
1. INTRODUCTION
Global food security remains a paramount concern with the pressure of a growing population and the emphasis on sustainable agriculture practices. One of the major hurdles of contemporary agriculture is the management of plant diseases, which profoundly impact crop productivity [1]. Late blight caused by Phytophthora infestans epitomizes such challenges by affecting globally popular plants like tomato (Solanum lycopersicum), leading to substantial loss in the agriculture sector [2]. The pathogen infects various parts of plants, including roots, tubers, and shoots, leading to substantial yield loss [3]. The traditional means of controlling this disease is by using fungicides, which led to multiple issues in the long term, such as environmental pollution, negative effects on non-target organisms, and the emergence of fungicide-resistant microorganisms [4].
Recent research has increasingly focused on the potential of biocontrol agents, particularly iturin A, a cyclic lipopeptide (CLP) produced by various Bacillus species such as Bacillus velezensis, Bacillus amyloliquefaciens and Bacillus atrophaeus [5–7]. Iturin A has been identified as the main active antimicrobial and antifungal compound in Bacillus strains, showcasing its potential as a biocontrol agent for combating a broad spectrum of bacterial and fungal diseases [7,8]. Studies have shown that iturin A effectively inhibits the growth of several fungal species such as Fusarium oxysporum, and Colletotrichum acutatum, by affecting their mycelial morphology, conidia germination, and cell membrane integrity [9,10,8,11]. The compound's mechanism of action appears to involve disrupting fungal cell membranes, increasing reactive oxygen species levels, and affecting gene expression related to fungal growth and toxin production, ultimately leading to fungal cell death [9,12].
Moreover, iturin A has proven effective in suppressing plant diseases caused by various pathogens, including Phytophthora species, which are responsible for root rot and blight, as well as controlling diseases like Verticillium wilt in cotton and Fusarium wilt in lettuce, and rice blast in rice [8,11–13]. These findings underscore the compound's potential as a sustainable biocontrol strategy for managing plant diseases. While the precise mechanism of iturin A's antimicrobial activity is not fully understood, it is believed to involve the disruption of microbial cell membrane integrity, resulting in cell lysis and death [11]. Additionally, iturin A possesses surfactant properties, which enhance the spreadability of liquids, making it a valuable component in various industrial applications, including the formulation of agricultural products and cleaning agents [13,14]. Its properties also suggest a promising role as a biopesticide for controlling fungal diseases [8,13,15].
Several chemical fungicides such as Polyversum and Biochikol 020 PC are known to significantly reduce P. infestans infestation in potato plants [16]. Similarly, Kurzawi?ska and Mazur reported decreased canopy infection and tuber infestation in potatoes treated with these biotechnical preparations [17]. Similarly, Tomar et al. [17] identified Pseudomonas aeruginosa-1 as a promising biocontrol agent against P. infestans, which inhibits disease in whole potato plant tests [18]. Ndala et al. [19] evaluated the efficacy of plant extracts of Plectranthus barbatus, Lantana camara, and Sphaeranthus suaveolens showing comparable results to a commercial fungicide in managing late blight disease in tomatoes [19]. Despite these promising findings, there remains a gap in developing a comprehensive bioformulation utilizing these agents. Therefore, the present study deals with the intent to utilize the bioactive potential of iturin A derived from B. velezensis for the development of a bioformulation specifically targeting the management of P. infestans-induced late blight in tomato (S. lycopersicum) cultivation, offering a sustainable alternative to traditional chemical fungicides.
2. MATERIAL AND METHODS
2.1. Inhibition of pathogen P. infestans by B. velezensis
2.1.1. Collection of samples
Kolar tomato field, at latitude 13.1362° N, longitude 78.1291°, situated in Karnataka, India was explored for late blight symptoms (green to light brown or tan patches) on tomato leaves in the month of June 2022. infected leaf samples were collected and stored in sterile plastic bags for transportation to the laboratory. Bacteria were isolated and purified from the soil of the same field where healthy plants were spread for at least 20m×20m block. The soil samples were collected and placed in sterile plastic bags, ensuring their cleanliness, and promptly storing them in a cold environment using ice packs.
