
1. INTRODUCTION
In the time of 1900s, the symphonic microbiological analysis discovered by Lorenz Hiltner finds out that the most diverse or microbial density was found very close to soil [1]. The biotic and abiotic elicitors have a major role in constituting the soil microbiome [2]. The carbon energy source is the primary constituent of microbial enrichment which is given by the plant. Furthermore, plant releases10–15% of their photosynthetic and absorbs into the rhizosphere [3]. Subsequently, the composition of root microbial networks is impacted by the plant species [4-6]. Indeed, there has been coevolution between the rhizosphere and plants occupied by communities of microbial. In different plant species, the composition of rhizo-deposit varies including species-species and unique rhizo-deposit, which require further studies on various plant species to recognize and learn their impact between microbes and root interaction.
Plants have been considered as a significant medicinal herb in the arrangement of Ayurvedic medicine [2,6]. Plants possess immense pharmacological significance thus are a great subject of research interest. According to the researchers, the Ashwagandha plant is helpful to treat neurological disorders and has types of properties such as anti-cancerous, antioxidative, and immunomodulatory [7-9]. The extraction of Ashwagandha was described to diminish the two-stage skin carcinogenesis decrease with the influence of croton oil and dimethylbenzanthracene [10]. It is likewise observed to be efficient in treating arthritis, behavioral, and problems related to stress [11]. Ashwagandha is an ancient medicinal herb with multiple health benefits in the reproductive or nervous system. Ashwagandha is helpful in early recovery after an illness. In India, the consumption of Ashwagandha is similar to the consumption of ginseng in China [12] and is likewise stated to create a positive cause onto the endocrine system, inflammation of joints, neural disorder, asthma, ulcers, insomnia, epilepsy female disorders, and osteoarthritic inflammation. The decoction of plant dried roots is utilized as a tonic for the medication of hiccup, cold, cough, and is viewed as exceptionally powerful in controlling diabetes and cholesterol level, overall rejuvenating [13]. Naturopathy gains positive impact worldwide for the medication of these infections [14-16]. As per the report of the WHO, the world’s 80% of the total population relies on various kinds of drugs and APIs for their health issues [17].
These chapters give the summary of interaction between microbes and plants and increment the creation of derivatives from hairy roots. The research has been empowered by the utilization of bushy root composite plant systems, discussing the future potential, and featuring important applications. The transformation-based studies have been an advantage for research purposes including stable transgenic plant production [18, 19], secondary metabolite production, plant-pathogen interactions, and gene analysis. At present, technologies used in genetic engineering employing transformation through Agrobacterium rhizogenes are rapidly improving for cloning of plants, gene transfer which plays a key role in agriculture and plant biotechnology.
A. rhizogenes also called Rhizobium rhizogenes [20] is a strain of Gram-negative soil bacterium. It causes disease of hairy root in infected monocot and dicotyledon plants species. Moreover, its Ri (root inducing) plasmid consists of transferrable DNA encrypt root locus (rol) gene loci (rolA, rolC, and rolB) and also volunteers for the demonstration of hereditary material into the host cell. They also activate the adequate amount of growth of extremely branched hairy roots at the place of inflammation usually cotyledon or hypocotyl. It has been accounted that for the hairy root production, a high concentration of auxin signaling is required not ethylene signaling [21]. With untransformed aerial tissue, these roots systems can be maintained by culturing and hosted by plants. Over the most recent 30 years, a variety of plants for different purposes is widely used for metabolic engineering and production of recombinant protein to analyses of rhizosphere physiology and organic chemistry [22]. More recently, a biotechnology tool has been used for A. rhizogenes mediated hairy root production in various types of plant species to find a new biological vision like rhizosphere and its modeling to know the biochemical pathways involved, transformation and development of novel transgenic crops that can be utilized as a food product [23,24]. Metabolic enzyme functions can be managed by RNA interference or gene overexpression using root with a hair transformation, for example, a particular protein clarification whose main function is depicted in the species Nicotiana glauca for the biosynthesis of pyridine alkaloid [25]. For further studies of stable and reliable isotopes, hairy roots are used for a better understanding and explanation of various reactions which take place inside a biosynthetic pathway. For example, many researches on hairy roots of Ophiorrhiza pumila show that camptothecin is obtained from shikimate pathways and 2C-methyl-Derythritol 4-phosphate [26].
2. SECONDARY METABOLITES IN PLANT DEFENSE MECHANISM
Plants produced chemical constituents in the natural form for the strategy of survival called as secondary metabolites. In the plant kingdom, these secondary metabolites are regulated under different taxonomic or chemotypic group. Unlike primary metabolites such as nucleotides, amino acids, phytosterols, lipids, and organic acids, the biosynthesis of these secondary metabolites was initially related with inessentiality which was restricted to particular plant groups. These secondary metabolites as such do not participate primarily in the metabolic function in the plants such as growth, development, or reproduction. Moreover, their absence not resulting in death, but after a specific timeframe can cause danger to the survivability of the plant.
3. PHARMACOLOGICAL SIGNIFICANCE OF THE BIOACTIVE METABOLITES
Due to its tremendous pharmacological significance, the plants are being explored worldwide as a subject of considerable modern scientific research. Thus, the evaluation of their phytopharmacological activity is of great importance [27]. Various pharmacological activities are taken from the natural products, including antitumor, antiangiogenic, anti-inflammatory, cardioprotective, and immunomodulatory effects [28, 29]. Various herbal medicines are prepared for the medication of stress and anxiety, osteoarthritis, immunomodulatory, conjunctivitis, and tuberculosis in which the secondary metabolite used a constituent [30-34]. Various effects possess by these metabolites are antioxidative, immunomodulatory, anti-convulsant, adaptogenic, anticancer, and neurological effects.In the treatment of osteoarthritis, geriatric, behavioral, and anxiety Ashwagandha has been found to be very efficient [2, 35-41].
