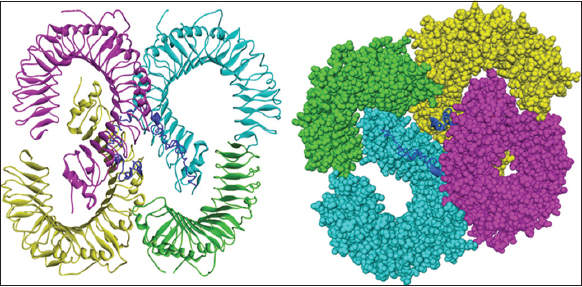
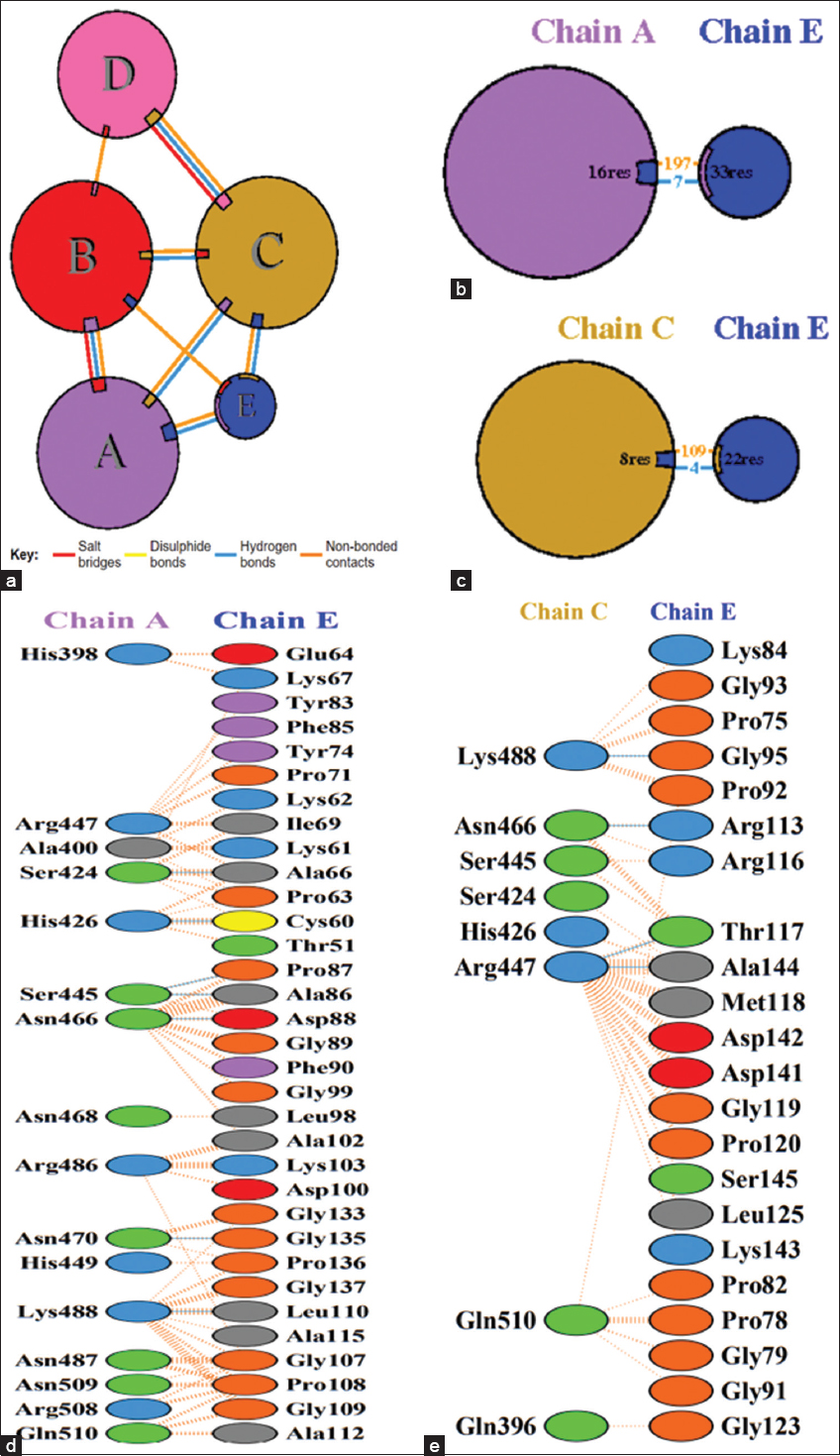
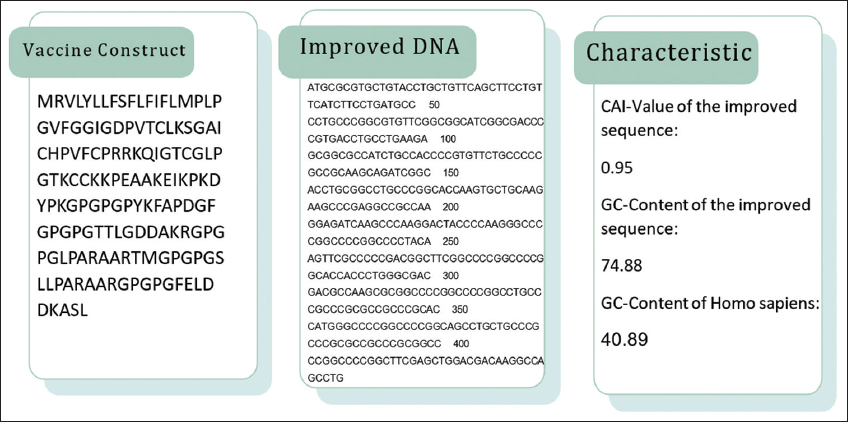
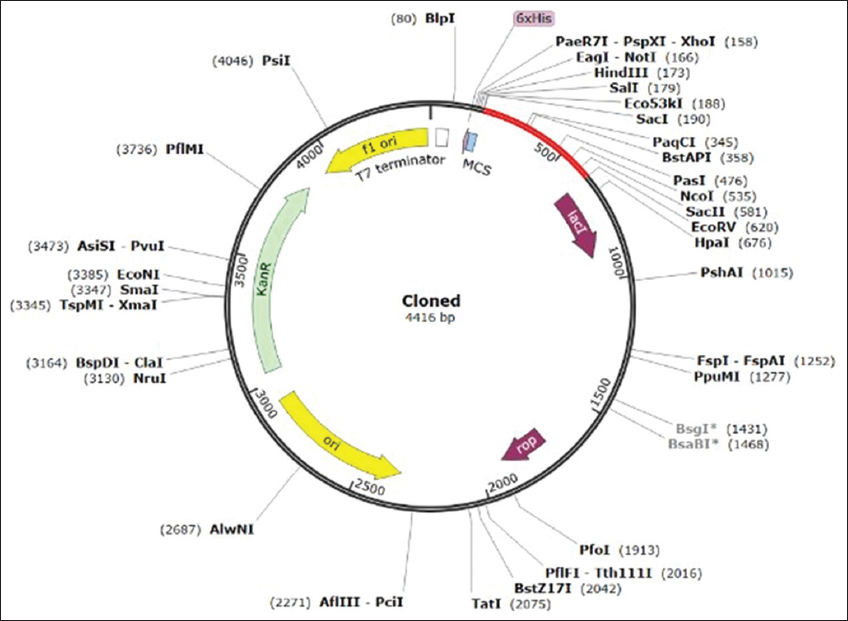
1. Lawrence R, Jeyakumar E. Antimicrobial resistance:A cause for global concern. BMC Proc 2013;7:S1. [CrossRef]
2. McEwen SA, Fedorka-Cray PJ. Antimicrobial use and resistance in animals. Clin Infect Dis 2002;34 Suppl 3:S93-106. [CrossRef]
3. Song Y, Wang LF, Zhou K, Liu S, Guo L, Ye LY, et al. Epidemiological characteristics, virulence potential, antimicrobial resistance profiles, and phylogenetic analysis of Aeromonas caviae isolated from extra-intestinal infections. Front Cell Infect Microbiol 2023;13:1084352. [CrossRef]
4. Batra P, Mathur P, Misra MC. Aeromonas spp.:An emerging nosocomial pathogen. J Lab Physicians 2016;8:1-4. [CrossRef]
5. Pereira C, Duarte J, Costa P, Braz M, Almeida A. Bacteriophages in the control of Aeromonas sp. in aquaculture systems:An integrative view. Antibiotics (Basel) 2022;11:163. [CrossRef]
