
1. INTRODUCTION
Breast cancer (BC), which has a high mortality rate and the capacity to spread, is the leading cause of death in women globally. Aside from the statistics, the condition is challenging to cure due to its heterogeneity [1]. When it comes to cancer, there are various mysteries, a major part of which is attributable to its tendency to spread. A big part of this is due to the active participation of extracellular vesicles, which affect the microenvironment while simultaneously causing intracellular modifications. Exosomes seem to be an active player in the development of metastatic illness, including cancer, among the extracellular vesicles [2]. This review will examine exosomes, their roles, and methods of generation in the context of cancer therapies and diagnostics, with an emphasis on BC in particular. This is done with participation in every stage of metastatic disease progression, examining it through the prism of Paget’s theory, resulting in issue solutions via detailed study [3]. Exosomes must be studied in depth since they play a crucial part in many of life’s key processes; hence, they must be thoroughly examined.
2. MOLECULAR CLASSIFICATION
Most research reveals that BC is a physiologically and phenotypically diverse group of illnesses with varying clinical and treatment response behaviors [2]. Only the morphological classification (nuclear grade, tubular grade, mitotic index, histological grade, and architectural characteristics) and clinical-pathological parameters (tumor size, lymph node involvement, and metastasis) are sufficient to predict the true behavior of breast tumor pathophysiology in this era of modern medicine [4]. As a result, past research concentrated on examining the molecular patterns of BC to classify these tumors or entities to aid in clinical care, the creation of epidemiological and functional analyses, and clinical performance [6]. As a result, these investigations indicated that there are BC subtypes with distinct gene expression patterns that represent individual phenotypes, illness prognosis, and systemic therapy planning. Based on extensive gene expression profile studies, four therapeutically important molecular subtypes were identified: luminal A, luminal B, enhanced HER2 (HER2+), and Triple Negative (TN) [7]. Genes associated with the expression of estrogen receptors (ER), progesterone receptors (PR), HER2 (Human epidermal growth factor receptor 2), and cell proliferation regulators are the key groupings of genes responsible for the segregation of molecular subtypes of BC (Ki-67). The Immunohistochemical (IHC) panel comprised these four biomarkers (ER/PR/HER2/Ki-67) has been shown to be effective and significant in the stratification of these molecular entities [1,9]. The ability to stratify BC into multiple entities that need different therapies and surveillance regimens, as well as a greater knowledge of the pathophysiological pattern and clinical prognosis, has revolutionized our perspective on the disease [Figure 1].
 | Figure 1: Molecular classification of breast carcinoma. [Click here to view] |
3. BIOLOGICAL FUNCTIONS OF EXOSOME
Exosomes are among the most recent bioactive molecules, with applications in cancer biology, phytochemistry, and genetic engineering. It is a vesicle found in a variety of biological fluids and the extracellular matrix of a variety of cell microenvironments, making it a significant molecule in the biological system. Exosomes are nanovesicles found in the cytoplasm of both prokaryotes and eukaryotes that have lately become a major study issue due to their involvement in various signaling cascades. They range in size from 50 to 140 nm and are found in a variety of biomolecules [9]. In exocytosis, the membrane is a phospholipid bilayer that merges with the plasma membrane. PGRL (PG regulatory-like protein) and LAMP1/2 (Lysosomal associated membrane proteins) are two transmembrane proteins and annexins found in the membrane. Lipid-related proteins, such as flotillin and cholesterol, are found in the cell membrane. Because they can be used to tell different types of cells apart, they could be good identifiers [11].
Exosomes have a crucial role in metastasis and cancer progression as part of the cell microenvironment, making them an important aspect of cancer genomics and stem cell research. They were initially found after the removal of transferrin receptors during reticulocyte development. A large percentage of the plasma membrane was regenerated as a consequence of having a significant number of alterations near the end of this process during the transition to becoming a mature erythrocyte [12]. Because of the existence of many vesicles inside them, these molecules were dubbed multivesicular bodies, and the vesicles were renamed intraluminal endosomal vesicles, which are now known as exosomes. While they play a role in numerous molecular pathways, including T cell specificity and neurological system presence [13], their involvement in the spread of BC will be explored in this work.
4. BIOGENESIS OF EXOSOMES
Exosomes are formed primarily along a basic route that is controlled by many different signals transmission channels. Both begin with primitive endosomes that evolve into mature endosomes known as MVBs (multivesicular bodies). ILVs are being created at the same time that this operation is taking place. Transmembrane proteins are injected into the curved membrane, while cytoplasmic material is contained inside the vesicles. A sorting complex is needed for deciphering transfer ESCRT (endosomal sorting complex required for transport) [14]. The interaction of all of these components causes vesicle bulging and cargo sorting through ubiquitin regulation, which is synthesized by the ESCRT [Figure 2]. The process begins with ESCRT-0, which recognizes ubiquitin marks on the membrane. The limiting membrane then shrinks, and the budding process is triggered by ESCRT-I/II and ESCRT-III. The ATPase Vps4 drives the complex. This process may also take place in the absence of ubiquitin and syntenies. MVBs then fuse with the plasma membrane, releasing freshly generated exosomes. Exosome secretion is controlled by Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Ca2+/Ca2+ dependent Rab-binding protein signaling pathway It has been shown that leptin, followed by leptin target shock protein, may induce tsg101, the regulator for the development of the ESCRT-I complex, in BC cells [16]. However, this process may also occur in the absence of ESCRT, revealing a mechanism that does not need complex creation. This was first observed in oligodendrocytes when the ESCRT pathway was inhibited. The process occurred after the depletion of sphingomyelinase as it involved the formation of proteolipid protein. While not much is known about the ESCRT independent pathway, it can be called simpler as no new involving it were detected. Exosomes, like CD9 and CD82, have a lot of tetracaine in them, which make beta-catenin, which is important for WNT (Wingless and Int-1) signaling, come out more [16].
