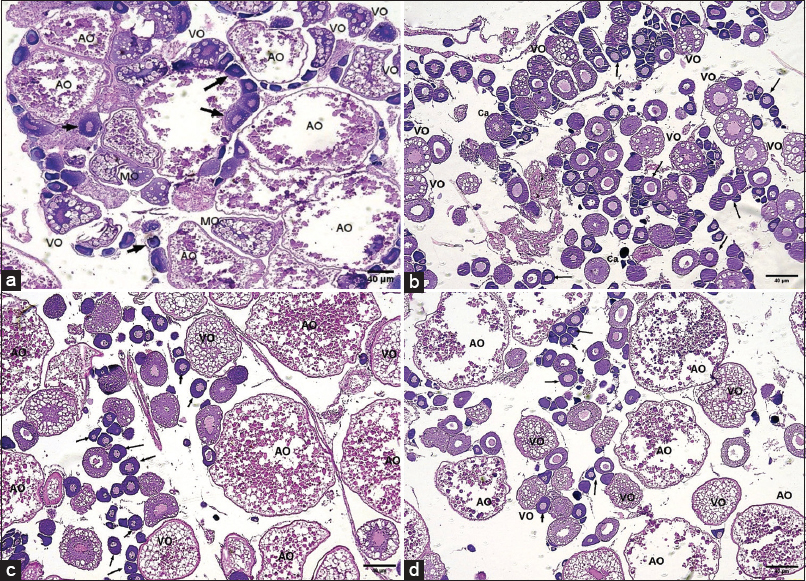
Histological examination of the ovaries demonstrated clear group-dependent alterations in follicular dynamics and overall tissue architecture as per Figure 6a-d. In the control group, ovaries exhibited a moderate but balanced representation of all follicular stages, including primary follicles, cortical alveoli, vitellogenic, and mature oocytes, consistent with normal asynchronous oogenesis. Occasional atretic oocytes were observed, though their incidence remained low, reflecting physiological turnover.
Figure 6: Histological analysis of zebrafish ovary sections under different treatments (Hematoxylin and Eosin staining, 10× magnification, scale bar = 40 μm). (a) Control group showing normal ovarian architecture with a balanced distribution of primary follicles, vitellogenic oocytes, mature oocytes (Marked arrows-black), and atretic oocytes. (b) bisphenol A (BPA)-exposed group displaying disrupted folliculogenesis with an increased proportion of primary and vitellogenic oocytes, fewer atretic oocytes, and limited mature follicle formation, indicative of follicular arrest. (c) Melatonin-treated group demonstrating enhanced follicular development with abundant vitellogenic and mature oocytes, along with a higher frequency of atretic oocytes, suggesting stimulation of folliculogenesis. (d) BPA + melatonin co-treated group showing near-normal ovarian morphology with follicular distribution resembling control levels, indicating a restorative effect of melatonin against BPA-induced disruption.
In contrast, ovaries from BPA-exposed fish displayed a striking disruption in follicular progression. These samples contained a disproportionately high number of primary and vitellogenic follicles, while the frequency of mature oocytes was markedly reduced. The relative paucity of atretic oocytes suggested that follicular degeneration was not the predominant pathology; rather, BPA exposure appeared to impose an arrest in folliculogenesis, with follicles unable to progress efficiently to later developmental stages. This pattern underscores the inhibitory impact of BPA on ovarian maturation.
Ovaries from melatonin-treated fish, however, presented a distinctly different profile. Follicles across all developmental stages were more abundant, with an especially notable increase in vitellogenic and mature oocytes, alongside the presence of well-developed cortical alveoli. Atretic oocytes were comparatively rare. These findings indicate that melatonin promotes follicular recruitment and maturation, highlighting its supportive role in maintaining ovarian function.
When BPA and melatonin were administered together, it largely restored ovarian architecture toward that of the controls. The distribution of primary, vitellogenic, and mature follicles, as well as cortical alveoli, resembled the control profile, and the incidence of atretic oocytes remained low. This restorative effect strongly suggests that melatonin mitigates the deleterious consequences of BPA exposure on folliculogenesis, thereby preserving the integrity of ovarian morphology and supporting normal reproductive physiology.
