
1. INTRODUCTION
Rice root-knot nematodes (RKN), specifically Meloidogyne graminicola, are reportedly among the most devastating plant parasitic nematodes affecting rice production worldwide. The first trace of M. graminicola in Indonesia was found on rice in Yogyakarta Province in 1993 [1]. Previous research reported that M. graminicola infected over 100 plant species from cereals to grasses and dicotyledonous plants, but the most severe was on rice, causing up to 70% loss of harvest [2]. Meloidogyne graminicola is claimed to be responsible for 16% to 73% yield loss across various crop cultivation systems [3].
The symptom characteristics of M. graminicola appear below the ground in the form of hook-like galls on the roots and above the ground, such as stunted growth and chlorotic leaves [4,5]. Diagnosis based on the nematode symptom characteristics is currently challenging, so it is crucial to identify nematode species to obtain a more precise diagnosis and to manage the M. graminicola population. Traditional identification methods based on morphological and morphometric characteristics need specialized expertise, are time-consuming, and potentially subjective, especially for limited samples. In contrast, molecular identification is empirically more efficient, effective for larger samples, and feasible for studying phylogenetic relationships and nucleotide variation among populations [6,7]. Recent studies have increasingly focused on ribosomal DNA (rDNA) genes to identify species and phylogenetic analysis [8], assess phylogenetic relationships, analyze genetic variation, and study genome evolution [9]. The rDNA genes consist of conserved coding regions (e.g., 28S, 18S, and 5.8S subunit genes) and noncoding regions. The 18S gene, universally present in eukaryotes, is an effective marker for nematode barcoding and biodiversity surveys using the Sanger sequencing technique [10]. The 5.8S rDNA gene is more highly conserved than the 18S and 28S rDNA genes, making it ideal for phylogenetic studies at the species and population levels [11].
In Indonesia, morphological and morphometric detections of M. graminicola are reported in West Java [12] and South Sulawesi [13], but molecular detection and characteristic methods are understudied in Java Island. This study aimed to fill this gap by investigating the molecular characteristics of the M. graminicola population in Java, Indonesia, to obtain an accurate diagnosis of the disease for better preventive measures.
2. MATERIALS AND METHODS
2.1. Nematode Collection
Rice RKN samples were collected purposively from three rice fields in three Java provinces: Sarwadadi Village in Cirebon District, West Java (6°47'17.7"S+108°30'13.0"E; 6°47'16.1"S+108°30'11.6"E; 6°47'18.3"S 108°30'11.9"E), Tambaksari Village in Banyumas District, Central Java (7°24'04.1"S 109°15'34.3"E; 7°23'49.5"S 109°15'38.1"E; 7°23'51.6"S 109°15'37.3"E), and Wonorejo Village in Tulungagung District, East Java (8°05'33.8"S 111°56'15.7"E; 8°05'34.0"S 111°56'14.1"E; 8°05'33.9"S 111°56'16.4"E). The total sampling site was nine fields as three fields of each province. Symptomatic plants (stunted growth and clumped distribution) were sampled for each field, three plants per field. To obtain the DNA sequence, three individuals (one J2 per plant) were pooled of each field for analysis. The nematode population was obtained by dissecting the root for further DNA extraction.
2.2. Molecular Identification
DNA was extracted from three juveniles per rice field site with a method described by Morindya et al. [14] using a commercial kit (GeneAidTM Tissued/Blood DNA Mini Kit). The final DNA template was subjected to polymerase chain reaction (PCR) amplification as described by Htay et al. [15], with specific primers of M. graminicola Mg-F3 (5`-TTATCGCATCATTTTATTTG-3`) / Mg-R2 (5`-CGCTTTGTTAGAAAATGACCCT-3`) and an amplified target fragment of 378 bp of rDNA sequence on the 5.8S region. Exactly 10 μl of the PCR mixture (5 μl of MyTaq HS Red Mix Bioline, 1 μl of nuclease-free water, 1 μl of each forward and reverse primer at 0.2 μM concentration, and 2 μl of DNA template) was incubated on the PCR machine and run for initial denaturation at 94°C for 2 minutes. Then, 35 cycles were performed: denaturation at 94°C for 15 seconds, annealing at 50°C for 30 seconds, and extension at 68°C for 60 seconds. The final synthesis was carried out at 68°C for 5 minutes, then 4°C for the final temperature. All PCR products were then separated by electrophoresis on a 2% agarose gel and 0.1% FloroSafe DNA Stain (1st BASE FloroSafe DNA Stain BIO-5170) at 0.01% v/v at 100 V for 25 minutes and then visualized in the UV transilluminator. The amplified products were sequenced using Sanger sequencing at the Integrated Laboratory for Research and Testing, Universitas Gadjah Mada, and the sequences were submitted to GenBank to obtain the accession numbers.