2.1.2. Isolation of P. infestans
Infected leaves were dried and cut into small pieces, which were cultivated on potato dextrose agar (PDA) plates fortified with tetracycline (10 mg/l) at 20ºC under dark conditions as described by Rakesh and Shivkumar [20]. The oomycete P. infestans was isolated for further study.
2.1.3. Isolation of B. velezensis
Three different culture media compositions were standardized and used for growing B. velezensis. All chemicals were purchased from Himedia Laboratories Pvt Ltd, India. In the M3A medium, the main components include malt extract at 6 g/l, glucose at a high concentration of 40 g/l, and yeast extract at 5 g/l, supplemented with 5 g/l each of K2HPO4 and MgSO4, along with a trace of ZnSO4 (1 g/l) in 1,000 ml of water. The F medium relied on 25 g/l of ragi flour as a complex carbon source, alongside 10 g/l of CaCO3 and KH2PO4, and only 2 g/l of glucose, in 1,000 ml of water. Another GMK medium was composed of 10 g/l glucose and 5 g/l L-monosodium glutamate as primary nutrients, supported by several salts: 0.5 g/l MgSO4, 0.78 g/l KCl, 1 g/l KH2PO4, 0.05 g/l FeSO4, 5 g/l MnSO4, and 0.16 g/l CuSO4 in distilled water, with the pH adjusted to 7.2. Each of these media provided a unique composition to support the growth and metabolic activity of B. velezensis.
Initially, 2 g of soil was mixed with 2 ml of sterile phosphate buffered saline and allowed to incubate for 2 h. Following this, the tubes underwent centrifugation at 5,000 rpm for 5 min, and subsequently, 100 µl of the supernatant were inoculated into each of the culture media in triplicate. Incubation took place at 30°C for 2 days [21]. Morphologically different colonies were isolated and maintained in the same media. The species were identified by molecular characterization, and sequences were submitted to NCBI.
2.1.4. Molecular characterization of B. velezensis
Genomic DNA was isolated from bacterial colonies using a bacterial DNA isolation kit (Qiagen, New Delhi). Genomic DNA was amplified using 27F Forward (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R Reverse (5′-CGGCTACCTTGTTACGACTT-3′) primers of 16S rRNA gene [21]. The PCR was performed in a 40 µl mixture containing Master mix, PCR water, primers, and template DNA. Cycling conditions included enzyme activation at 96°C for 10 minutes, followed by 40 cycles of denaturation at 95°C, annealing at 52°C, and elongation at 72°C. The final extension was at 72°C for 10 minutes. Amplicon size were confirmed on agarose gel and sequenced using Thermofisher Applied Biosystems Sequencer (India).
2.1.5. In vitro Inhibition of P. infestans using B. velezensis
Bacillus velezensis living cells (LCs) were incubated on the same liquid medium for 24 hours at 37ºC with agitation (200 rpm), and the final concentration was adjusted to 1×108 CFU/ml with distilled water. Cellulose acetate membranes (0.22 µm) were used to filter cell suspension (CS) and to obtain a cell-free supernatant.
Phytophthora infestans mycelium was agitated at 10ºC for 3 hours to release sporangium followed by a zoospore formation. Zoospore suspension having 1 × 107 colony-forming unit in 1 ml culture was used to find inhibitory effects of B. velezensis. Phytophthora infestans mycelium growth was evaluated using B. velezensis living cells and CS through the dual-culture plate method as described by Wang et al. [22]. After coincubation at 25ºC for 5 days, the inhibitory zones were identified and inhibition rates were observed under a compound microscope (Magnus Compound Microscope). The inhibition rate (%) was determined using the following formula: (C - T)/C × 100, where T is the radius of the treatment group and C represents the radius of the colony of the control group.