3.7. Intestinal Histology
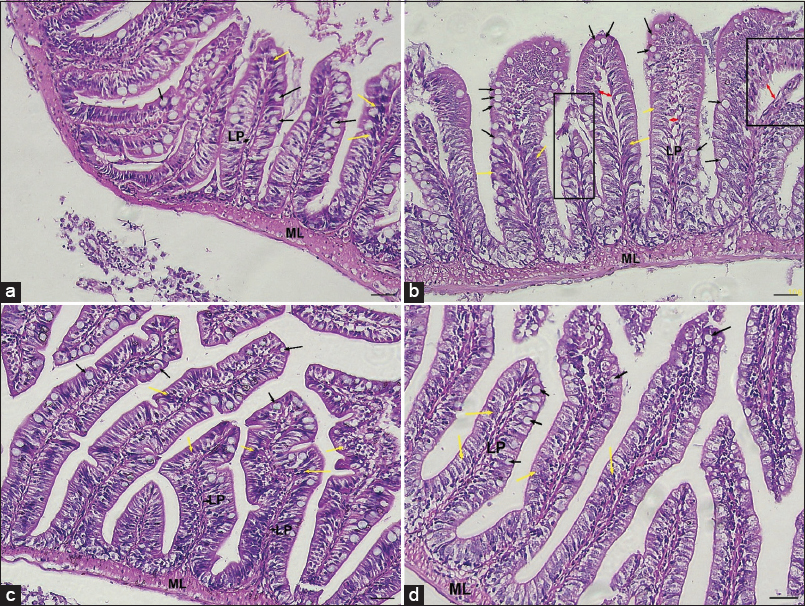
Histological evaluation of the zebrafish gut revealed distinct alterations across the treatment groups. In the control group, the intestine exhibited normal architecture, characterized by well-preserved and elongated villi, intact epithelial lining, and a clearly defined lamina propria without signs of structural disruption. Enterocytes were orderly arranged with intact brush borders, and goblet cells were present in moderate numbers, indicative of healthy mucosal function [Figure 7a].
Figure 7: Representative histological sections of zebrafish gut (Hematoxylin and Eosin staining, 10× magnification; scale bar = 40 μm) across experimental groups. (muscularis layer [ML]; Lamina propria [LP], black arrows- Goblet cells; yellow arrows- Enterocytes; red arrows- Widened Lamina propria), (a) Control gut showing intact and well-organized villi with narrow inter-villus spaces, regular epithelial lining, and a compact LP. (b) bisphenol A (BPA)-exposed gut exhibiting disrupted architecture with shortened and damaged villi, widened inter-villus gaps, epithelial cell disorganization, and vacuolization within the lamina propria, indicating structural injury. (c) Melatonin-treated gut demonstrates preserved histoarchitecture, with elongated villi, intact epithelium, and a well-defined lamina propria, reflecting enhanced tissue integrity. (d) BPA + Melatonin co-treated gut showing near-normal morphology with partially restored villus length, reduced gaps at the lamina propria, and improved epithelial continuity compared to BPA-exposed samples, indicating a restorative effect of melatonin.
In contrast, the BPA-exposed group displayed marked histopathological damage as shown in Figure 7b. The intestinal villi were shortened and blunted, with evident epithelial disruption and vacuolation. The lamina propria exhibited widened intercellular gaps, suggestive of compromised barrier integrity, and the overall tissue structure appeared disorganized. These features are consistent with BPA-induced impairment of epithelial homeostasis and disruption of mucosal defense.
The melatonin-treated group demonstrated a protective morphological profile. The intestinal villi were long and slender, with intact epithelial continuity and well-defined lamina propria. Goblet cells were maintained in higher numbers compared to controls, contributing to enhanced mucosal barrier function. The overall tissue architecture appeared to be preserved, with minimal evidence of degeneration or inflammation, highlighting melatonin’s barrier-protective effects, as shown in Figure 7c.
Finally, the BPA + melatonin combined group exhibited a near-complete restoration of intestinal architecture, as shown in Figure 7d. The villi appeared elongated and structurally organized, with epithelial continuity largely re-established. The lamina propria was compact, showing reduced widening of intercellular spaces compared to BPA alone. Goblet cell distribution and epithelial organization approached control levels, reflecting the restorative influence of melatonin against BPA-induced intestinal injury.