4. PLANTS SERVE AS RICH SOURCE OF BIOACTIVE METABOLITES
The direct relationship between biomedical and local use exists for the advancement of many different medicines by the utilization plants [42]. Worldwide for health and medicinal purpose on an average around 35,000–45,000 plant species are used for the treatment of various health ailments. [43]. Herbal medicine is utilized in wide forms (infusions, decoctions, ointments, powder, and syrup) worldwide [44-46] for the treatment of varied health ailments in different age groups of patients with no or fewer side effects [47,48]. Steroidal lactones, alkaloids, flavonoids, tannin, etc., are the several groups of chemical components that have been detected, extracted, and isolated from different plant sources. However, lower yield, genotypic variation, and variation in the substance of the pharmacologically active metabolite, long incubation period which is about 4–5 years between sowing, and uneconomical chemical synthesis and harvesting are the bottleneck in industrial creation of the pharmacological bioactive metabolites.
5. ENHANCEMENT OF SECONDARY METABOLITE FROM IN VITRO
This is a novel approach for producing bioactive substances with the utilization of cell or tissue culture techniques. With the utilization of this technique, we produce therapeutically important compounds from many medicinal plants. Under suitable conditions, in vitro culture of plants techniques gives a good deal with concentrating the production, regulation, and enhancement of secondary metabolites. In few cases, the cultures have been taken advantage for commercial production [49]. In vitro culturing of plant tissues gives an outstanding experimental and ethical framework to study growth, enhancement, and regulation of subsidiary products by providing them favorable conditions.
6. HAIRY ROOT COMPOSITE PLANTS AS A COMPLEMENTARY SOLUTION TO STABLE PLANT TRANSFORMATION
Colonization of rhizosphere by microorganisms results changes in plant development and growth. In different plant species, there is an absence of effective transformation method which is significant for research. However, this is present in parts by the usage of A. rhizogenes, a direct relation of A. tumefaciens and a normally developing microbe of plants [50]. Most recent 100 years have revolutionized plant molecular hereditary thus dedication gives to the genetic modification to give birth to a new industry. Both organisms are responsible of transferring T-DNA into the plant. Roots with hair are evolved out of a wide variety of different dicotyledons plant families and also few gymnosperms. Hairy roots have the capacity to promote their growth in the absence of external plant chemicals (unlike organ culture system) which are considered as an advantage of to produce hairy root cultures, at that point, the roots are highly developed and branched under sterilized in vitro conditions. For a stable genetic engineered plant generation, studies of root microbe interaction and other secondary metabolite productions hairy root cultures are used [51]. These strategies are transformative particularly for root-microorganism interaction studies because of decreased time required to create transgenic plant tissues of the recalcitrant plants and the capacity to maintained independent of tissue culture. Working Agrobacterium and different framework recommend that discharge system may give a method for Agrobacterium to accomplish the benefit along with different microbes in the acidic region of the rhizosphere [52,53].
7. MECHANISM OF AGROBACTERIUM AND PLANT CELL INTERACTION
For the gene transfer from the bacterial cell, it is critical to study the bacterium host relationship and to optimize its DNA transfer systems [54]. The bacterium attached to the plant cell surface is helpful for Agrobacterium to interact with a plant cell. A plant is infected by root inducing plasmid to transform the cell gene of a plant due to which unorganized growth of plant cells occurs. Auxin and cytokinin help stimulate plant development hormones bring about in the arrangement of profusely branched hairy roots transferring the hairy root inducing plasmid to the plant cell which is encoded by protein. Vir area of the bacterial plasmid is activated by phenolic compounds like acetosyringone that is delivered by injured cells of the host plant.
8. AGROBACTERIUM PLASMIDS CHARACTERISTICS
Agrobacterium plasmid is large in size greater than 800 kb, it contains virulence region and T-DNA which is mobile DNA component that is integrated into plant cells useful for rhizogenesis. According to opine synthesized by hairy root, the Ri plasmid is categorized into two principal classes that are agropine type (more virulent) and other types of opine [55]; Rhodes et al. (1990). Plasmid size is between 180 and 250 kbp. As indicated by the development of opine by transformed plant tissue, the plasmid is arranged into agropine, mannopine, and cucumopine. For agropine, the T region is divided into TR-DNA and TL-DNA has the size 8–30 kbp and 15–20 kbp, respectively. Others two plasmid contains a single T-DNA [55,54]. Different Agrobacterium strains plasmid show varying degrees of similarity and share a large region of similarity. Ri plasmid has origin of replication, catabolism, opine synthesis, and virulence [44]. Ends of the T-DNA have 25 bp repeats. The right end of the T-DNA is utilized for the DNA transfer but not by the left sequence [56]. The wounded area of the host plant release phenolic signal compounds to this response vir region gene product induces transmit of T-DNA [44]. Agrobacterium is treated with acetosyringone which is a plant signal compound as an outcome single-stranded linear T-DNA molecule is induced [57]. Virulence areas contain vir genes that do not enter the plant cell but cause communication of T-DNA when joining the chromosomal DNA. Border sequence also helps in the cell of plant transformation for direct T-DNA processing [58]. From the vir region, six transcripts are integrated subsequently two consecutively expressed operons (virA and virG), signals are recognized and other four operons are activated (virE, virB, virD, and virC) and this leads to T-DNA transfer [59].