2.2. Extraction of iturin A
To extract iturin A from a culture of B. velezensis cells (1 × 108 CFU/ml) CS was inoculated into 500 ml of medium within a 1l flask. The flask was then incubated at 37°C for a period of 72–96 hours with shaking at 200 rpm until the culture reached the stationary phase of growth. It was followed by the centrifugation of bacterial culture at 8,000 rpm for 10 minutes to pellet the cells, after which the supernatant was carefully decanted. The decanted supernatant was then combined with an equal volume of ethyl acetate in a separatory funnel. After vigorous shaking for 1 hour, the mixture was allowed to settle, facilitating the separation of layers. Subsequently, the upper organic layer, containing the extracted CLPs was collected. This organic extract was then dried using a rotary evaporator operating at 40°C under reduced pressure, resulting in the isolation of crude CLPs [22].
2.3. High-performance liquid chromatography (HPLC) Purification of iturin A
HPLC system (Shimadzu Analytical Pvt. Ltd., India) with a reverse-phase C18 column was used for the study. Standard iturin A (Sigma-Aldrich) and extract solution (10 mg/l) were run separately on an HPLC system. The solutions were then introduced into the system, where elution was carried out using a gradient of water and methanol. The rate of injection was 1 ml/minute and the eluate was monitored at 280 nm to detect the peaks corresponding to iturin A. Once identified, these peaks were collected for further processing.
2.4. Electrospray ionization mass spectrometry
Purified lipopeptides, at a concentration of 1 mg/ml (obtained from HPLC peaks), were analyzed using ESI-MS in collision-induced dissociation mode to determine the amino acid sequences of CLP iturin A. An optimal collision energy ranging from 35 to 50 eV was chosen based on the precursor targeted ions. The molecular mass was determined, and molecular structure was confirmed.
2.5. Nuclear magnetic resonance spectroscopy
The purified iturin A was dissolved in deuterated chloroform (CDCl3). Subsequently, 1H and 13C NMR spectroscopy were performed to determine the structure in detail, with a specific focus on the peptide and lipid components.
2.6. Preparation of bioformulation using iturin A
Two different compositions of co-formulant (Table 1) were prepared and iturin A was dissolved in it to make the final concentration of iturin A 5, 10, 15, and 20 mg/ml. Twelve bioformulations (Table 2) were explored for their potential against pathogen P. infestans and compared with metalaxyl fungicide which is commonly used to control Oomycete fungi diseases in plants.
 | Table 1. Ingredients of co-formulants. [Click here to view] |
 | Table 2. Details of bioformulations and coformulations used to inhibit P. infestans mycelium under laboratory conditions. [Click here to view] |
2.7. Effect of bioformulation on mycelium growth of P. infestans
Bioformulation (RPB1- RPB12) at different doses (5%, 10%, 15%, 20%, 25%, and 30%) were added PDA media fortified with tetracycline (10 mg/l). To assess antagonism against mycelial growth, dual culture assays were conducted. In this setup, 10 days old P. infestans mycelial of size 7-mm-diameter was placed at the center of a 90 mm PDA medium and medium containing different doses of bioformulation. Media without Bioformulation was taken as negative control and media with metalaxyl (10%) was taken as positive control. Each composition was taken in 10 petri plates in triplicate. After 7 days of dark growth at 20°C, P. infestans colony diameter was measured using the cross method with a ruler, and the inhibition rate was calculated.
2.8. In vivo antagonism tests of pathogenicity in tomato plant
Solanum lycopersicum hybrid Variety Saaho (TO-3251) was purchased from Syngenta, India. Plug trays filled with sterile cocopeat were used to sow the seeds. Germination started after 6 days followed by seedling formation. After 24 days, seedlings were transferred to pot containing sterile red soil containing 50 g of farmyard manure per kg of soil. The experiment was performed after 25 days when tomato plants achieved 25–40 cm height and 4–5 leaves. 5 ml of P. infestans culture broth at a concentration of 108 CFU/ml was mixed with 100 ml water and applied to both the adaxial and abaxial leaf surfaces using a hand sprayer (Oriley Handheld, India). 100 µl of this suspension was applied per 600 cm2 leaf surface. A day after fungal spray, bioformulation RPB 3, RPB4, and RPB 12 were sprayed on tomato leaves. 100 µl of the bioformulation composition was applied per 600 cm2 leaf surface. Disease assessment was performed after 2, 4, and 6 days.
