
1. INTRODUCTION
Guppies (Poecilia reticulata) are ornamental fish. They have a high commercial value. This is true in both domestic and international markets. Male guppies exhibit more attractive morphology than females, including body shape, body color patterns, and fins [1-5]. Therefore, male guppies are often raised with the monoculture method, due to more profitable than female guppies [1,2,6,7]. Increasing the opportunity to produce male P. reticulata can be achieved through the application of sex reversal techniques [5,8].
The process of sex reversal in fish can be achieved through two distinct mechanisms: Masculinization and feminization. Masculinization generates male mono-sex fish [3,9]. Synthetic hormones are often required to implement the sex reversal techniques [10]. For masculinization process, the synthetic hormones utilized are 17α-methyltestosterone (17α-MT), androstenedione, 19-norethynyl testosterone, 17α-ethyl testosterone, 17α-methyl-dihydrotestosterone, and dihydrotestosterone [2,3,11,12]. The 17α-MT is the most commonly-used hormone for fish masculinization [9]. However, the synthetic hormone 17α-MT is classified as a hard drug and prohibited for further usage in fish culture activities in Indonesia [13]. Consequently, it is necessary to select other alternative substances for synthetic hormones. An alternative material that has been found to help increase the number of male fish in the masculinization process is the purwoceng (Pimpinella alpina) [14-19].
Purwoceng (P. alpina) is an indigenous plant of Indonesia [20-23]. This plant belongs to Pimpinella genus that contains phytosterols, including α-spinasterol, campesterol, stigmasta-5,7,2-trien-3-ol, Δ7-avenasterol, and Δ5-avenasterol. In another study, P. anisum contains campesterol, stigmasterol (stigmasta-5,22-dien-3β-ol), β-sitosterol (stigmast-5-en-3β-ol), and α-spinasterol [24]. Phytosterols have been demonstrated to affect endocrinal reproductive functions in fish [25-27]. Phytosterol (β-sitosterol) has been shown to reduce low-density lipoprotein cholesterol and triglyceride levels in trout (Salvelinus fontinalis) [28]. Consequently, β-sitosterol inhibits the synthesis of estrogen hormones, while stimulating the synthesis of androgen hormones. Androgen hormones influence the differentiation of sex toward the male phenotype.
Studies on the use of P. alpina for fish masculinization have been conducted on Betta fish (Betta splendens) [15], rainbowfish (Iriatherina werneri) [14], Synodontis catfish (Synodontis sp.) [16], and Nile tilapia (Oreochromis niloticus) [19]. These studies indicate that the P. alpina can be employed as an alternative material to synthetic hormones in the male monosex fish production. The immersion of Nile tilapia larvae in P. alpina extract for 10 h obtained 79.22% male fish [19]. The immersion method on pregnant females of the P. reticulata in P. alpina extract for 8 h produced 63.98% of male offspring [17]. Nevertheless, the impact of the immersion method on P. reticulata larvae in P. alpina extract has yet to be investigated. Therefore, this study determined the effect of the immersion duration on the success of P. reticulata masculinization with P. alpina extract. The findings of this study are anticipated to be instrumental in the field of sex reversal in fish, with a focus on the utilization of naturally occurring compounds, such as phytosterols, as a substitute for synthetic hormones.
2. MATERIALS AND METHODS
2.1. Period and Location
This study was conducted at the Laboratory of Fisheries, Laboratory of Aquaculture, Laboratory of Microbiology and Biotechnology for Fishery Products, Department of Fisheries, Faculty of Agriculture, Universitas Sriwijaya, and the Batanghari-9 Indralaya Breeding Unit from October to December, 2023.
2.2. Fish Spawning
This study used 24 P. reticulata broodstocks (12 males and 12 females, with a 1:1 sex ratio) from the black-Moscow strain at 3–4 months old with a total length of 3–4 cm. Broodstocks were fed with Tubifex sp. and Daphnia sp. at 2–3 times/day. The mass-spawning was conducted in a box container (50 × 30 × 32 cm3). After the mating period for 14 days, the larvae were produced. The larvae were subsequently collected in different box containers (49 × 33 × 27 cm3) for further treatments.
2.3. P. alpina Extraction
The 100 g of P. alpina powder were dissolved in 1 L of ethanol (70%) and macerated for 24 h, equipped with stirring every 2 h. After 24 h, the solution was filtered using a filter bag with a pore size of 200 μm. The filtrate was subsequently stored in an Erlenmeyer flask and evaporated using a rotary evaporator at 60 rpm in 48°C for 3 h and 30 min. This process produced a liquid P. alpina extract, which was ready for further treatments.