6. Stabler RA, Marsden GL, Witney AA, Li Y, Bentley SD, Tang CM, et al. Identification of pathogen-specific genes through microarray analysis of pathogenic and commensal Neisseria species. Microbiology (Reading) 2005;151:2907-22. [CrossRef]
7. Solanki V, Tiwari M, Tiwari V. Prioritization of potential vaccine targets using comparative proteomics and designing of the chimeric multi-epitope vaccine against Pseudomonas aeruginosa. Sci Rep 2019;9:5240. [CrossRef]
8. Rahmani A, Baee M, Saleki K, Moradi S, Nouri HR. Applying high throughput and comprehensive immunoinformatics approaches to design a trivalent subunit vaccine for induction of immune response against emerging human coronaviruses SARS-CoV, MERS-CoV and SARS-CoV-2. J Biomol Struct Dyn 2022;40:6097-113. [CrossRef]
9. Alcock BP, Raphenya AR, Lau TT, Tsang KK, Bouchard M, Edalatmand A, et al. CARD 2020:Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res 2020;48:D517-25. [CrossRef]
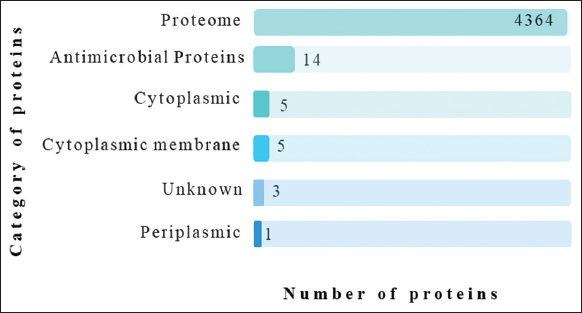
10. Yu NY, Wagner JR, Laird MR, Melli G, Rey S, Lo R, et al. PSORTb 3.0:Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010;26:1608-15. [CrossRef]
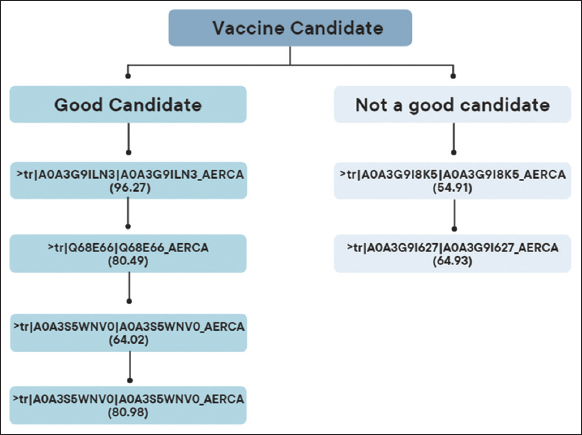
11. Rawal K, Sinha R, Nath SK, Preeti P, Kumari P, Gupta S, et al. Vaxi DL:A web-based deep learning server to identify potential vaccine candidates. Comput Biol Med 2022;145:105401. [CrossRef]
12. Galanis KA, Nastou KC, Papandreou NC, Petichakis GN, Pigis DG, Iconomidou VA. Linear B-cell epitope prediction for in silico vaccine design:A performance review of methods available via command-line interface. Int J Mol Sci 2021;22:3210. [CrossRef]
13. Janahi EM, Dhasmana A, Srivastava V, Sarangi AN, Raza S, Arif JM, et al. In silico CD4+, CD8+T-cell and B-cell immunity associated immunogenic epitope prediction and HLA distribution analysis of Zika virus. EXCLI J 2017;16:63-72.