2.9. Assessment of disease severity
According to Romanazzi et al. [23], the percentage of leaf surface covered by late blight symptoms was used to assign the disease severity to 11 levels, ranging from 0 (healthy leaf) to 10 (over 90% of leaf surface exhibiting symptoms) [23]. Percent disease incidence on leaves and disease severity index (%) were calculated for control, iturin A formulation, and standard metalaxyl (10%). The efficacy of formulation in reducing the disease incidence (%) was calculated by the formula: Disease incidence of control- Disease incidence of formulation/Disease incidence of control X 100 [24].
2.10. Statistical analysis
All data obtained was subjected to ANOVA analysis using SPSS 26.0 Software. p-value was calculated for each dataset. p = <0.05% was considered a significant difference.
3. RESULTS
3.1. In vitro inhibition of P. infestans by B. velezensis
3.1.1. Sample collection and isolation of P. infestans
Infected leaf samples showing amorphous, dark brown, or tan spots were collected from the field. P. infestans was isolated from the PDA culture medium and identified through molecular characterization of ITS sequences. Sequences were deposited at NCBI with accession numbers OR677916. Spraying of 1 ml of P. infestans culture (10 ml inoculum/l) on 1,000cm3 resulted in black/brown lesions followed by up to 80% wilting of leaves and fruits within 7 days of spray. It confirmed the isolated fungi was P. infestans.
3.1.2. Isolation and characterization of B. velezensis
Three bacterial cultures were isolated from media M3A. DNA isolation and molecular characterization using 16S ribosomal RNA gene primers identified the sequences. BLAST similarity analysis of these sequences confirmed this strain as B. velezensis. The sequence was submitted to NCBI and GenBank accession number PP434602 was allotted.
3.1.3. In vitro Inhibition of P. infestans
Bacillus velezensis living cells and CS showed 85.4% ± 2.31% and 95.6% ± 4.34% inhibition rate against P. infestans.
3.2. Extraction and identification of iturin A
The extraction process resulted in 2.5 g/l dried powder of CLPs. HPLC analysis of this powder was compared with standard iturin A to know the concentration of iturin in the sample. Standard iturin A peaked at 5.5 minutes at 200 AU intensity. Purification of crude lipopeptide extract (CLE) from B. velezensis showed the chromatogram for the CLE peaked at 5.2 minutes at 263.20 AU intensity upon analytical ultracentrifugation. It was used against a calibration curve to determine the concentration of iturin (Fig. 1 a–c). The concentration of iturin A in the sample was noted as 210 mg/g of dried CLP.
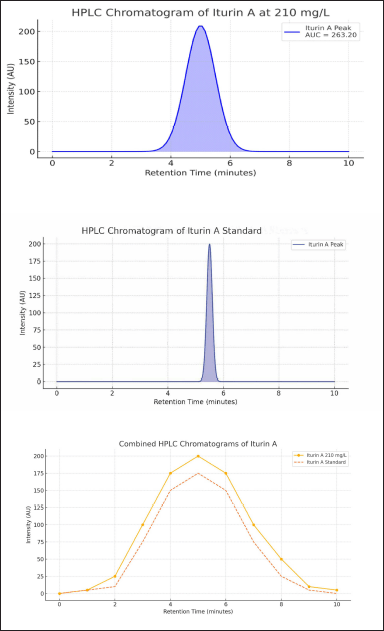 | Figure 1. HPLC chromatogram of iturin A. 1a. iturin A chromatogram from B. velezensis Sample 1b: iturin A chromatogram of standard 1c. Combined HPLC chromatogram of iturin A [Click here to view] |
Mass spectrometry showed the most abundant molecular weight at 1043 Da along with several smaller peaks at lower and higher m/z values representing fragment ions (Fig. 2). This ion peak at m/z 1,043 was considered iturin A. It confirmed the abundance of iturin A in B. velezensis. The NMR data show chemical shifts for hydrogen atoms from 2.70–6.90 ppm. Figure 3 showed signals of a methyl group at δ 2.80 ppm and protons of tyrosine at δ 6.81 ppm at C14 fatty acid side chain in iturin A.