2.4. Immersion and Rearing
Larvae were immersed in 12 different jars with a volume of 1.5 L. Each jar was filled with 1 L of water. Before immersion, 0.2 mg/L of P. alpina extract was mixed with the water and aerated for 24 h. The concentration of P. alpina extract in the present study was determined on the basis of the findings of previous research. The present study utilized a total of 120 P. reticula larvae. A total of 10 larvae were subjected to a 24-h immersion period in each jar, with the immersion duration applied for 8, 10, and 12 h. A group of larvae was not subjected to the P. alpina solution and served as the control treatment. Subsequently, the larvae were transferred to 12 distinct jars (5 L). The P. reticulata larvae were reared for 45 days and fed with Daphnia sp. and Artemia cysts (Polar Red Artemia®) at ad libitum condition. The survival rate was calculated from the duration of the immersion and rearing periods, namely:
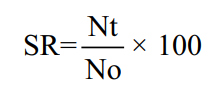
Where:
SR: Survival rate (%)
Nt: Number of fish at the final treatment and rearing (fish)
No: Number of fish at the initial treatment and rearing (fish)
2.5. Sex Determination
This study has been approved by the Dean of the Faculty of Agriculture, Sriwijaya University, Number 4612/UN9.1.5/AK.16/2022. The sex determination was conducted through the surgical method. The fish were dissected to obtain their gonads using forceps. Subsequently, the gonads were placed on an object glass and minced with a scalpel. Two drops of acetocarmine solution were added and allowed to remain for a minute. The object glass was then covered with a cover glass. The gonads were observed under a binocular microscope (Olympus CX 23®) at 40× magnification. The male fish gonads were identified by the presence of developing sperm cells. The percentage of male fish was calculated using the following formula:
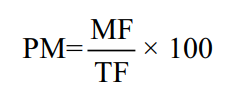
Where:
PM: Percentage of male fish (%)
MF: Number of male fish at the initial rearing period (fish)
TF: Total number of fish at the final rearing period (fish)
2.6. Water Quality
The water quality parameters were measured using the following instruments: A pH meter, a thermometer, a DO meter, and a spectrophotometer. The parameters included pH, water temperature, dissolved oxygen, and ammonia. The temperature and pH were recorded on a daily basis, while the dissolved oxygen and ammonia levels were monitored on a weekly basis.
2.7. Data Analysis
All data were tabulated and calculated using the Microsoft Excel 2019. Subsequently, a statistical analysis was conducted using the analysis of variance with the assistance of the Statistical Package for the Social Sciences 20.0 version. The percentage of male fish and survival rate data were presented in bar charts, while the water quality data were presented in tabular form and analyzed descriptively.
3. RESULTS AND DISCUSSION
3.1. Percentage of Male Fish
The present study indicates that the immersion duration with P. alpina extract has a significant effect (P < 0.05) on the percentage of male fish. In the 0-h treatment, the percentage of males was 43.33%. In the 8-h treatment, the percentage increased by 70.00%. In the 10-h treatment, the percentage elevated to 83.33%. Yet, in the 12-h treatment, the percentage reduced to 73.33%. The 0-h treatment exhibited a significant difference (P < 0.05) from all other treatments. The 8-h treatment exhibited a statistically significant difference (P < 0.05) from the 10-h treatment. Meanwhile, the 12-h treatment had no statistically significant difference (P > 0.05) from the 8- and 12-h treatments [Figure 1].
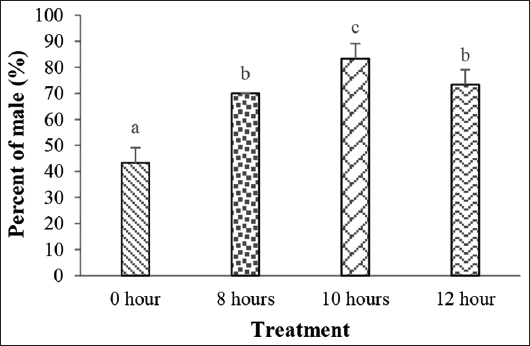 | Figure 1: Percentage of male guppy fish (Poecilia reticulata) after immersion in purwoceng (Pimpinella alpina) extract. Different letters indicate significant differences between treatments (P < 0.05). [Click here to view] |
This study yielded higher results from a previous study that reported a male fish percentage of 63.98% [17]. This condition has occurred as thought to be due to the presence of phytosterol compounds in P. alpina, which are known to influence gonadal differentiation toward males. The phytosterol contents in P. anisum are 551.9 mg/100 g sample [24]. In addition, the level of stigmasterol compounds in 10 mg of P. alpina extract is 0.538 mg [18]. Hypothetically, this condition occurred as phytosterol compounds may result in a reduction of low-density lipoprotein cholesterol and triglycerides in P. reticulata. Consequently, phytosterol compounds inhibit estrogenic hormone synthesis, while stimulating androgen hormone synthesis. The success of sex reversal is influenced by several factors, including the fish species of fish, treatment initiation time [29], the type of hormone and hormone dosage [5], treatment duration [30], and environmental factors (water temperature) [31].