9. ROLE OF T-DNA IN HAIRY ROOT INDUCTION
In the transformed plant cells, T-DNA genes are expressed [54,56]. In oncogenes, there is unlimited proliferation in the changed cell occur without the presence of externally added phytochromes [60]. Tobacco has strong impact on the differentiation with having high degree of auxins which results in the formation of roots, although increased levels of cytokinins lead to shoot induction [61-63]. The transformed cells can be grown without the trace of phytohormones due to the presence of these hormones [55,64]. Plant cells cannot catabolize opine, its gene is present in T-DNA. Opine can be used as a significant source of C and N (carbon and nitrogen) by Agrobacterium [63].
10. MECHANISM OF TRANSFER T-DNA FROM BACTERIUM TO HOST PLANT
Mechanism of transmit of transfer DNA into plant cell is a multistep process involves repair, replication, and recombination activity but its complete integration is unknown [64]. For the transmit of transfer DNA from plasmid to plant cell, certain genetic elements are essential like three chromosomal vir gene, T-DNA right border sequences, and vir genes. The three different chromosomal genes are responsible for the switch in the bacterial cell surface composition and this promotes the attachment of T-DNA to the plant cell wall [65]. Border sequences are used to delimitate the regions of Ri plasmid that is transmitted to plant cell [66]. Endonuclease enzyme is encoded by virD operon which makes the gaps in the T-DNA on both the sides of same strand. As a conclusion, free ssDNA (single-stranded DNA) is formed. By cutting RB region, the 5’-3’ single-stranded T-DNA is created. Then, this strand is transferred to plant cells [67]. When these border sequences are cut by VirD2 and VirD1, then T-DNA transfer initiates [68]. VirD2 helps to transfer to the nucleus by a nuclear localization sequence through the host importin alpha protein interaction and for efficient transfer of T-strands, this sequence is required [69]. The different effector proteins that are important for gene transfer are encoded by root inducing plasmid such as VirE, VirH, VirF, and VirD5 [Table 1]. Ri plasmid that induces adventitious hairy roots is formed closer to the site of infection. The transformed plant tissue produces unusual metabolites called opines by T-DNA gene [70,71]. TL-DNA of agropine plasmid and T-regions of cucumopine and mannopine plasmid induce the root formation [72]. TR-DNA agropine plasmid induces the roots which are phenotypically like to ordinary roots [61]. It was seen that alone TL and TR regions do not respond as strongly as when these both transformation regions are expressed together [73]. Alteration of auxin metabolism plays an important role for the outflow of root having hair phenotype in transformed cells [74]. T-DNA expression plays a crucial part in the induction of hairy root in transformed plant cell but it seems like auxin does not play a part for this expression. Transformed cells are more sensitive for auxin formation because the gene responsible for this is located on TL-DNA, yet it is restricted in certain plant species [75].
Table 1: Genes and other factors responsible in inducing transformation by Agrobacterium rhizogenes.
| Gene | Function | References |
|---|---|---|
| VirA | For the sensing of phenolic compounds and acidity | [76] |
| VirG | Induction of the vir regulon by phenolic and monosaccharide inducers. | [77] |
| Dimethoxyphenol [acetosyringone and hydroxyacetosyringone or phenolic inducer | Induce the vir genes | [78] |
| Monosaccharides | Cell walls act in concert with phenolic inducers to increase the level of induction | [76] |
| Acidic conditions (pH 5.5) | Vir gene induction | [79] |
| VirD; VirD2 | VirD2 is an endonuclease that nicks one of the two strands of the Ti plasmid at two sites which flank and delineate the T-DNA contains nuclear localization signals (NLSs) that help in direct transport of the T-DNA into the nucleus of the plant cell | [78] |
| VirB; VirB1–11 | VirB1 encodes a transglycosylase which is not included and cleaves beta-1,4 glycosidic bonds, VirB2 encodes the synthesis of pilin, the subunit of the T-pilus | [78] |
| VirC; C1 and C2 | VirC1 enhances the site-specific nicking by the VirD endonuclease and binds to the overdrive sequences, VirC1 helps to increase the no. of T-DNA copies per cell. | [80] |
| VirE; E1, E2, and E3 | VieE1 helps to protects the T-DNA against nuclease degradation and maintains the integrity of the 3’ end of the T-DNA prior to integration by the help of DNA transfer VirE1 encodes a chap erone which keeps the VirE2 protein from aggregating with itself inside Agrobacterium, virE3 encodes a host range locus | [81] |
| VirH | For a variety of compounds, these genes are usually associated with detoxification. | [81,82] |
| VirF | Mediates ubiquitination of proteins targeted for degradation by the proteasome, may be involved in proteolysis of proteins such as VirE2 | [80] |
| VirD5 | To overcome the instability of VirF, Agrobacterium transfers VirD5 into the plant where it binds to VirF and prevents its rapid turnover. | [80] |
11. HAIRY ROOTS CULTURES CHARACTERISTICS
Culture of root having hair is able to grow in hormone-free medium and have fast-growing tendency with laterally highly branched [Figure 1]. The hairy roots are generally white to brown in color, soft, adventitious, and fast growing. The root having hair has growth rate ranging between 0.1 and 2.0 g dry weight/liter per day. The capacity to form many new growing points of hairy roots making them more advantageous as compared to the conventional roots [76]. Between different species, the increment rate of hairy roots varies but difference also viewed in same species with different root clones [77].
 | Figure 1: Agrobacterium rhizogenes-induced profuse production of highly branched hairy roots on nutrient media using aseptic tissue culture technique. [Click here to view] |
12. HAIRY ROOTS CULTURE ESTABLISHMENT IN IN VITRO
Transformation is done aseptically by inoculating thick viable A. rhizogenes suspension cells with wounded plant parts. Emergence of roots takes place later the succeeding of 1–4 weeks at the particular place of transformation which is being cut off for further transfer it inside the growth medium which is hormone free but contains antibiotic to prevent contamination [78].