 | Figure 2. ESI-Mass Spectrum of iturin A. [Click here to view] |
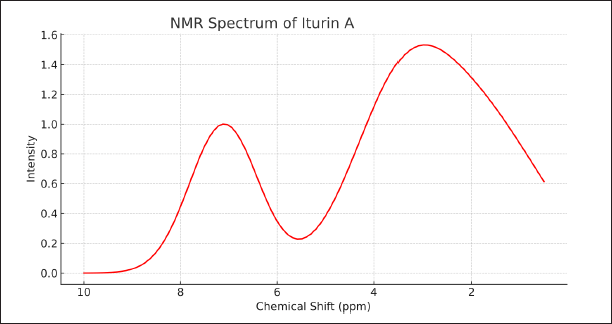 | Figure 3. NMR spectrum of iturin A. [Click here to view] |
3.3 Antagonism tests of iturin A against P. infestans
The effectiveness of different bioformulations and coformulations containing varying concentrations of iturin A against P. infestans mycelium was evaluated (Table 2). Full growth of P. infestans mycelium was observed after 4 days of inoculation (Fig. 4a). RPB1-RPB12 based on coformulations 1 and 2 had iturin A concentrations ranging from 5 mg/ml to 30 mg/ml. RPB1- RBP6 based on co-formulation 1 corresponded the percent inhibitions ranging from 40.5% to 100%. Notably, higher concentrations of iturin A (RPB4, RPB5, and RPB6) with concentrations of 20 mg/ml and above achieved complete inhibition (Fig. 4b). However, bioformulations with coformulation 2 were noted less effective in suppressing P. infestans mycelium growth (Fig. 4c). Petri plates containing metalaxyl (10%) showed 95% inhibition of P. infestans mycelium (Fig. 4d).
 | Figure 4. Antagonism tests of iturin A against P. infestans. 4a: P. infestans; 4b: P. infestans and RPB4; 4c. P. infestans and RPB8; 4d. P. infestans with metalaxyl (10%) [Click here to view] |
3.4. Evaluation of bioformulation on tomato under greenhouse condition
The bioformulations RPB3, RPB4, and RPB12 showed promising efficacy in reducing disease incidence and severity index in leaves. Specifically, RPB4 exhibits the highest efficacy among the bioformulations in reducing disease incidence and severity index, with values of 95.50% and 96.66%, respectively. Its impact was comparable with metalaxyl (10%), a chemical fungicide (Fig. 5a–e). RPB3 and RPB12 also demonstrate considerable efficacy, with disease incidence reduction rates of 81.50% and 77.27%, respectively, and disease severity index reduction rates of 84.25% and 80.5%, respectively (Table 3; Fig. 5b, d).
 | Table 3. Efficacy of bioformulations to control late blight caused by P. infestans. [Click here to view] |
 | Figure 5. Study of bioformulations efficiency in controlling late blight caused by P. infestans. 5a: Tomato plant infected with P. infestans; 5b: Effect of the spray of RPB3 on infected plant; 5c. Effect of the spray of RPB4 on infected plant; 5d. Effect of the spray of RPB12 on infected plant; 5e: Effect of the spray of metalaxyl on the infected plant [Click here to view] |
4. DISCUSSION
Several species of Bacillus have emerged as promising biological control agents (BCAs) in recent years [25]. The present investigation confirms that the bacterium B. velezensis, isolated from soil in tomato fields, has emerged as a highly effective biological control agent (BCA) against P. infestans, the pathogen responsible for late blight disease in tomatoes. This study highlights the significant potential of B. velezensis strains in biocontrol applications, primarily through the secretion of enzymes such as proteases and cellulases, and the production of stable antagonistic compounds that lead to morphological deformation of P. infestans hyphae, achieving up to 85%–95% inhibition of mycelial growth [26]. Strain SDTB038 of B. velezensis notably reduced late blight incidence by 40.79%, primarily through the synthesis of lipopeptides that inhibit P. infestans infection [27]. Additionally, various strains of B. velezensis, including ND, UTB96, T701, ES2-4, and BR-01, have been reported to contain gene clusters encoding secondary metabolites such as iturin, difficidin, and bacilysin. These metabolites are crucial in downregulating genes associated with the pathogenicity and virulence of P. infestans, thereby enhancing biocontrol efficacy [28–32].