14. Ong E, He Y, Yang Z. Epitope promiscuity and population coverage of Mycobacterium tuberculosis protein antigens in current subunit vaccines under development. Infect Genet Evol 2020;80:104186. [CrossRef]
15. Karkashan A. Immunoinformatics assisted profiling of west Nile virus proteome to determine immunodominant epitopes for the development of next-generation multi-peptide vaccine. Front Immunol 2024;15:1395870. [CrossRef]
16. Doytchinova IA, Flower DR. VaxiJen:A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinformatics 2007;8:4. [CrossRef]
17. Dimitrov I, Flower DR, Doytchinova I. AllerTOP - a server for in silico prediction of allergens. BMC Bioinformatics 2013;14 Suppl 6:S4. [CrossRef]
18. Wei L, Ye X, Sakurai T, Mu Z, Wei L. ToxIBTL:Prediction of peptide toxicity based on information bottleneck and transfer learning. Bioinformatics 2022;38:1514-24. [CrossRef]
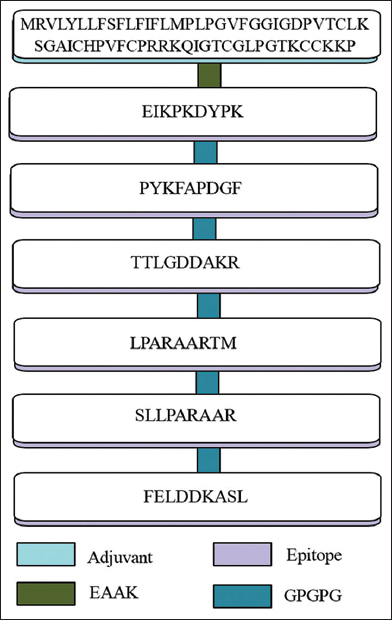
19. Lei Y, Zhao F, Shao J, Li Y, Li S, Chang H, et al. Application of built-in adjuvants for epitope-based vaccines. PeerJ 2019;6:e6185. [CrossRef]
20. Cie?lik M, Bagi?ska N, Górski A, Jo?czyk-Matysiak E. Human b-defensin 2 and Its postulated role in modulation of the immune response. Cells 2021;10:2991. [CrossRef]
21. Nguyen TL, Kim H. Immunoinformatics and computational approaches driven designing a novel vaccine candidate against Powassan virus. Sci Rep 2024;14:5999. [CrossRef]
22. Shams M, Nourmohammadi H, Majidiani H, Shariatzadeh SA, Asghari A, Fatollahzadeh M, et al. Engineering a multi-epitope vaccine candidate against Leishmania infantum using comprehensive immunoinformatics methods. Biologia (Bratisl) 2022;77:277-89. [CrossRef]
23. Ramalingam PS, Arumugam S. Reverse vaccinology and immunoinformatics approaches to design multi-epitope based vaccine against oncogenic KRAS. Med Oncol 2023;40:283. [CrossRef]
24. Buchan DW, Jones DT. The PSIPRED protein analysis workbench:20 years on. Nucleic Acids Res 2019;47:W402-7. [CrossRef]
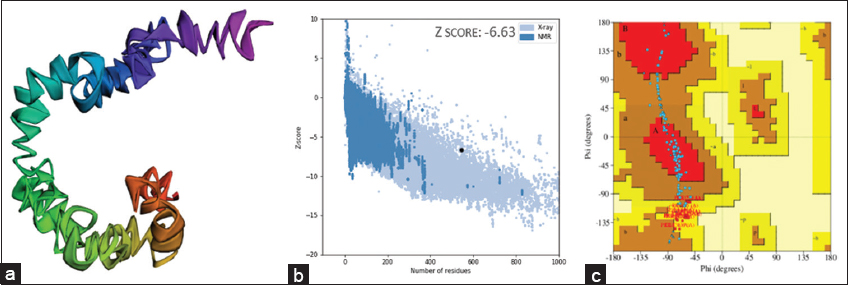
25. Prajapat R, Marwal A, Gaur RK. Recognition of errors in the refinement and validation of three-dimensional structures of AC1 proteins of Begomovirus strains by using ProSA-web. J Viruses 2014;2014:752656. [CrossRef]
26. Laskowski RA, MacArthur MW, Moss DS, Thornton JM. Procheck:A program to check the stereochemical quality of protein structures. J Appl Crystallogr 1993;26:283-91. [CrossRef]
27. DeVore GR. Computing the Z score and centiles for cross-sectional analysis:A practical approach. J Ultrasound Med 2017;36:459-73. [CrossRef]
28. Yan Y, Zhang D, Zhou P, Li B, Huang SY. HDOCK:A web server for protein-protein and protein-DNA/RNA docking based on a hybrid strategy. Nucleic Acids Res 2017;45:W365-73. [CrossRef]
29. Gupta AK, Khan MS, Choudhury S, Mukhopadhyay A, Sakshi, Rastogi A, et al. CoronaVR:A computational resource and analysis of epitopes and therapeutics for severe acute respiratory syndrome coronavirus-2. Front Microbiol 2020;11:1858. [CrossRef]
30. Laskowski RA. PDBsum:Summaries and analyses of PDB structures. Nucleic Acids Res 2001;29:221-2. [CrossRef]
31. Grote A, Hiller K, Scheer M, Münch R, Nörtemann B, Hempel DC, et al. JCat:A novel tool to adapt codon usage of a target gene to its potential expression host. Nucleic Acids Res 2005;33:W526-31. [CrossRef]
32. Naas AE. Optimzing the 5'-End of Coding Sequences in Recombinant mRNA to Achieve High-Level Expression in the Bacterium Escherichia coli. Trondheim:Institutt for Bioteknologi;2012.