4. DISCUSSION
The present study provides a comprehensive evaluation of the adverse effects of BPA on zebrafish, spanning molecular gene expression, histopathology, and organismal performance, while also highlighting the protective and modulatory roles of melatonin.
4.1. Growth Performance and Survival Outcomes
Growth performance indicators demonstrated clear evidence of BPA toxicity. Zebrafish exposed to BPA exhibited significant reduced weight gain, SL, and SGR, together with decreased survival. These findings corroborate earlier reports linking BPA exposure to growth retardation and reduced survivability in fish models due to endocrine disruption and metabolic dysregulation [4,23,24]. In contrast, melatonin supplementation alone marginally enhanced growth and survival compared to controls, consistent with its antioxidant and metabolic regulatory properties [9]. Importantly, simultaneous melatonin treatment partially restored growth parameters and survival in BPA-exposed groups, underscoring its role as a mitigator of endocrine and oxidative stress–mediated damage.
4.2. Oxidative Stress and Antioxidant Responses
BPA exposure perturbs redox balance in zebrafish tissues, as indicated by elevated LPO and reduced efficiency of enzymatic antioxidants, such as SOD and CAT. This pattern is consistent with previous findings that bisphenols generate reactive oxygen species (ROS), impair antioxidant defense systems, and thereby promote oxidative injury in teleosts [25,26]. In our study, the brain and gut exhibited marked increases in LPO under BPA, paralleled by villus damage and pro-inflammatory gene expression, suggesting that oxidative stress is a key driver of tissue pathology.
Simultaneous administration of melatonin with BPA markedly reduced LPO and enhanced SOD and CAT activities across tissues. These outcomes align well with the established evidence of melatonin’s antioxidant properties, both as a direct free radical scavenger and as an inducer of antioxidant enzymes via Nrf2-dependent signaling [27]. In mammals, melatonin has been shown to counteract BPA-induced oxidative damage, and our results extend these protective effects to fish, particularly in reproductive and gut tissues, where structural restoration paralleled improvements in redox balance [9].
Notably, the ovarian data link oxidative stress with reproductive dysfunction. Elevated oxidative load under BPA coincided with the arrest of folliculogenesis and reduced mature oocyte formation, whereas melatonin improved antioxidant tone and partially restored follicular progression. Similar associations between ROS burden and reproductive impairment have been reported in fish exposed to environmental estrogen [25,26,28-31].
However, the antioxidant responses were tissue-specific, and CAT activity remained relatively unchanged under BPA in some organs. This variability is consistent with prior reports showing that CAT regulation under xenobiotic stress is context-dependent, possibly reflecting compensatory involvement of glutathione peroxidase or other antioxidant pathways [32].
4.3. Molecular Responses and Inflammatory Signaling
Expression analysis of circadian rhythm–related genes revealed pronounced downregulation of Clock1a, Cry3a, and Per1b in the BPA group, aligning with evidence that BPA disrupts circadian regulation and metabolic homeostasis [33]. Melatonin treatment alone enhanced expression of these genes, while given simultaneously to BPA showed partial restoration, suggesting that melatonin stabilizes circadian rhythms even under sustained oxidative stress. Similarly, BPA suppressed MT1 receptor and tight-junction gene (Claudin, Occludin) expression, while pro-inflammatory mediators (IL-6, TNF-α) were strongly upregulated. These molecular alterations are consistent with barrier dysfunction and heightened inflammatory tone, both of which have been reported in rodent and fish models of BPA exposure studies [34-36]. Melatonin supplementation countered these effects by upregulating barrier proteins and downregulating inflammatory markers, in agreement with its known role as an anti-inflammatory and gut barrier–protective agent [37,38].