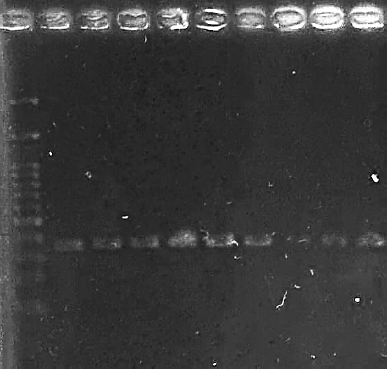 | Figure 1. Electrophoresis of PCR products amplified with specific primer of M. graminicola Mg-F3/Mg-R2. M: 1 Kb DNA marker. Lanes 1–3: West Java samples, 4–6: Central Java samples, 7–9: East Java samples. [Click here to view] |
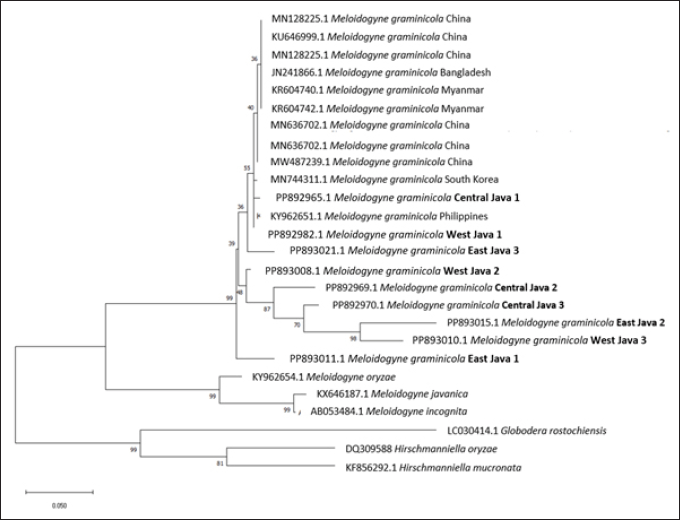 | Figure 2. Phylogenetic tree of M. graminicola population in Java. The evolutionary distance was measured using maximum likelihood method and Kimura 2-parameter with evolutionarily invariable component (K2+I) of tree model. [Click here to view] |
2.3. Data Analyses
All DNA sequences were aligned and trimmed using ClustalW on BioEdit software (version 7.2). A phylogenetic tree was constructed in the Molecular Evolutionary Genetic Analysis (MEGA) software (version 11).
3. RESULTS
Molecular detection using the specific primer Mg-F3/Mg-R2 amplified a specific band of 378 bp in nine samples of the Java population (Fig. 1). Nucleotide sequence data of this study were identified as M. graminicola, deposited on GenBank with the accession numbers for samples from West Java (PP892982.1; PP893008.1; PP893010.1), Central Java (PP892965.1; PP892969.1; PP892970.1), and East Java (PP893011.1; PP893015.1; PP893021.1).
3.1. Phylogenetic Relationship
We constructed the phylogenetic tree using the maximum likelihood method and the Kimura 2-parameter model with invariant sites (K2 + I) and performed 1,000 bootstrap replicates. The analysis used all sites, treating gaps as missing data. The phylogenetic tree of M. graminicola indicated that all isolates from Java were in the same clade as those from China, Myanmar, Bangladesh, South Korea, and the Philippines. Most isolates formed a group with the Philippines and South Korea, while those from China, Bangladesh, and Myanmar were in a separate group. Additionally, several ingroup genera with different species, Meloidogyne oryzae, Meloidogyne incognita, and Meloidogyne javanica, cluster in a different clade with M. graminicola. The other clades, Globodera rostochiensis, Hirschmanniella oryzae, and Hirschmanniella mucronata, act as an outgroup from different genera (Fig. 2).