13. THE SENSITIVITY OF PLANT SPECIES TO AGROBACTERIUM
Different strains have different transformation ability that varies with other strains [79]. The chance for successful transformation is afflicted by plant tissue age and its differentiation status. The capacity to give rise to transformed cell also varies with the elevation of tissue differentiated after A. rhizogenes inoculation [80-83]. By assaying opine productions, the genetic modification can be confirmed. However, in hairy roots, opine production is unstable or may disappear after some time. Hence, to confirm the genetic modification, Southern blot hybridization techniques are performed to detect T-DNA [81].
14. A. RHIZOGENES: A MOLECULAR TOOL FOR RHIZOSPHERE MODELING
Hairy root production mediated by A. rhizogenes is known as a remarkable aid for the biosynthesis of subsidiary intermediates and other metabolic studies. It can also be used for various biotechnological formations of compounds which are derived from roots [82]. Roots with hairs consist of a better framework for persistent production of these intermediates in a sterile condition without using the cost-effective plant hormones in the culture medium. These organs are genetically fixed, and in host plant, they produce high content of secondary metabolites. By culture of root having hair, the alkaloid production is stable for years but its production decreases when callus is formed due to the induced roots. When roots redifferentiation is allowed, then the formation of alkaloid reappears [83,84]. Secondary metabolites are excreted into the growth medium by some hairy roots but the secondary product release varies in species. Secondary metabolite growth pattern and production also vary of root having hair culture. For commercial production, the secondary metabolites are dissociated from the growth [85] [Figure 2].
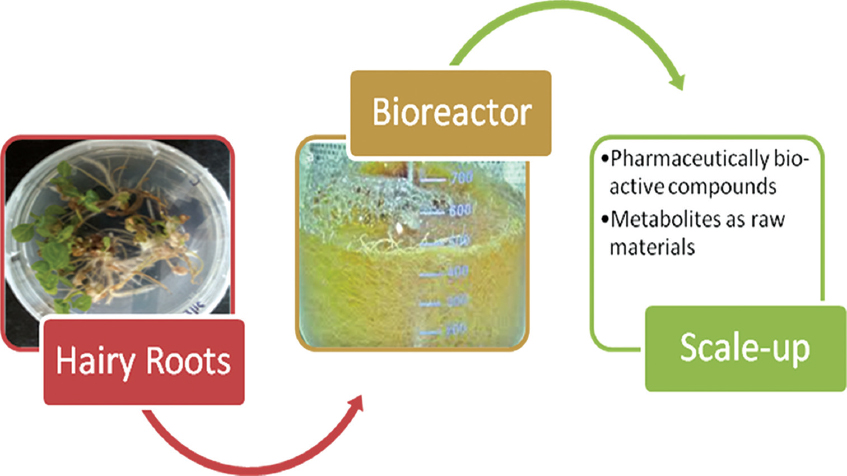 | Figure 2: Hairy root culture for mass production of bioactive compounds. [Click here to view] |
15. A. RHIZOGENES: FOR TRANSGENIC CROPS PRODUCTION
The crops which were obtained using transformation mediated by Agrobacterium are broccoli, pepper, sugarcane, carrot, barley, alfalfa, soybean, cotton, maize, wheat, rice, potato, and tomato [86,87]. Agrobacterium species has ability to transform a wide range of different neoplastic diseases, including cane gall from A. rubi, hairy root from A. rhizogenes, and crown gall from A. vitis and A. tumefaciens [88]. In the 1930s, A. rhizogenes was first identified, it belongs to Rhizobiaceae family in the alpha-2 subclass of Proteobacteria [50]. This assay is rapid and inexpensive, required simple media for rapid growth and genetic manipulation can easily be grown in Escherichia coli.
16. MOLECULAR CHARACTERIZATION OF A. RHIZOGENES-MEDIATED HAIRY ROOT INDUCTION
Effective implementation of many crop yield improvement plans through molecular rearing includes isolation of gene regulatory sequences and useful genes by exploring different plant hereditary resources. For this means, the genomic DNA arrangement should be fit for restriction cleavage, polymerase chain reactions (PCR), and construction of complete and partial gene library. Besides, the genomic DNA samples are regularly use in restriction fragment length polymorphism, random amplified polymorphic DNA, Southern blot analyses, genome fingerprinting and genome mapping, screening of transgenic lines, or in other important molecular method [2]. Literature overview throughout the previous two decades clearly uncovers that the cetyltrimethyl ammonium bromide, cationic detergent-based DNA extraction conventions were significantly more time utilized for various plant materials as compared to different protocols.