Sex determination serves as the primary regulatory mechanism for the initial determination of bipotential gonads that initiate differentiation pathways [32]. The differentiation of fish sex is influenced by numerous factors, including hormonal systems and environmental factors. Environmental factors exert a direct influence on biochemical pathways that regulate the differentiation and maturation of gonads [33]. Gonadal differentiation in P. reticulata can occur up to the larval stage [34]. The vulnerability period in the poeciliids is limited to 30-day post-birth, while in P. reticulata, it encompasses both the embryonic and post-birth periods [35,36].
3.2. Survival Rate
The immersion of P. reticulata larvae in P. alpina extract had no significant effect on their survival rate during the immersion and post-immersion periods. During the immersion period, the survival rate of the larvae was 100% in all treatments [Figure 2]. The survival rate data of P. reticulata during the rearing period are presented in Figure 3.
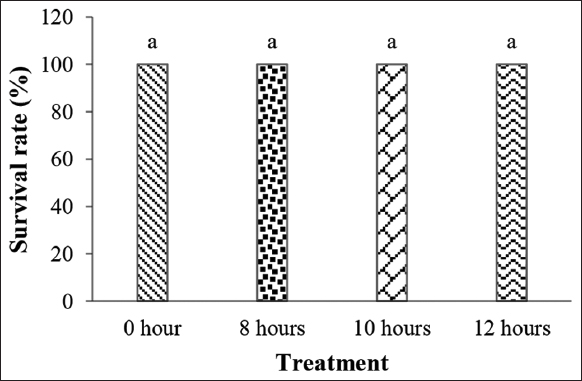 | Figure 2: Survival of guppy fish (Poecilia reticulata) larvae during immersion in purwoceng (Pimpinella alpina) extract. The same letters indicate non-significant differences between treatments (P > 0.05). [Click here to view] |
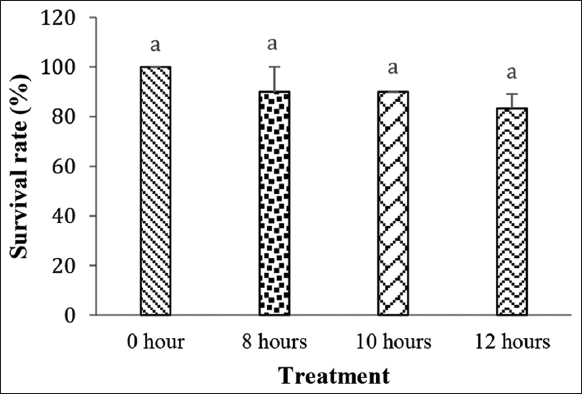 | Figure 3: Survival of guppy fish (Poecilia reticulata) during a 45-day rearing period. The same letters indicate non-significant differences between treatments (P > 0.05). [Click here to view] |
The findings of the study indicate that immersing P. reticulata larvae in P. alpina extract for 8–12 h at a dosage of 0.02 mL obtained a zero mortality level among the P. reticulata larvae. This condition was occurred as P. alpina contains limonene and γ-himachalene, which have been demonstrated to enhance the immune system of P.reticulata [23].
The survival rate of P. reticulata after 45 days of rearing was 100% in the 0-h treatment, 90.00% (8 h), 90.00% (10 h), and 83.33% (12 h). These results present a declining trend in the survival rate of guppy fish. This decline is presumed to be due to inappropriate feeding and water quality exchange from the immersion medium to the rearing environment. In addition, high levels of ammonia are toxic to fish and pH levels that are too low or too high can cause fish tissue damage [37]. The survival rate of the fish larvae is highly influenced by feeding [38], whereas initial feeding stage is particularly susceptible to suboptimal conditions [39].