The genetic distance analysis revealed a similar trend to that observed in the phylogenetic tree, in which smaller genetic distance values indicate closer relationships among the isolates [16]. Figure 2 shows that several isolates from Java (ID14; ID15; ID19; ID20) were closely related to those from the Philippines (ID5), with a distance ranging from 0.00 to 0.04. The lowest distance among isolates from Java and references was between the isolate from West Java 3 (ID20) and the Philippines (ID5), while the highest distance was from isolates of East Java 2 (ID16) to those of Myanmar, Bangladesh, and China (ID6-ID11) (Fig. 3).
3.2. Multiple Alignment Analysis
The sequences of all populations in Java were aligned using MEGA v.11, and the final trimmed alignment length was 378 bp. Multiplesequence alignment of M. graminicola among Java populations showed a high variation on the first 100 bp, became more identical on the next 200 bp, and varied on the last 69 bp (Fig. S1). Among these samples, there were 69 variable sites and nine distinct haplotypes. Single-nucleotide polymorphisms (SNPs) were counted by comparing all sequences to the first sequence in the alignment, which was used as the reference sequence (West Java 1). The variable sites of the M. graminicola population in West Java, Central Java, and East Java were 31, 25, and 56, respectively, with three distinct haplotypes for each population. SNPs for the population across these locations were determined by comparing all sequences in each location to the first sequence in the alignment of each location, namely West Java 1, Central Java 1, and East Java 1 (Figs. S2–4).
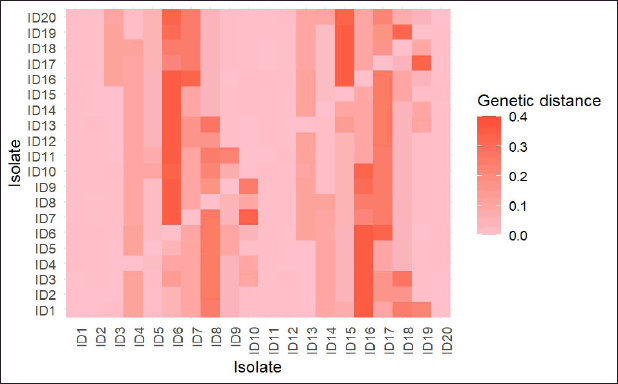 | Figure 3. Genetic distance among isolates. ID1 OP941600.1 China; ID2 MW487239.1 China; ID3 MN636702.1 China; ID4 MN744311.1 South Korea; ID5 KY962651.1 Philippines; ID6 KR604742.1 Myanmar; ID7 KR604740.1 Myanmar; ID8 JN241866.1 Bangladesh; ID9 MN521459.1 China; ID10 MN128225.1 China; ID11 KU646999.1 China; ID12 PP892969.1 Central Java 2; ID13 PP892970.1 Central Java 3; ID14 PP892965.1 Central Java 1; ID15 PP893021.1 East Java 3; ID16 PP893015.1 East Java 2; ID17 PP893011.1 East Java 1; ID18 PP893010.1 West Java 3; ID19 PP893008.1 West Java 2; ID20 PP892982.1 West Java 1. [Click here to view] |
4. DISCUSSION
Meloidogyne graminicola is among the most destructive plantparasitic nematodes of rice and has been reported in Indonesia, including the sampled Java locations. where the infestation is reported in sampled locations from west to east Java. This study confirmed the rapid detection using species-specific primers for M. graminicola. A previous study on RKN detection infesting onion samples using the same primer as this study also showed the same target band for M. graminicola detection [17]. Phylogenetic analysis revealed that nine samples of nematodes from rice samples in Java were grouped with M. graminicola reference sequences with the highest support branch (99%). It shows a significant difference from M. oryzae as the same primary host on rice, as well as other prevalent rice RKNs, such as M. incognita and M. javanica, on different host species [18–20]. A recent study used a novel method that enables the detection of M. graminicola and screening of M. oryzae in the same population [21].