4.4. Ovarian Histology and Reproductive Effects
Histological analysis of ovaries revealed significant impairments in folliculogenesis in the BPA-exposed group. Specifically, a higher prevalence of primary and vitellogenic follicles with reduced mature follicles and corpus luteum suggests arrested follicular progression. This pattern indicates an interference with normal ovarian maturation, consistent with endocrine-disrupting effects of BPA reported in fish and mammalian ovaries [31,39]. In contrast, melatonin-treated ovaries displayed an abundance of follicles at all developmental stages, alongside healthier atretic oocytes and cortical alveoli, indicative of enhanced reproductive capacity [40]. Notably, combined treatment with BPA and melatonin largely restored ovarian architecture to near-control levels, suggesting melatonin’s ability to ameliorate BPA-induced arrest in folliculogenesis.
4.5. Intestinal Histopathology and Barrier Function
Examination of gut histology further highlighted the deleterious impact of BPA. Compared with the well-organized villi, intact mucosal layers, and compact lamina propria observed in controls, BPA-treated fish exhibited shorter and damaged villi, widened gaps within the lamina propria, and compromised epithelial integrity. These disruptions are characteristic of chemical-induced gut barrier dysfunction [29,30]. Melatonin alone maintained healthy villi morphology, whereas BPA + melatonin markedly ameliorated the structural damage, supporting molecular findings of restored tight-junction protein expression. Together, these results point toward melatonin’s ability to preserve intestinal homeostasis under toxicant stress [9,28,41].
By integrating molecular, histological, and physiological data, this study highlights the multifaceted toxicity of BPA, ranging from impaired circadian regulation and inflammation to reproductive and intestinal dysfunction, culminating in reduced growth and survival. Importantly, melatonin exerted consistent protective effects across all levels of biological organization. These findings extend previous work on BPA’s endocrine-disrupting properties. They also provide novel insights into melatonin’s potential as a therapeutic modulator against environmental toxicants in aquatic organisms.
5. CONCLUSION
Our findings indicate that BPA disrupts zebrafish physiology at molecular, histological, and organismal levels, impairing circadian regulation, barrier integrity, growth, and reproductive outcomes. Melatonin treatment consistently mitigated these alterations, restoring gene expression, protecting ovarian and gut architecture, and improving growth performance and survival. These results highlight melatonin’s protective role against endocrine-disrupting chemicals, with potential applications in aquaculture and environmental toxicology. Melatonin regulates circadian rhythms, gut-barrier integrity, and inflammation. These properties support its potential application in aquaculture and mammalian research. The study also indicates that a higher dose of melatonin may serve to completely reverse the BPA-induced toxicity. This study indicates that BPA exposure may disturb both gut and ovarian physiology, and that melatonin treatment ameliorates these alterations, suggesting a possible coordination between the two systems rather than a direct mechanistic cross-talk. However, further studies on metabolite signaling will provide intricate insights into the matter.
6. ACKNOWLEDGMENTS
The authors would like to acknowledge the SHODH fellowship awarded to Yashshvini Patel and Navrachana University for providing the necessary infrastructure and support to carry out the research work.
7. AUTHOR CONTRIBUTION STATEMENT
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. CONFLICT OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
The study protocol was approved by the Institutional Ethics Committee of Division of Biomedical and Life Sciences, School of Science, Navrachana University, Gujarat, India (Approval No.: AEC/01/Inst/BMS/2022).
10. DATA AVAILABILITY
All data generated or analyzed during this study are included in this published article and are available from the corresponding author upon reasonable request.
11. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
12. Use of artificial intelligence (AI)-assisted technology
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Sun C, Wang L, Lu Y, Zhen H, Xiu G. The profiles and potential risk of Bisphenol A and its analogues of ambient airborne fine particulate matter in a chemical industrial park in Shanghai. Atmos Environ. 2025;359:121349.[CrossRef]
2. Rubin BS. Bisphenol A. An endocrine disruptor with widespread exposure and multiple effects. J Steroid Biochem Mol Biol. 2011;127:27-34.[CrossRef]
3. Della Rocca Y, Traini EM, Diomede F, Fonticoli L, Trubiani O, Paganelli A, et al. Current evidence on bisphenol a exposure and the molecular mechanism involved in related pathological conditions. Pharmaceutics. 2023;15:908.[CrossRef]
4. Santangeli S, Maradonna F, Gioacchini G, Cobellis G, Piccinetti CC, Dalla Valle L, et al. BPA-induced deregulation of epigenetic patterns: Effects on female zebrafish reproduction. Sci Rep. 2016;6:21982.[CrossRef]
5. Chen L, Guo Y, Hu C, Lam PK, Lam JC, Zhou B. Dysbiosis of gut microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol A: Implications for host health in zebrafish. Environ Pollut. 2018;234:307-17. [CrossRef]
6. Kamfar WW, Khraiwesh HM, Ibrahim MO, Qadhi AH, Azhar WF, Ghafouri KJ, et al. Comprehensive review of melatonin as a promising nutritional and nutraceutical supplement. Heliyon. 2024;10:24266. [CrossRef]
7. Lunkes LC, Paiva IM, Egger RC, Braga WF, Alvarez-Leite JI, Da Cunha Barreto-Vianna AR, et al. Melatonin administration attenuates acute stress by inducing sleep state in zebrafish (Danio rerio). Comp Biochem Physiol Part C Toxicol Pharmacol. 2021;246:109044.[CrossRef]
8. Kopustinskiene DM, Bernatoniene J. Molecular mechanisms of melatonin-mediated cell protection and signaling in health and disease. Pharmaceutics 2021;13:129.[CrossRef]
9. Joo SS, Yoo YM. Protective effect of melatonin against bisphenol a toxicity. Int J Mol Sci. 2025;26:7526.[CrossRef]
10. Baker ME, Hardiman G. Transcriptional analysis of endocrine disruption using zebrafish and massively parallel sequencing. J Mol Endocrinol. 2014;52:R241-56.[CrossRef]
11. Choi JY, Choe JR, Lee TH, Choi CY. Effects of bisphenol A and light conditions on the circadian rhythm of the goldfish Carassius auratus. Biol Rhythm Res 2018;49:502-14.[CrossRef]
12. Akbary P, Aminikhoei Z. Effect of water-soluble polysaccharide extract from the green alga Ulva rigida on growth performance, antioxidant enzyme activity, and immune stimulation of grey mullet Mugil cephalus. J Appl Phycol. 2018;30:1345-53.[CrossRef]
13. Sunarma A, Carman O, Zairin M, Alimuddin A. Interpopulation crossbreeding of farmed and wild African catfish Clarias gariepinus (Burchell 1822) in Indonesia at the nursing stage. Aquat Living Resour. 2016;29:303.[CrossRef]
14. Gabr HR, Hanlon RT, Hanafy MH, El-Etreby SG. Maturation, fecundity and seasonality of reproduction of two commercially valuable cuttlefish, Sepia pharaonis and S., in the Suez Canal Fish Res. 1998;36:99-115.[CrossRef]
15. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248-54.[CrossRef]
16. Van Noorden CJ, Butcher RG. The involvement of superoxide anions in the nitro blue tetrazolium chloride reduction mediated by NADH and phenazine methosulfate. Anal Biochem 1989;176:170-4.[CrossRef]
17. Hadwan MH. Simple spectrophotometric assay for measuring catalase activity in biological tissues. BMC Biochem. 2018;19:7.[CrossRef]
18. Rio DC, Ares M, Hannon GJ, Nilsen TW. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb Protoc. 2010;2010(6):pdb.prot5439.[CrossRef]
19. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods. 2001;25:402-8.[CrossRef]
20. Flores EM, Nguyen AT, Odem MA, Eisenhoffer GT, Krachler AM. The zebrafish as a model for gastrointestinal tract-microbe interactions. Cell Microbiol. 2020;22:13152.[CrossRef]
21. Koç(Yön) ND, Aytekin Y, Yüce R. Ovary maturation stages and histological investigation of ovary of the Zebrafish (Danio rerio). Braz Arch Biol Technol. 2008;51:513-22.[CrossRef]