33. Chatterjee R, Sahoo P, Mahapatra SR, Dey J, Ghosh M, Kushwaha GS, et al. Development of a conserved chimeric vaccine for induction of strong immune response against Staphylococcus aureus using immunoinformatics approaches. Vaccines (Basel) 2021;9:1038. [CrossRef]
34. Wareth G, Brandt C, Sprague LD, Neubauer H, Pletz MW. WGS based analysis of acquired antimicrobial resistance in human and non-human Acinetobacter baumannii isolates from a German perspective. BMC Microbiol 2021;21:210. [CrossRef]
35. Sabikhi Y, Surabhi, Dixit P, Sharma T, Sharma P, Rai M, et al. N-Acetylmuramoyl-L-Alanine Amidase:A Competent Vaccine Candidate against. United States:IPSID;2022. [CrossRef]
36. Alzarea SI. Identification and construction of a multi-epitopes vaccine design against Klebsiella aerogenes:Molecular modeling study. Sci Rep 2022;12:14402. [CrossRef]
37. Kobayashi H, Wood M, Song Y, Appella E, Celis E. Defining promiscuous MHC class II helper T-cell epitopes for the HER2/neu tumor antigen. Cancer Res 2000;60:5228-36.
38. Aldakheel FM, Abrar A, Munir S, Aslam S, Allemailem KS, Khurshid M, et al. Proteome-wide mapping and reverse vaccinology approaches to design a multi-epitope vaccine against Clostridium perfringens. Vaccines (Basel) 2021;9:1079. [CrossRef]
39. Khan S, Irfan M, Hameed AR, Ullah A, Abideen SA, Ahmad S, et al. Vaccinomics to design a multi-epitope-based vaccine against monkeypox virus using surface-associated proteins. J Biomol Struct Dyn 2023;41:10859-68. [CrossRef]
40. Asif Rasheed M, Awais M, Aldhahrani A, Althobaiti F, Alhazmi A, Sattar S, et al. Designing a highly immunogenic multi epitope based subunit vaccine against Bacillus cereus. Saudi J Biol Sci 2021;28:4859-66. [CrossRef]
41. Naveed M, Hassan JU, Ahmad M, Naeem N, Mughal MS, Rabaan AA, et al. Designing mRNA- and peptide-based vaccine construct against emerging multidrug-resistant Citrobacter freundii:A computational-based subtractive proteomics approach. Medicina (Kaunas) 2022;58:1356. [CrossRef]
42. Alawam AS, Alwethaynani MS. Construction of an aerolysin-based multi-epitope vaccine against Aeromonas hydrophila:An in silico machine learning and artificial intelligence-supported approach. Front Immunol 2024;15:1369890. [CrossRef]
43. Dey J, Mahapatra SR, Patnaik S, Lata S, Kushwaha GS, Panda RK, et al. Molecular characterization and designing of a novel multiepitope vaccine construct against Pseudomonas aeruginosa. Int J Pept Res Ther 2022;28:49. [CrossRef]
44. Basak S, Deb D, Narsaria U, Kar T, Castiglione F, Sanyal I, et al. In silico designing of vaccine candidate against Clostridium difficile. Sci Rep 2021;11:14215. [CrossRef]
45. Kumar P, Kumar P, Shrivastava A, Dar MA, Lokhande KB, Singh N, et al. Immunoinformatics-based multi-epitope containing fused polypeptide vaccine design against visceral leishmaniasis with high immunogenicity and TLR binding. Int J Biol Macromol 2023;253:127567. [CrossRef]
46. Tian J, Yan Y, Yue Q, Liu X, Chu X, Wu N, et al. Predicting synonymous codon usage and optimizing the heterologous gene for expression in E. coli. Sci Rep 2017;7:9926. [CrossRef]
47. Basmenj ER, Arastonejad M, Mamizadeh M, Alem M, KhalatbariLimaki M, Ghiabi S, et al. Engineering and design of promising T-cell-based multi-epitope vaccine candidates against leishmaniasis. Sci Rep 2023;13:19421. [CrossRef]
48. Zheng B, Suleman M, Zafar Z, Ali SS, Nasir SN, Namra, et al. Towards an ensemble vaccine against the pegivirus using computational modelling approaches and its validation through in silico cloning and immune simulation. Vaccines (Basel) 2021;9:818. [CrossRef]
49. Furman D, Davis MM. New approaches to understanding the immune response to vaccination and infection. Vaccine 2015;33:5271-81. [CrossRef]