 | Figure 2: Entry mechanism of exosomes. [Click here to view] |
5. CARRIERS OF EXOSOMES
In the case of BC, exosomes mostly include proteins, enzymes, and nucleic acids. There are various forms of RNA (ribonucleic acid) in nucleic acids, including mi RNA, mRNA (messenger RNA), and tiny quantities of piwi RNA, including RNA, transfer RNA, and ribosomal RNA. GTPases (Guanosine triphosphatases) and annexin proteins, for example, are found in exosomes and interact with target cells. As previously stated, exosomes are involved in practically every phase of cancer prognosis. This encompasses both early cell proliferation and metastasis [Figure 3].
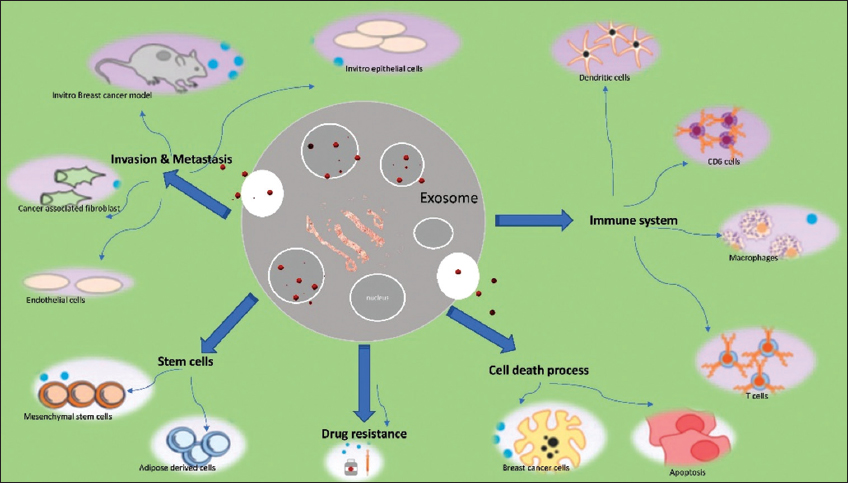 | Figure 3: Role of exosomes in breast cancer. [Click here to view] |
6. EXOSOMES IN BREAST CANCER GROWTH AND METASTASIS
Exosome components play a key role in the carcinogenesis process. This is accomplished by inducing cancer cell growth-boosting factors [17]. This may be accomplished by activating oncogenic proteins such as RAS (Rat sarcoma virus) and EGFR (Epidermal growth factor receptor). Meanwhile, miR-10b and miR-21 are found in the exosomes of BC cells and serum of cancer patients due to dicer-mediated deletion of the phosphatase and tension homolog on chromosome ten (PTEN) and homeobox D10 (HOXD10) genes [19]. Meanwhile, miR-100 has transcription regulator effects that are controlled by the appearance of EMT (Epithelial-mesenchymal transition), as well as tumor-promoting gene regulation. Then, docetaxel-resistant miRNAs (miR-26a, miR-24, and miR-271) play a role in activating MAPK (Mitogen-activated protein kinase) [18] [Figure 4].
 | Figure 4: Tumor progression in breast. [Click here to view] |
Neoangiogenesis is the process by which a malignant tumor creates a food supply for itself via rearrangement of existing capillaries or the formation of new capillaries, enabling the tumor to grow further. This is one of the markers of the development of cancer. It is a mechanism regulated by an intracellular signaling cascade that includes exosomes [19]. Exosomes and Annexin II, a Ca2+-dependent phospholipid-binding protein, are known to promote angiogenesis. This is accomplished by the use of vascular endothelial growth factors (VEGF) and its isoforms VEGFA, as well as the receptors. This pathway is necessary for vascular supply synthesis. Exosomes deliver cargo comprising CD106, 49d, and tetracaine 6, all of which are responsible for triggering the VEGF signal. The protein is in charge of active cell division, migration, and maturation of endothelial cells [21]. Sphingomyelinase 2 controls neo-angiogenesis through modulating miRNA. miRNAs, such as miR-210, are delivered to endothelial cells, enhancing their capacity to migrate and create capillaries [22] [Figure 5].