22. Fischer AH, Jacobson KA, Rose J, Zeller R. Hematoxylin and eosin staining of tissue and cell sections. Cold Spring Harb Protoc. 2008;2008:pdb.prot4986.[CrossRef]
23. Chen Y, Wu K, Huang W. Parental BPA exposure disrupts offspring transcriptomic profiles and impairs embryonic lineage specification during zygotic genome activation. J Hazard Mater Lett. 2025;6:100146.[CrossRef]
24. Wang J, Wang X, Xiong C, Liu J, Hu B, Zheng L. Chronic bisphenol A exposure alters behaviors of zebrafish (Danio rerio). Environ Pollut. 2015;206:275-81.[CrossRef]
25. Gu Z, Jia R, He Q, Cao L, Du J, Jeney G, et al. Oxidative stress, ion concentration change and immune response in gills of common carp (Cyprinus carpio) under long-term exposure to bisphenol A. Comp Biochem Physiol Part C Toxicol Pharmacol. 2020;230:10∇.[CrossRef]
26. Han Y, Fei Y, Wang M, Xue Y, Liu Y. Effects of BPZ and BPC on oxidative stress of zebrafish under different pH conditions. Molecules. 2022;27:1568. https://doi.org/10.3390/molecules27051568[CrossRef]
27. Ding K, Wang H, Xu J, Li T, Zhang L, Ding Y, et al. Melatonin stimulates antioxidant enzymes and reduces oxidative stress in experimental traumatic brain injury: The Nrf2-ARE signaling pathway as a potential mechanism. Free Radic Biol Med. 2014;73:1-11.[CrossRef]
28. Ahmadi S, Taghizadieh M, Mehdizadehfar E, Hasani A, Khalili Fard J, Feizi H, et al. Gut microbiota in neurological diseases: Melatonin plays an important regulatory role. Biomed Pharmacother. 2024;174:116487.[CrossRef]
29. Feng L, Chen S, Zhang L, Qu W, Chen Z. Bisphenol A increases intestinal permeability through disrupting intestinal barrier function in mice. Environ Pollut. 2019;254:112960.[CrossRef]
30. Hu C, Bai Y, Sun B, Tang L, Chen L. Significant impairment of intestinal health in zebrafish after subchronic exposure to methylparaben. Sci Total Environ. 2022;838:156389.[CrossRef]
31. Sudhakaran G, Kesavan D, Ranjan Nayak SP, Madesh S, Meenatchi R, Pachaiappan R, et al. Bisphenol A-induced ovarian damage countered by luteolin: Experiments in in vitro CHO cells and in vivo PCOS phenotype zebrafish. Tissue Cell. 2024;91:102532.[CrossRef]
32. Jomova K, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, Valko M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol. 2024;98:1323-67.[CrossRef]
33. Patisaul HB, Sullivan AW, Radford ME, Walker DM, Adewale HB, Winnik B, et al. Anxiogenic effects of developmental bisphenol a exposure are associated with gene expression changes in the juvenile rat amygdala and mitigated by soy. PLoS One. 2012;7:43890.[CrossRef]
34. Costa HE, Cairrao E. Effect of bisphenol A on the neurological system: A review update. Arch Toxicol. 2024;98:1-73.[CrossRef]
35. Rochester JR. Bisphenol A and human health: A review of the literature. Reprod Toxicol. 2013;42:132-55.[CrossRef]
36. Giommi C, LombóM, Habibi HR, Rossi G, Basili D, MangiaterraS, et al. The probiotic SLAB51 as agent to counteract BPA toxicity on zebrafish gut microbiota-liver-brain axis. Sci Total Environ. 2024;912:169303.[CrossRef]
37. Esteban MÁ, Cuesta A, Chaves-Pozo E, Meseguer J. Influence of melatonin on the immune system of fish: A review. Int J Mol Sci. 2013;14:7979-99.[CrossRef]
38. Cho JH, Bhutani S, Kim CH, Irwin MR. Anti-inflammatory effects of melatonin: A systematic review and meta-analysis of clinical trials. Brain Behav Immun. 2021;93:245-53.[CrossRef]
39. Canesi L, Fabbri E. Environmental effects of BPA. Dose Response. 2015;7;28;13(3):1559325815598304.[CrossRef]
40. Zhang J, Li F, Zhang X, Xie T, Qin H, Lv J, et al. Melatonin improves turbot oocyte meiotic maturation and antioxidant capacity, inhibits apoptosis-related genes mRNAs in vitro. Antioxidants. 2023;12:1389.[CrossRef]
41. Bonmatí-Carrión MÁ, Rol MA. Melatonin as a Mediator of the Gut Microbiota-Host Interaction: Implications for health and disease. Antioxidants. 2023;13:34.[CrossRef]