Recognition of T-DNA in the apparently changed lines was recognized and displayed by PCR [29]. Cross primers such as rolB and rolA are utilized to recognize the TL T-DNA (5-ATGGAATTAGCCGACTAAACG-3 and 5-ATGGATCCCAAATTGCTATTCC-3), which is known as the normal fragment size of around 1440 base pairs. Primers used for the TRDNA (5-AATCGTTCAGAGAGCGTCCGAAGTT-3 and 5-CGGAAATTGTGGCTCGTTGTGGAC-3) (Slightom et al., 1986) created a large fragment of 1672 base pairs. Agropine synthase gene (ags) was identified using a primer (5-AGGTCTGGCGATCGCGAGGA-3 and 5-GCGCATCCCGAGGCGATG-3) [89,90], by producing a 512 base pair fragment as an complementary indicator for the TR T-DNA. Primers used for detecting the virD1 gene (5-ATGTCGCAAGGACGTAAGCCCA-3 and 5-GGAGTCTTTCAGCATGGAGCAA-3), which is present at the external region of the T-DNA of the root inducing plasmid and are not allowed to enter inside the plant genome, were utilized to remove the chances of error of polymerase chain reaction due to A. rhizogenes contamination caused by the root lines [91].
17. ADVANTAGES OF TRANSFORMATION
Transformation through A. rhizogenes has low copy number with few rearrangements as comparison to other technologies. A. tumefaciens method is preferred to transfer bacterial virulence protein to plant which will help to target the T-DNA into the nucleus, and hence at the time of integration inside the genome, it maintains the integrity. Agrobacterium is fast growing with genetic stability and can be cultured in huge scale ferment or for the secondary metabolite production. The transformation study helps in knowing better the rhizosphere microbiome outside and inside of a susceptible plant, to know the mechanisms which maintain Agrobacterium niche in the rhizosphere and what plant signal and environmental cues are coopted to promote its function, how the signals are recognized, does any signal given from Agrobacterium to plant that can promote signaling and become beneficial to it, the molecules excreted by the bacterium affect which gene of a plant does it show any host defense, should know the outcome of Agrobacterium to protoplasm attachment to know whether this attachment affect bacterium physiology, does opine, phytohormones, and transformation products cause any changes to Agrobacterium physiology [Figure 3].
 | Figure 3: Applications of rhizosphere modeling by Agrobacterium rhizogenes-mediated transformation. [Click here to view] |
The advancement of a quick developing root culture framework would vary unique chances for construct root drugs in the research center crude turning on Pasteur developing [2,70]. In contrast with interrupted cell culture, modified roots having hair are exceptionally changed and can produce plant materials almost rich in derivatives. This raised the quantity of secondary products and, accordingly, the quick growth of transfect roots having hair is only the features of a successful production model for helpful phytochemicals.
Transgenic root cultures transfigured the job of plant tissue culture in the formation of subsidiary intermediate. They are interesting for biosynthetic and genetic conditions, proliferate, and more simply support. Utilizing this system, a broad of chemical components has been modified [55,92,93]. The promotive benefits of roots with hairs are that their cultures are frequently manifest approximately on an equivalent or larger synthesized limit for the production of secondary metabolite contrast to their mother plants [94].
In the middle of organ culture systems or different plant cell, culture of bushy root is one of the significant tools for the formation of root-derived compounds, biosynthesis of optional metabolites, and metabolic engineering studies [90]. Preceding reports recorded nearly low yields of withanolides from cultures of furry root [95].
A. rhizogenes moderate bristly root culture is a significant aid for the biosynthesis of valuable subsidiary products such as withanolides. Hairy roots are viewed a great technology for union of beneficial intermediatary components in a purified surroundings within the shortfall of cost effective development controllers inside the medium [86,74]. For some reason, A. rhizogenes moderate culture of root with hairs hairy root is consumed in various prime pharmaceutical plants for the proffering of intermediates of secondary production [95]. However, A. rhizogenes intervened hairy root inductive in Withania somnifera is restricted due to shortage of obtainable and proficient root with hairs initiation method [86]. Another methodology for a proficient root with hairs initiation is varying for extensive production of withanolides. Exertion is made to beat issues related to host/tissue to extend the amount of contaminated sites, such as utilization of highly toxic Agrobacterium strains and inclusion of few components to the crop productive medium. Laterally, sonication-assisted Agrobacterium-mediated transformation attracted high consideration in some species of plant [48,59]. It has been profitably put in for hairy root construction in opium poppy [70] and Verbascum xanthophoeniceum [52]. It gives high assurance for the increment of root with hairs creation. The lead of this technique is that the cavities achieved by sound effect create as many of microwounds on the outer layer of the explants. These microwounds allow Agrobacterium to tint prick and more absolutely all-round the explant than standard wounding, extend the probability of contamination to host cells (productivity in Agrobacterium was likewise refined in various species of plant by the use of heat care) [74].
The changed cultured of hairy roots in LB medium did not show any bacterial development demonstrating the without the presence of live A. rhizogenes, in accordance with perceptions by Hayta et al. (2011) in the culture of roots with hairs of Gentiana cruciata. In this review, a high rate of transformation (90%) was acquired in leaf explants which are infected with R1000 strain, with the development of 28.2 hairy roots (2–3 cm root length) per explants after the culturing of 12 days. The roots with hairs are delivered seen to be highly branched, fine, and soft. This broad branching, because numerous meristems are present, represented greater development rates of roots with hairs in culture as this occurrence was normal for the components of Solanaceae [78]. The novel framed roots with hairs were at first white and eventually became brown with the exception of the growth in root tips and they exhibited the typical highlights of the crown ball disorder, that is, extensive lateral branching, hormone independence, and plagiotropic growth. Plagiotropism of furry roots was trademark as the consequence of A. rhizogenes intervened change [56]. In the current review, the furry roots got risen up out of the profound injured locales of the midrib of leaf explants as seen by Tiwari et al. (2007) in Gentiana macrophylla. Nilsson and Olsson (1997) guessed that cells that contain undeniable degree of sucrose and auxin are ideal focuses for bristly root enlistment. Tiwari et al. (2007) seen that the phloem cells, situated somewhere down in plant organs, could be the target of A. rhizogenes.