The global prevalence of M. graminicola on rice-producing countries is evident in Asia, including Bangladesh, Cambodia, China, India, Indonesia, Laos, Malaysia, Myanmar, Nepal, Pakistan, Philippines, Singapore, Sri Lanka, Thailand, and Vietnam [22]. Based on the genetic distance, M. graminicola of several Indonesian isolates were closely related to the Philippines, in which M. graminicola is the major threat to rice production and has been molecularly detected by ribosomal and mitochondrial DNA genes [23] and on weedy rice roots using the Sequence Characterized Amplified Region marker [24].
This study revealed a nucleotide variation of the DNA sequence within the M. graminicola population in Java on the small locus of the ribosomal gene (Figs. S1–4). Although this study found large SNPs in M. graminicola population in Java, we cannot claim that this population is diverse because the sample size determined by the small specific rDNA gene was small, namely, less than 1% of the total M. graminicola genome, approximately 41.5 Mb [25]. This study reported multiprovince molecular confirmation in Java and new sequences deposited. Despite the limitation on the number of samples due to undetected symptoms in the field, this study showed an updated distribution of M. graminicola infection, particularly in Java, Indonesia. Future studies can probe into M. graminicola populations in Java using whole genome sequencing to evaluate the genetic variation between populations.
5. CONCLUSION
Rice RKN infestation in Java has been detected in sampled locations in West Java, Central Java, and East Java, and molecularly identified as M. graminicola. Based on DNA sequences on short loci of ribosomal genes, high nucleotide variations were demonstrated across the population. Further study was required to assess the genetic variation of M. graminicola in Java.
6. ACKNOWLEDGMENTS
The authors acknowledge the financial support provided by Universitas Gadjah Mada (UGM) through the Final Assignment Recognition under contract No. 5286/UN1.P1/PT.01.03/2024. We also gratefully acknowledge Rendyta Morindya, Nada Ayu Nurlaili, and Muhammad Iqbal Maulana for assisting with the analysis, also Ms. Rebecca Santi for the language editing and proofreading services.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. CONFLICT OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. DISCLOSURE
A preprint version of this manuscript is available at Research Square https://www.researchsquare.com/article/rs-5629720/v1.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
14. SUPPLEMENTARY MATERIAL
The supplementary material can be accessed at the journal’s website: https://jabonline.in/admin/php/uploadss/1482_pdf.pdf
REFERENCES
1. Netscher C, Erlan E. A root-knot nematode, Meloidogyne cf graminicola, parasitic on rice in Indonesia. Afro-Asian J Nematol 1993;3(1):90–5.
2. Rusinque L, Maleita C, Abrantes I, Palomares-Rius JE, Inacio M. Meloidogyne graminicola-a threat to rice production: review update on distribution, biology, identification, and management. Biol (Basel) 2021;10(1163):1–9. CrossRef
3. Tian ZL, Maria M, Barsalote EM, Castillo P, Zheng JW. Morphological and molecular characterization of the rice root-knot nematode, Meloidogyne graminicola, Golden and Birchfeild, 1965 occurring in Zhejiang, China. J Integr Agric 2018;17(12):2724–33. CrossRef
4. Jabbar A, Javed N, Munir A, Abbas H, Khan SA, Moosa A, et al. Occurrence and molecular characterization of Meloidogyne graminicola on rice in Central Punjab, Pakistan. J Nematol 2021;52:1–1. CrossRef
5. Khan MR, Ahamad F. Incidence of root-knot nematode (Meloidogyne graminicola) and resulting crop losses in paddy rice in Northern India. Plant Dis 2020;104(1):186–93. CrossRef
6. Hodda M. Phylum Nematoda: a classification, catalogue and index of valid genera, with a census of valid species. Zootaxa 2022;5114(1):1–289. CrossRef