3.3. Water Quality
The water quality parameters during the immersion period and larval rearing period are presented in Tables 1 and 2, respectively.
Table 1: Water quality in the guppy (Poecilia reticulata) larval immersion media.
| Treatment | Water quality parameters | |||
|---|---|---|---|---|
| pH (unit) | Temperature (°C) | DO (mg/L) | Ammonia (mg/L) | |
| 0 h | 7.6–6.8 | 24.7–27.0 | 4.4–4.7 | 0.028–0.091 |
| 8 h | 7.7–6.5 | 26.9–27.0 | 4.6–4.9 | 0.026–0.052 |
| 10 h | 7.7–6.4 | 26.8–27.0 | 4.4–4.7 | 0.024–0.060 |
| 12 h | 7.6–6.5 | 26.9–27.0 | 4.4–4.6 | 0.038–0.060 |
Table 2: Water quality in the guppy fish (Poecilia reticulata) rearing media.
| Treatment | Water quality parameters | |||
|---|---|---|---|---|
| pH (unit) | Temperature (°C) | DO (mg/L) | Ammonia (mg/L) | |
| 0 h | 7.0–7.8 | 24.4–27.5 | 4.4–4.6 | 0.024–0.060 |
| 8 h | 7.0–7.8 | 24.3–27.6 | 4.6–4.9 | 0.06–0.072 |
| 10 h | 7.2–7.8 | 24.4–27.7 | 4.4–4.8 | 0.030–0.072 |
| 12 h | 7.1–7.8 | 24.4–27.8 | 4.4–4.6 | 0.030–0.091 |
The results showed that the P. alpina extract impacted the water’s pH and ammonia levels. Before the addition of P. alpina extract to the immersion media, the pH was among 7.6–7.7, while the ammonia levels were among 0.024–0.038 mg/L. After mixing the immersion water with P. alpina extract, the pH level decreased from 6.4 to 6.8, while ammonia levels increased by 0.052–0.091 mg/L. Despite the observed changes in pH and ammonia levels, they remain within the established safe limits for P. reticulata larval rearing. The optimal water quality for guppy rearing is pH of 7.0–8.3 and temperature of 24–32°C [40]. In this study, the temperature was 24.3–27.8°C, while the pH was 7.0–7.8. The dissolved oxygen levels were among 3–6.8 mg/L, ammonia levels were 0.01–0.09 mg/L, and temperature was 20.1–28.9°C [41]. The pH of the rearing media exhibited at 7.6–7.8. The optimal pH for fish rearing is between 6.5 and 9.0 [42].
4. CONCLUSION
The immersion duration of P. reticulata larvae in P. alpina extract had no significant effect on fish survival rate. However, a significant effect has occurred on the percentage of male fish. For an effective treatment, an immersion duration of 10 h obtained the highest percentage of male fish at 83.33%. The findings of this research are expected an assist for reversal techniques, as they demonstrate the potential for the use of natural substances as alternatives to synthetic hormones.
5. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
6. FUNDING
There is no funding to report.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVAL
Ethical approvals details are given in the ‘Materials and Methods’ Section.
9. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
10. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
11. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI..
REFERENCES
1. Chakraborty SB, Molnár T, Hancz C. Effects of methyltestosterone, tamoxifen, genistein and Basella alba extract on masculinization of guppy (Poecilia reticulata). J Appl Pharm Sci. 2012;2(12):48-52. [CrossRef]
2. Turan F, Çek S, Atik E. Production of monosex male guppy, Poecilia reticulata, by 17a-methyltestosterone. Aquacu Res. 2006;37:200-3. [CrossRef]
3. Golan M, Sivan LB. Artificial masculinization in tilapia involves androgen receptor activation. Gen Comp Endocrinol. 2014;207:50-5. [CrossRef]
4. Kavumpurath S, Pandian TJ. Production of a YY female guppy, Poecilia reticulata, by endocrine sex reversal and progeny testing. Aquaculture. 1993;118(3-4):183-9. [CrossRef]
5. Pandian TJ, Sheela SG. Hormonal induction of sex reversal in fish. Aquaculture. 1995;138(1-4):1-22. [CrossRef]
6. Mousavi-Sabet H, Langroudi HF, RohaniRad M. Sex reversal, mortality rate and growth of guppy (Poecilia reticulata) affected by 17-alpha methyltestosterone. Poeciliid Res. 2012;2(1):1-8.