The present study also exhibited remarkable efficacy in inhibiting the P. infestans mycelium using iturin A extracted from B. velezensis. The extraction of iturin A from B. velezensis and using that to formulate a bioformulation to control P. infestans mark it as the first documented instance of such pronounced effectiveness. This underscores the significance of Bacillus species as prevalent and extensively researched biological control agents (BCAs), as highlighted in prior studies.
A key focus of this investigation is the use of iturin A, a secondary metabolite produced by B. velezensis. Recent findings indicate that strains capable of synthesizing iturin A, such as those mentioned above, can effectively inhibit the growth of P. infestans mycelium. The present study also exhibited remarkable efficacy in inhibiting the P. infestans mycelium using iturin A extracted from B. velezensis. The extraction of iturin A from B. velezensis and using that to formulate a bioformulation to control P. infestans mark it as the first documented instance of such pronounced effectiveness. This underscores the significance of Bacillus species as prevalent and extensively researched biological control agents (BCAs), as highlighted in prior studies [5,30,32]. Beyond its role in plant pathogen control, iturin A has diverse industry applications. It has been employed in the synthesis of silver nanoparticles and as a biosurfactant for enhanced oil recovery in the petroleum sector [33]. Furthermore, iturin-based bioformulations have shown potential against various pathogens, including bacteria and insects, and have applications in food packaging and water purification [34–35]. However, its use against plant pathogenic fungi, specifically in controlling late blight, remains underexplored. The development of the bioformulation RPB4, which leverages the potential of iturin A to combat P. infestans, represents a promising and innovative solution in sustainable agriculture. This study underscores the pivotal role of Bacillus species as BCAs, reinforcing their importance in ongoing and future research efforts aimed at managing plant diseases effectively and sustainably.
5. CONCLUSION
The present study has suggested that bioformulations, particularly RPB4, having 20 mg/l of iturin A can be an effective alternative to chemical fungicides like metalaxyl for managing diseases in leaves, offering potential benefits such as reduced environmental impact and improved sustainability in agriculture. Further field application of Bioformulation RPB4 could be explored to optimize the application methods and dosages. Additionally, optimizing the formulation parameters, such as concentration, application method, and timing, is crucial for maximizing efficacy while minimizing costs and environmental impact. This optimization process can be facilitated through field trials conducted under diverse environmental conditions to ensure the reliability and consistency of the results. Furthermore, assessing the long-term effects of these bioformulations on soil health, microbial communities, and non-target organisms is essential for evaluating their overall sustainability and safety. Comprehensive risk assessments will enable informed decision-making regarding their integration into agricultural practices on a larger scale
6. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
7. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
11. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
12. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Talukder B, Blay-Palmer A, Vanloon GW, Hipel KW. Towards complexity of agricultural sustainability assessment: main issues and concerns. Environ Sustain Indic 2020;6:100038.
2. Mazumdar P, Singh P, Kethiravan D, Ramathani I, Ramakrishnan N. Late blight in tomato: insights into the pathogenesis of the aggressive pathogen Phytophthora infestans and future research priorities. Planta 2021;253(6):119.
3. Ivanov AA, Ukladov EO, Golubeva TS. Phytophthora infestans: an overview of methods and attempts to combat late blight. J Fungi 2021;7(12):1071.