Transformation mediated by Agrobacterium is one of the techniques for genetic modification in different species of plant. The biosynthetic pathway of complicated compounds of Agrobacterium is still not revealed. The pharmaceutical products that are derived by hairy roots are not been profit oriented. The main cause for this is too low content production as compare to conventional extraction. The compound produce through Agrobacterium is re-evaluated by authorities for safety, efficiency, and quality which are hard for an industry to use.
The work on the results of molecular and genetic method toward creation of genetic plants with desired characteristics in integrated methodology under various national programs will provide a basis and opportunity to develop reliable biotechnology for functional purpose and create designer plants. There is an acute need to identify and characterize medicinal plants, that is, the chemistry of the active components, as they are the reservoir of the “medicine of the future.” Further development is essential to protect wild populations of different plant species with their inherent interspecific diversity. Biotechnological approaches, obviously, have found potential as an upgrade to conventional agriculture in the culture of plant tissue, the advanced manufacture of plant bioactive metabolites, in search of alternative production of useful medicinal compounds from plants. A. rhizogenes are stable and show high productivity in hormone-free culture conditions so can be used for the manufacture of derivatives. Certain modifications in culture conditions can induce growth and increased alkaloid production. Components of signal acceptance by plant cells, and guidelines for the differential expression of enzymes and genes are attractive areas in the investigation of the biosynthesis of routine products of nature, and plant cell culture would be a very suitable model framework for these examinations. It is necessary to know the pivotal biosynthetic enzymes that affect its regulation or its expression. Misuse of varietal cell strains for such examinations will serve up the possibilities of using cell suspension cultures for the bioproduction of metabolites by bioreactor cultivation, biotransformation, and immobilization, which needs further consideration. Furthermore, rhizosphere modeling requires the development of efficient lineage transformation protocols, which can be used to recover plants containing transgenes encoding enzyme(s) for the rate-limiting step(s) of biosynthetic pathways. This may guide for enhanced enzymatic(s) activity in vivo and higher bioproduction of pharmaceutically active secondary metabolites.
18. FUTURE PERSPECTIVES
With the increasing demands of the pharmaceutical industries, natural products are being utilized at large scale for the manufacture of drugs. The best about the natural formulations is that the patients develop no to very less side effects. Thus, enhancement of pharmaceutically significant plant secondary metabolite through Agrobacterium can serve as very efficient and promising tool for the plant tissue culturists. Thus, we intend to transform the elite varieties of Ashwagandha with A. rhizogenes and develop new tissue culture of plant developed elite plantlets for commercial farming. Later, these plants can be utilized by pharma companies for manufacturing of natural drugs for the treatment of ailments with very less or no side effects.
19. AUTHORS’ CONTRIBUTIONS
There are no conflicts of interest of the authorship.
20. FUNDING
No funding applicable in this work.
21. CONFLICTS OF INTEREST
There are no conflicts of interest of the authorship. The idea of manuscript and main manuscript was written by Dr. Manali Singh; while Kuldeep Jayant, Shruti Bhasin, Deep Chandra Suyal, and Dr. Sanjeev Agrawal helped in final editing.
22. ACKNOWLEDGMENT
We acknowledge G.B. Pant University of Agriculture and Technology, Pantnagar, and Invertis University, Bareilly, for providing the platform and necessary support and cooperation in preparing the manuscript.
23. ETHICAL APPROVALS
The work does not need any ethical appoval.
24. DATA AVAILABILITY
All the data pertaining to the manuscript has been provided in the manuscript.
25. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Hinsinger P, Marschner P. Rhizosphere-perspectives and challenges-a tribute to Lorenz Hiltner 12-17 September 2004-Munich, Germany. Plant Soil 2006;283:7-8. [CrossRef]
2. Singh M, Poddar NK, Singh D, Agrawal S. Foliar application of elicitors enhanced the yield of withanolide contents in
3. Jones D, Nguyen C, Finlay R. Carbon flow in the rhizosphere:Carbon trading at the soil-root interface. Plant Soil 2009;321:5-33. [CrossRef]
4. Mougel C, Offre P, Ranjard L, Corberand T, Gamalero E, Robin C,
5. Micallef SA, Channer S, Shiaris MP, Colon-Carmona A. Plant age and genotype impact the progression of bacterial community succession in the Arabidopsis rhizosphere. Plant Signal Behav 2009;4:777-80. [CrossRef]
6. Weisskopf L, Abou-Mansour E, Fromin N, Tomasi N, Santelia D, Edelkott I,
7. Kelgane SB, Salve J, Sampara P, Debnath K. Efficacy and tolerability of ashwagandha root extract in the elderly for improvement of general well-being and sleep:A prospective, randomized, double-blind, placebo-controlled study. Cureus 2020;12:7083. [CrossRef]
8. Singh B, Saxena AK, Chandan BK, Gupta DK, Bhutani KK, Anand KK. Adaptogenic activity of a novel, withanolide free aqueous fraction from the roots of
9. Verma N,Gupta SK, Tiwari S, Mishra AK. Safety of ashwagandha root extract:A randomized, placebo-controlled, study in healthy volunteers. Complement Ther Med 2021;57:102642. [CrossRef]
10. Davis L, Kuttan G. Effect of
11. Bhasin S, Singh M, Singh D. Review on bioactive metabolites of
12. Singh M, Shah P, Punetha H, Agrawal S. Varietal comparison of withanolide contents in different tissues of
13. Umadevi M, Rajeswari R, Sharmila Rahale C, Selvavenkadesh S, Pushpa R, Kumar KP,
14. Mills E, Cooper C, Seely D, Kanfer I. African herbal medicines in the treatment of HIV:Hypoxis and Sutherlandia. An overview of evidence and pharmacology. Nutr J 2005;4:19. [CrossRef]
15. Pandey MM, Rastogi S, Rawat AK. Indian traditional Ayurvedic system of medicine and nutritional supplementation. Evid Based Complement Alternat Med 2013;2013:376327. [CrossRef]
16. Kuo YT, Liao HH, Chiang JH, Wu MY, Chen BC, Chang CM,
17. Renu S, Manvi M, Sapna B. Evaluation of antibacterial potential of stem and bark of
18. Singh M, Shah P, Punetha H, Gaur AK, Kumar A, Agrawal S. Isolation and quantification of a potent anti cancerous compound, Withaferin A from the aerial parts of
19. Young JM, Kuykendall LD, Martínez-Romero E, Kerr A, Sawada H. A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of
20. Lima JE, Benedito VA, Figueira A, Peres LE. Callus, shoot and hairyroot formation
21. Ono NN, Tian L. The multiplicity of hairy root cultures:Prolific possibilities. Plant Sci 2011;180:439-46. [CrossRef]
22. Ozyigit II, Dogan I, Tarhan EA.
23. Keeling PJ, Palmer JD. Horizontal gene transfer in eukaryotic evolution. Nat Rev Genet 2008;9:605-18. [CrossRef]
24. De boer KD, Lye JC, Aitken CD, Su AK, Hamill JD. The A622 genein
25. Yamazaki Y, Kitajima M, Arita M, Takayama H, Sudo H, Yamazaki M,
26. John J. Therapeutic potential of
27. Datta A, Jain G, Avashthi H, Singh M, Agrawal S.Molecular docking of withanolides from
28. Choudhary D, Bhattacharyya S, Joshi K. Body weight management in adults under chronic stress through treatment with ashwagandha root extract:A double-blind, randomized, placebo-controlled trial. J Evid Based Complementary Altern Med 2017;22:96-106. [CrossRef]
29. Chandran U, Patwardhan B. Network ethnopharmacological evaluation of the immunomodulatory activity of
30. Abhyankar GA and Chinchanikar GS. Response of
31. Prakash J, Gupta SK, Dinda AK.
32. Gupta SK, Dua A, Vohra BP.
33. Chen WY, Chang FR, Huang, ZY, Chen, JH and Wu YC. Tubocapsenolide A, a novel Withanolide, inhibits proliferation and induces apoptosis in MDAMB-231 cells by thiol oxidation of heat shock proteins.
34. Ahmed BM, Akhter S, Aminual MD and Frazana SA.
35. Jayaprakasam B, Nair MG. Cyclooxygenase-2 inhibitory withanolides from
36. Sengupta P, Agarwal A, Pogrebetskaya M, Roychoudhury S, Durairajanayagam D, Henkel R. Role of
37. Dongre S, Langade D, Bhattacharyya S. Efficacy and safety of ashwagandha (
38. Wankhede S, LangadeD, Joshi K, Sinha SR, Bhattacharyya S. Examining the effect of
39. Mishra LC, Singh BB, Dagenais S. Scientific basis for the therapeutic use of
40. Dhuley JN. Nootropic-like effect of Ashawagandha (
41. Kaur P, Mathur S, Sharma M, Tiwari M, Srivastava KK, Chandra R. A biologically active constituent of
42. Gupta GL, Rana AC. PHCOG MAG:Plant review.
43. Choudhary D, Bhattacharyya S, Bose S. Efficacy and safety of ashwagandha (
44. Davis L, Kuttan G. Effect of
45. Kumar A, Kaul MK, Bhan MK, Khanna PK, Suri KA. Morphological and chemical variation in 25 collections of the Indian medicinal plant,
46. Chukwuma CI, Matsabisa MG, Ibrahim MA, Erukainure OL, Chabalala MH, Islam MS. Medicinal plants with concomitant anti-diabetic and anti-hypertensive effects as potential sources of dual acting therapies against diabetes and hypertension:A review. J Ethnopharmacol 2019;235:329-60. [CrossRef]
47. Agarwal AV, Gupta P, Singh D, Dhar YV, Chanda D and Trivedi PK. Comprehensive assessment of the gene involved in withanolide biosynthesis from
48. Rao SR, Ravishankar GA. Plant cell cultures, chemical factories of secondary metabolites. Biotechnol Adv 2002;20:101-53. [CrossRef]