7. Piferrer F, Lim LC. Application of sex reversal technology in ornamental fish culture. Aquarium Sci Conserv. 1997;1:113-8. [CrossRef]
8. Baroiller JF, Cotta DH. The reversible sex of gonochoristic fish:Insights and consequences. Sex Dev. 2016;10(5-6):242-266. [CrossRef]
9. Tran NK, Kwan TN, Purser J, Patil JG. Masculinization of adult Gambusia holbrooki:A case of recapitulation of protogyny in a gonochorist? Biology. 2022;11(5):694. [CrossRef]
10. Homklin S, Watanodorn T, Ong SK, Limpiyakorn T. Biodegradation of 17alpha- methyltestosterone and isolation of MT-degrading bacterium from sediment of Nile tilapia masculinization pond. Water Sci Technol. 2009;59(2):261-5. [CrossRef]
11. Yamazaki F. Sex control and manipulation in fish. Aquaculture. 1983;33(1-4):329-54. [CrossRef]
12. Mlalila N, Mahika C, Kalombo L, Swai H, Hilonga A. Human food safety and environmental hazards associated with the use of methyltestosterone and other steroids in production of all-male tilapia. Environ Sci Pollut Res. 2015;22:4922-31. [CrossRef]
13. Permen KP. Peraturan Menteri Kelautan Dan Perikanan Republik Indonesia Nomor 1/Permen-Kp/2019 Tentang Obat Ikan. Indonesian, Jakarta:Menteri Kelautan Perikanan Republik Indonesia;2019. Available from: https://peraturan.bpk.go.id/details/159085/permen-kkp-no-1permen-kp2019-tahun-2019 [Last accessed on 2024 Apr 01].
14. Nurkhasanah A. Masculinization of Rainbowfish (Iriatherina werneri) Through Embryo Immersion in Purwoceng Plant Extract (Pimpinella alpina). Indonesian:Thesis Bogor Agricultural University;2015. Available from: https://repository.ipb.ac.id/handle/123456789/77820 [Last accessed on 2024 Apr 01].
15. Arfah H, Tri SD, Asep B. Masculinization of betta fish (Betta splendens) through embryo immersion in purwoceng extract (Pimpinella alpina). J Akuatr Indones. 2013;12(2):144-9. [CrossRef]
16. Rohmy S, Camar O, Junior MZ. Masculinization of ornamental fish Synodontis eupterus through embryo immersion using purwoceng extract (Pimpinella alpina). Indonesian:Thesis Bogor Agricultural University;2018. Available from: https://repository.ipb.ac.id/handle/123456789/95199 [Last accessed on 2024 Apr 10].
17. Matondang AH, Basuki F, Nugroho RA. The Effect of Immersion Duration of Female Broodstock in Purwoceng Extract (Pimpinella alpina) on the Masculinization of Guppy Fish (Poecilia reticulata). Uttar Pradesh:JAMTech;2018. 10-17. Available from: https://ejournal3.undip.ac.id/index.php/jamt/article/view/20363 [Last accessed on 2024 Apr 15].
18. Putra S. Masculinization of Nile Tilapia (Oreochromis niloticus) Through Immersion in Purwoceng Extract (Pimpinella alpina). Indonesian:Thesis Bogor Agricultural University;2011. Available from: https://repository.ipb.ac.id/handle/123456789/52627 [Last accessed on 2024 Apr 17].
19. Pradana FM, Basuki F, Nugroho RA. The Effect of Purwoceng Extract (Pimpinella alpina) on the Masculinization of Nile Tilapia (Oreochromis niloticus) with Different Larval Immersion Durations. Indonesian:JAMTech;2017. 85-94. Available from: https://ejournal3.undip.ac.id/index.php/jamt/article/view/20481/19292 [Last accessed on 2024 May 21].