4. Pavani Y, Janaki P, Jagadeeswaran R, Sankari A, Ramalakshmi A, Arthanari PM. Response of tomato to fertilizer nutrients integration and herbicides spray: evaluating growth, yield, fruit quality and herbicides residue. Plant Sci Today 2024;11(1):93–101.
5. Zhongming T, Xiaojie C, Huili Z. Production of iturin A by Bacillus velezensis ND and its biological control characteristics. J Basic Microbiol 2023;63(2):179–89.
6. Junhua W, Jiying Q, Xiaoyu Y, Jinyu Y, Shuangzhi Z, Qingxin Z, et al. Identification of Lipopeptide iturin A produced by Bacillus amyloliquefaciens NCPSJ7 and its antifungal activities against Fusarium oxysporum f. sp. niveum. Foods 2022;11(19):2996.
7. Moghtadi SP, Madani M, Shakib P. Detection of iturin A from Bacillus atrophaeus by HPLC and mass spectrometry analysis and evaluation of its antifungal activity. Curr Drug Ther 2022;19(6):742–7.
8. Singh P, Xie J, Qi Y, Qin Q, Jin C, Wang B, et al. A thermotolerant marine Bacillus amyloliquefaciens S185 producing iturin A5 for antifungal activity against Fusarium oxysporum f. sp. cubense. Mar Drugs 2021;19(9):516.
9. Hua M, Deng Q, Qiu M, Cai D, Sun L, Fang Z, et al. iturin A strongly inhibits the growth and T-2 toxin synthesis of Fusarium oxysporum: a morphological, cellular, and transcriptomics study. Foods 2023;12(6):1278.
10. Yang DD. Target mechanism of iturinic lipopeptide on differential expression patterns of defense-related genes against Colletotrichum acutatum in pepper. Plants 2022;11(9):1267.
11. Fujita S, Yokota K. Disease suppression by the cyclic lipopeptides iturin A and surfactin from Bacillus spp. against Fusarium wilt of lettuce. J Gen Plant Pathol 2019;85:44–8.
12. Ma Z, Zhang S, Sun K, Hu J. Identification and characterization of a cyclic lipopeptide iturin A from a marine-derived Bacillus velezensis 11-5 as a fungicidal agent to Magnaporthe oryzae in rice. J Plant Dis Prot 2020;127:15–24.
13. Dhuldhaj UP, Bora CR. Microbial surfactants an overview. Microb Surfactants 2021;29:1–26.
14. Gao L, She M, Shi J, Cai D, Wang D, Xiong M, et al. Enhanced production of iturin A by strengthening fatty acid synthesis modules in Bacillus amyloliquefaciens. Front Bioeng Biotechnol 2022;10:974460.
15. Jiang M, Pang X, Liu H, Lin F, Lu F, Bie X, et al. Iturin A induces resistance and improves the quality and safety of harvested cherry tomato. Molecules 2021;26(22):6905.
16. Kurzawi?ska H, Mazur S. The application of some biotechnical preparations in potato protection against Phytophthora infestans. Phytopathologia 2012;63:31–7.
17. Kurzawi?ska H, Stanis?aw M. The evaluation of Pythium oligandrum and chitosan in control of Phytophthora infestans (Mont.) de Bary on potato plants. Folia Hortic 2009;21(2):3–23.
18. Tomar S, Singh BP, Lal M, Khan MA, Hussain T, Sharma S, et al. Screening of novel microorganisms for biosurfactant and biocontrol activity against Phytophthora infestans. J Environ Biol 2014;35(5):893.
19. Ndala RI, Mbega ER, Ndakidemi PA. Different plant extracts against Phytophthora infestans (Mont.) de Bary in tomato in vitro. Am J Plant Sci 2019;10(4):698–708.
20. Rakesh P, Shivkumar N. Identification, characterization, and pathogenicity assessment of oomycetes fungi responsible for damping off and late blight in Indian tomato crops. Biochem Cell Arch 2024;24(1):1143–8.