7. Zhou S, Yan B, Li F, Zhang J, Zhang J, Ma H, et al. RNA-Seq analysis provides the first insights into the phylogenetic relationship and interspecific variation between Agropyron cristatum and wheat. Front Plant Sci 2017;8:1–3. CrossRef
8. Shao H, Liu MY, Huang WK, Li CH, Peng D, Peng H, et al. Genetic diversity of the root-knot nematode Meloidogyne enterolobii in mulberry based on the mitochondrial COI gene. Ecol Evol 2020;10:5391–401. CrossRef
9. Bik HM, Fournier D, Sung W, Bergeron RD, Thomas WK. Intragenomic variation in the ribosomal repeats of nematodes. PLoS One 2013;8(10):1–8. CrossRef
10. Floyd RM, Rogers AD, Lambshead PJD, Smith CR. Nematodespecific PCR primers for the 18S small subunit rRNA gene. Mol Ecol Notes 2005;5(3):611–2. CrossRef
11. Valadas V, Laranjo M, Mota M, Oliveira S. A survey of entomopathogenic nematode species in continental Portugal. J Helminthol 2013;88(3):1–5. CrossRef
12. Nurjayadi MY, Munif A, Suastika G. Identifikasi Nematoda Puru Akar, Meloidogyne graminicola, pada Tanaman Padi di Jawa Barat. J Fitopatol Indones 2015;11(4):113–20. CrossRef
13. Mirsam H, Kurniawati F. Laporan Pertama di Sulawesi Selatan: karakter Morfologi dan Molekuler Nematoda Puru Akar yang Berasosiasi dengan Akar Padi di Kabupaten Wajo, Sulawesi Selatan. J Perlindungan Tanam Indones 2018;22(1):58–65. CrossRef
14. Morindya R, Indarti S, Soffan A, Hartono S. Optimization of DNA extraction methods for genomic analysis of rice root-knot nematode (Meloidogyne graminicola) using PCR (polymerase chain reaction) and sanger sequencing. J Plant Prot Res 2023;63(1):50–8.
15. Htay C, Peng H, Huang W, Kong L, He W, Holgado R, et al. The development and molecular characterization of a rapid detection method for rice root-knot nematode (Meloidogyne graminicola). Eur J Plant Pathol 2016;146(2):281–91. CrossRef
16. Trisyani N, Rahayu DA. DNA barcoding of razor clam Solen spp. (Solenidae, Bivalvia) in Indonesian beaches. Biodiversitas 2020;21(2):478–84. CrossRef
17. Li Q, Yang Y, Liu F, Li Y, Yao H, Peng D, et al. Morphological and molecular characterization and life cycle of Meloidogyne graminicola infecting Allium cepa. Agronomy 2025;15(1994):1–5. CrossRef
18. Hamidi I, Mutaqin KH, Kurniawati F. Spesies Meloidogyne penyebab ubi kentang berbintil pada tiga sentra produksi di Sumatra Meloidogyne species cause pimple-like swelling on potato tuber in three production centers in Sumatra. J Fitopatol Indones 2022;18(2):66–74. CrossRef
19. Hikmia Z, Suastika G. Identifikasi Spesies Meloidogyne spp. penyebab umbi bercabang pada tanaman wortel di Jawa Timur Species identification of Meloidogyne spp., the cause of branched tuber disease in carrot in East Java. J Fitopatol Indones 2012;8(3):73–8. CrossRef
20. Mutala’Liah M, Indarti S, Wibowo A. Short communication: the prevalence and species of root-knot nematode which infect on potato seed in Central Java, Indonesia. Biodiversitas J Biol Divers 2019;20(1):11–6. CrossRef
21. Rusinque L, Pasupuleti S, Maleita C, Inacio M, Somvanshi V, De Andrade E. Rapid and sensitive detection of the rice root-knot nematode Meloidogyne graminicola and screening of M. oryzae using recombinase polymerase amplification. Physiol Mol Plant Pathol 2025;139:1–9. CrossRef
22. EPPO. EPPO datasheet: meloidogyne graminicola. Hoboken, NJ: John Wiley & Sons, Inc; 2025.
23. Cabasan MTN, Kumar A, Bellafiore S, De Waele D. Reproductive, pathogenic and genotypic characterisation of five Meloidogyne graminicola populations from the Philippines on susceptible and resistant rice varieties. Nematology 2018;20(4):299–318. CrossRef
24. Latina RA, Tabernilla CHR, Ramirez AHM, Latina RA, Tabernilla CHR, Henedina A, et al. Detection of root-knot nematodes infecting weedy rice in San Juan, Batangas, Philippines. Arch Phytopathol Plant Prot 2023;56(9):753–9. CrossRef
25. Phan NT, Orjuela J, Danchin EGJ, Klopp C, Perfus-Barbeoch L, Kozlowski DK, et al. Genome structure and content of the rice root-knot nematode (Meloidogyne graminicola). Ecol Evol 2020;10(20):1–16. CrossRef