20. Purwestri YA, Kartikasari NA, Putri SG, Wilson W, Sembiring L. Metabolic Profiling of Endophytic Bacteria from Purwoceng (Pimpinella pruatjan molkend) Root and Antibacterial Activity Against Staphylococcus aureus and Pseudomonas aeruginosa. AIP Conference Proceedings; 2016.1744. [CrossRef]
21. Sumarni E, Soesanto L, Farid N, Baroroh HN. Growth and development of purwoceng plants in hydroponic cultivation using the nutrient film technique (NFT). J. Litbang. 2017;15(2):145-51. [CrossRef]
22. Darwati I, Roostika I. Research status of purwoceng (Pimpinella alpina Molk) Indonesia. Bull Plas Nutf. 2006;12(1):9-15. [CrossRef]
23. Wu J, Cao Z, Hassan SS, Zhang H, Ishaq M, Yu X, et al. Emerging biopharmaceuticals form Pimpinella genus. Molecules. 2023;28(4):1571. [CrossRef]
24. Saini RK, Song MH, Yu JW, Shang X, Keum YS. Phytosterol profiling of apiaceae family seeds spices using GC-MS. Foods. 2021;10(10):2378. CrossRef]
25. Maclatchy DL, Vanderkraak GJ. The phytoestrogen beta-sitosterol alters the reproductive endocrine status of goldfish. Toxicol Appl Pharmacol. 1995;134(2):305-12. [CrossRef]
26. MacLatchy D, Peters L, Nickle J, Vanderkraak G. Exposure to ß-sitosterol alters the endocrine status of goldfish differently than 17b-estradiol. Environ Toxicol Chem. 1997;16(9):1895-904. [CrossRef]
27. Qasimi MI, Mohibbi H, Nagaoka K, Watanabe G. Effects of phytosterols as food additives on adrenal and reproductive endocrine function during sexual maturation in male Japanese quail (Coturnix coturnix japonica). J Poul Sci. 2018;55(2):155-161. [CrossRef]
28. Gilman CI, Leusch FD, Breckenridge WC, Maclatchy DL. Effects of a phytosterol mixture on male fish plasma lipoprotein fractions and testis P450scc activity. Gen Comp Endocrinol. 2023;130(2):172-84. [CrossRef]
29. Piferrer F, Donaldson EM. Gonadal differentiation in Coho Salmon, Oncorhynchus kisutch, after a single treatment with androgen or estrogen at different stages during ontogenesis. Aquaculture. 1989;77(1989):251-62 [CrossRef]
30. Weber GM, Leeds TD. Effects of duration and timing of immersion in 17a-methyltestosterone on sex reversal of female rainbow trout. Aquacu Rep. 2022;23:101014. [CrossRef]
31. Devlin RH, Nagahama Y. Sex determination and sex differentiation in fish:An overview of genetic, physiological, and environmental influences. Aquaculture. 2002;208(3-4):191-364. [CrossRef]
32. Rajendiran P, Jaafar F, Kar S, Sudhakumari C, Senthilkumaran B, Parhar IS. Sex determination and differentiation in teleost:Roles of genetics, environment, and brain. Biology (Basel). 2021;10(10):973. [CrossRef]
33. Kamaszewski M, Skrobisz M, Wójcik M, Kawalski K, Szczepanski A, Bujarski P, et al. The role of transcription factors in gonad development and sex differentiation of a teleost model fish-guppy (Poecilia reticulata). Animals (Basel). 2020;10(12):2401. [CrossRef]
34. Piferrer F. Endocrine sex control strategies for the feminization of teleost fish. Aquaculture. 2001;197(1-4):229-81. [CrossRef]
35. George T, Pandian TJ. Production of ZZ females in the female-heterogametic black molly, Poecilia sphenops, by endocrine sex reversal and progeny testing. Aquaculture. 1995;136:81-90. [CrossRef]
36. Kavumpurath S, Pandian TJ. Masculinization of Poecilia reticulata by dietary administration of synthetic or natural androgen to gravid females. Aquaculture. 1993b;116:83-9. [CrossRef]
37. SvobodováZ, Lloyd R, MáchováJ, VykusováB. Water Quality and Fish Health. Rome:FAO;1993. Available from: https://www.nativefishlab.net/library/textpdf/15062.pdf [Last accessed on 2024 May 17].
38. Yúfera M, Darias MJ. The onset of exogenous feeding in marine fish larvae. Aquaculture. 2007;268(1-4):53-63. [CrossRef]
39. Brownell CL. Water quality requirements for first-feeding in marine fish larvae. 1. Ammonia, nitrite, and nitrate. J Exp Mar Biol Ecol. 1980;44(2):269-83. [CrossRef]
40. Fernando AA, Phang VP. Culture of the guppy, Poecilia reticulata, in Singapore. Aquaculture. 1985;51(1):49-63. [CrossRef]
41. Malik T, Syaifudin M, Amin M. Masculinization of guppy fish (Poecilia reticulata) through the use of coconut water (Cocos nucifera) with different concentrations. J Akuaktr Raw Indones. 2019;7(1):13-24. [CrossRef]
42. Boyd CB, Tucker CS. Pond Aquaculture Water Quality Management. Boston:Kluwer Academic Publishers;1998. [Last accessed on 2024 Apr 17]. [CrossRef]