21. Wekesa TB, Wekesa VW, Onguso JM, Wafula EN, Kavesu N. Isolation and characterization of Bacillus velezensis from Lake Bogoria as a potential biocontrol of Fusarium solani in Phaseolus vulgaris L. Bacteria 2022;1(4):279–93.
22. Wang Y, Zhang C, Liang J, Wu L, Gao W, Jiang J. Iturin A extracted from Bacillus subtilis WL-2 affects Phytophthora infestans via cell structure disruption, oxidative stress, and energy supply dysfunction. Front Microbiol 2020;11:536083.
23. Romanazzi G, Mancini V, Feliziani E, Servili A, Endeshaw S, Neri D. Impact of alternative fungicides on grape downy mildew control and vine growth and development. Plant Dis 2016;100:739–48.
24. Chiang K, Liu H, Bock CH. A discussion on disease severity index values: warning on inherent errors and suggestions to maximize accuracy. Ann Appl Biol 2017;171:139–54.
25. Palmieri D, Ianiri G, Del Grosso C, Barone G, De Curtis F, Castoria R, et al. Advances and perspectives in the use of biocontrol agents against fungal plant diseases. Horticulturae 2022;8(7):577.
26. Zhang J, Huang X, Hou YC, Xia X, Zhu Z, Huang AM, et al. Isolation and screening of antagonistic endophytes against Phytophthora infestans and preliminary exploration on anti-oomycete mechanism of Bacillus velezensis 6-5. Plants 2023;12(4):909.
27. Ivana SP, Vlajkov V, Dodi? J, Loc M, Grahovac M, Grahovac J. Bacillus velezensis: Biocontrol activity of cells and extracellular compounds against Xanthomonas spp. J Process Energy Agric 2022;26(1):15–8.
28. Yan HH, Qiu Y, Yang S, Wang YQ, Wang K, Lili J, et al. Antagonistic activity of Bacillus velezensis SDTB038 against Phytophthora infestans in potato. Plant Dis 2021;105(6):1738–47.
29. Zhou Q, Tu M, Fu XQ, Chen Y, Fang Y, Yan Y, et al. Antagonistic transcriptome profile reveals potential mechanisms of action on Xanthomonas oryzae pv. oryzicola by the cell-free supernatants of Bacillus velezensis 504, a versatile plant probiotic bacterium. Front Cell Infect Microbiol 2023;13:1175446.
30. Abdelkhalek A, Behiry SI, Al-Askar AA. Bacillus velezensis PEA1 inhibits Fusarium oxysporum growth and induces systemic resistance to cucumber mosaic virus. Agronomy 2020;10(9):1312.
31. Vahidinasab M, Adiek I, Hosseini BN, Akintayo SO, Abrishamchi B, Pfannstiel J, et al. Characterization of Bacillus velezensis UTB96, demonstrating improved lipopeptide production compared to the strain B. velezensis FZB42. Microorganisms 2022;10(11):2225.
32. Zhou J, Xie Y, Liao YT, Li X, Li Y, Ma XY, et al. Characterization of a Bacillus velezensis strain isolated from Bolbostemmatis Rhizoma displaying strong antagonistic activities against a variety of rice pathogens. Front Microbiol 2022;13:983781.
33. Zhao X, Ai C, Li Z, Xu X, Zhao H, Yan L, et al. Development of a paper-based method to detect Hg 2+ in waste water using iturin from Bacillus subtilis. Appl Microbiol Biotechnol 2019;03:8609–18.
34. Wan C, Fan X, Lou Z, Wang H, Olatunde A, Rengasamy KR. iturin: cyclic lipopeptide with multifunction biological potential. Crit Rev Food Sci Nutr 2022;3(29):7976–88.
35. Yaraguppi DA, Bagewadi ZK, Mahanta N, Singh SP, Khan TY, Deshpande SH, et al. Gene expression and characterization of iturin A lipopeptide biosurfactant from Bacillus aryabhattai for enhanced oil recovery. Gels 2022;8(7):403.