49. Tripathi N, Shrivastava D, Ahmad Mir B, Kumar S, Govil S, Vahedi M,
50. Georgiev MI, Ludwig-Muller J, Alipieva K, Lippert A. Sonication-assisted
51. Ma LS, Hachani A, Lin JS, Filloux A, Lai EM.
52. Russell AB, Peterson SB, Mougous JD. Type VI secretion system effectors:Poisons with a purpose. Nat Rev Microbiol 2014;12:137-48. [CrossRef]
53. Mishra LC, Singh BB, Dagenais S. Scientific basis for the therapeutic use of
54. Shi Y, Lee LY, Gelvin SB. Is VIP1 important for
55. Willig CJ, Duan K, Zhang ZJ. Transcriptome profiling of plant genes in response to
56. Liu Y, Zhang Z, Fu J, Wang G, Wang J, Liu Y. Transcriptome analysis of maize immature embryos reveals the roles of cysteine in improving
57. Ara TALAT and Chaudhary AK. Study on efficacy of two strains (ATCC 15834 and MTCC 532) of Agrobacterium rhizogene on hairy root induction of
58. Gelvin SB.
59. Chaudhuri KN, Ghosh B, Tepfer D, Jha S. Genetic transformation of
60. Sudha CG, Seeni S. Establishment and analysis of fast growing normal root culture of
61. Giri A, Narasu ML. “Transgenic hairy roots:Recent trends and applications.“Biotechnol Adv 2000;18:1-22. [CrossRef]
62. Al-Hindawi MK, Al-Khafaji SH and Abdul-Nabi MH. Anti-granuloma activity of Iraqi
63. Choi HR, Choi JS, Han YN, Bae SJ and Chung HY. Peroxynitrite scavenging activity of herb extracts.
64. Pawar PK, Teli NP, Bhalsing SR, Maheshwari VL. Micropropagation and organogenetic studies in
65. Bandyopadhyay M, Jha S, Tepfer D. Changes in morphological phenotypes and withanolide composition of Ri-transformed roots of
66. Murthy HN, Dijkstra C, Anthony P, White DA, Davey MR, Power JB,
67. Hu ZB, Du M. Hairy root and its application in plant genetic engineering. J Integr Plant Biol 2006;48:121-7. [CrossRef]
68. Baburaj S and Gunasekaran K.
69. Barche S, Kirad SK and Sharma AK.
70. Wise AA, Fang F, Lin YH, He F, Lynn DG, Binns AN. The receiver domain of hybrid histidine kinase VirA:An enhancing factor for vir gene expression in Agrobacterium tumefaciens.
71. Hu X, Zhao J, Degrado WF, Binns AN. Agrobacterium tumefaciens recognizes its host environment using ChvE to bind diverse plant sugars as virulence signals.
72. Yuan ZC, Liu P, Saenkham P, Kerr K, Nester EW. Transcriptome profiling and functional analysis of Agrobacterium tumefaciens reveals a general conserved response to acidic conditions (pH 5.5) and a complex acid-mediated signaling involved in Agrobacterium-plant interactions.
73. Lai EM, Shih HW, Wen SR, Cheng MW, Hwang HH, Chiu SH. Proteomic analysis of Agrobacterium tumefaciens response to the vir gene inducer acetosyringone.
74. Atmakuri K, Cascales E, Christie PJ. Energetic components Vir D4, Vir B11 and Vir B4 mediate early DNA transfer reactions required for bacterial Type IV secretion. Mol Micrbiol 2004;54:1199-211. [CrossRef]
75. Gelvin SB.
76. Kalogeraki VS, Zhu J, Stryker JL, Winans SC. The right end of the vir region of an octopine-type Ti plasmid contains four new members of the vir regulon that are not essential for pathogenesis.
77. Magori S, Citovsky V.
78. Kumar A, Kaul MK, Bhan MK, Khanna PK, Suri KA. Morphological and chemical variation in 25 collections of the Indian medicinal plant,
79. Ozyigit II.
80. Abou-Douh AM. New Withanolides and Other Constituents from the Fruit of
81. Dewir YH, Chakrabarty D, Lee SH, Hahn EJ and Paek KY. Indirect regeneration of
82. Zhong JJ. Biochemical engineering of the production of plant-specific secondary metabolites by cell cultures.
83. Abraham A, Kirson I, Glotter E and Lavie D.Achemotaxonomical study of
84. Kim M, Ahn JW, Song K, Paek KH, Pai HS. Forkhead-associated domains of the tobacco NtFHA1 transcription activator and the yeast Fhl1 forkhead transcription factor are functionally conserved.
85. Chatterjee S, Srivastava S, Khalid A, Singh N, Sangwan RS, Sidhu OP
86. Chaurasiya ND, Gupta VK and Sangwan RS. Leaf ontogenic phase related dynamics of withaferin A and withanone biogenesis in ashwagandha (
87. Chauhan S, Joshi A and Jain D. RAPD Based Genetic Diversity Analysis in 25 Genotypes of
88. Bouchez D, Tourneur J. Organization of the agropine synthesis region of the T-DNA of the Ri plasmid from Agrobacterium rhizogenes. Plasmid. 1991;25(1):27–39. [CrossRef]
89. Ahuja A, Kaur D, Sharada M, Kumar A, Suri KA and Dutt P. Glycowithanolides accumulation in vitro shoot cultures of Indianginseng (
90. Le Flem-Bonhomme V, Laurain-Mattar D, Fliniaux MA. Hairy root induction of
91. Sivanandhan G, Dev GK, Jeyaraj M, Rajesh M, Muthuselvam M, Selvaraj N,
92. Liu Z, Park BJ, Kanno A, Kameya T. The novel use of a combination of sonication and vacuum infiltration in
93. Subramanyam K, Subramanyam K, Sailaja KV, Srinivasulu M, Lakshmidevi K. Highly efficient
94. Khanna H, Becker D, Kleidon J, Dale J. Centrifugation Assisted
95. Hiei Y, Ishida Y, Kasaoka K, Komari T. Improved frequency of transformation in rice and maize by treatment of immature embryos with centrifugation and heat prior to infection with