 | Figure 5: Exosome and its cargo in breast cancer. [Click here to view] |
Exosomes, on the other hand, are implicated in the mechanism by which their cargo triggers AKT/PI3k/mTOR (Protein Kinase B) signaling, resulting in angiogenesis. Several chemicals, including IGF (Insulin-like growth factor), EGF, VEGF, IL-6 (Interleukin), leptin, and insulin, activate this route, and their trafficking is controlled by exosomes. Hypoxia is the primary force for angiogenesis. Exosomes are involved in the process of intercellular communication with the tumor microenvironment, which aids in the dissemination of the signal that tumors are hypoxic [20]. MiR-210 is released by exosome HIF-1 (Hypoxia-inducible factor 1) alpha release, regulating angiogenesis. Exosomes may enter the receptor cell through lipid rafts, clathrinid coated pits and caveolae, receptor-mediated endocytosis, micropinocytosis, and phagocytosis, all of which result in the production of intracellular signals. Phagocytosis is carried out by the actin cytoskeleton and is accomplished by actin polymerization. PI3K is crucial for exosome entrance, and dynamin 2 may regulate phagocytosis through a GTPase for clathrinid-mediated receptor-mediated endocytosis [24]. Endocytosis is occasionally reliant on caveolae, a substructure of glycolipid rafts. Clathrinid independent endocytosis, for example, is dependent on cholesterol present in lipid rafts. Epithelial to mesenchymal transition Degradation of matrix-metalloproteinases may be connected to cancer’s potential for metastasis. MCF-20A (Mitochondrial carrier family protein) BC, for example, is produced from a highly metastatic MDA-MB 231 cell line through enhanced MMP (Matrix metalloproteinase) and MMP 9 secretion. miRNA, which is released by exosomes, has the potential to be a biomarker for several forms of BCs. EMT may be induced by molecules such as TGF-beta. EVs that come from HER2 BC, for example, are resistant to trastuzumab-induced cell death because they exocytose TGF-beta 1 and PDL1.
Epithelial-mesenchymal transition is a process of change or motility as well as shape that follows the mechanism of epithelial cell transformation into mesenchymal cells. Similarly, to angiogenesis, EMT is mediated by the sensing of hypoxia [25]. This is a transition of basic cancer cells into motile metastasizing cells. It has considerable flexibility, with epithelial cells oriented apicobasal and a large degree of extracellular matrix attachment, but mesenchymal cells do not adhere to other cells and are extremely migratory. There are significant molecular variations between the cells, such as epithelial cells having attachment e cadherin, while mesenchymal cells contain the adhesin protein fibronectin or N cadherin [26].
Exosomes are responsible for most of the epithelial to mesenchymal transition, as well as alterations in the microenvironment. They transport cargo that contains many functional compounds that protect motile mesenchymal cells. This is because mesenchymal cells are designed to travel through the circulation, encountering immune cells, increased blood pressure, and anoikic as a result of this metamorphosis. Exosomes synthesize proteins that help cells survive while undergoing metamorphosis [27]. They are responsible for the production of chemicals such as VEGF, EGF, and fibroblast growth factors. Exosomes from fibroblasts increase autocrine WNT signaling in BC. It can both positively and negatively regulate carcinogenesis by maintaining and proliferating leukocytes while also boosting immunological tolerance and decreasing anti-tumor immunity. It is important in EMT because it increases the survival of cancer stem cells. The route, according to studies, may be canonical (beta-catenin dependent) or non-canonical (beta-catenin independent). Both pathways share Frizzled receptors, which activate the cytoplasmic protein Dash, which works upstream of beta-catenin to induce phosphorylation when attached to Lerp. In normal settings, beta-catenin induces the system by transporting a pro-apoptotic complex to the nucleus, resulting in T cell-specific factors. This route is activated with the aid of a Cs81-positive exosome. It is also proposed that the route is anti-apoptotic, helping cell cycle elements [28]. In addition, exosomes are also important for mesenchymal cell development, which in the case of BC, leads to the production of a myofibroblast phenotype [29].
When cancer progresses to metastatic illness, the death rate skyrockets, necessitating a plethora of various sorts of therapy. According to Paget’s and Fiddler’s seed and soil theories, tumor cells survive metastasis through interacting with the tumor microenvironment. This was proposed based on the finding that some tumors had a propensity to spread to certain locations [3]. For example, BC often spreads to the bones, brain, liver, and lungs. This implies that the tumor or seed can only survive in inhospitable soil or a favorable microenvironment. When the tumor cells are circulating, the theory comes into play. They have a pre-metastatic niche that is only active in particular microenvironments. Intravasation, or homing, happens when the seed comes into contact with a favorable microenvironment. Tumor-secreted factors play an important role in metastasis. Exosomes are included in this. As previously stated, this contributes to metastasis through hypoxia-inducible factor 1 alpha. Exosomes are crucial for transforming the main soil into a favorable habitat for metastasis. This comprises MCF7 and MDA-MB secretion, which induces mesenchymal cell development into myofibroblasts. Later on, the exosome produces chemokines and participates in neutrophil infiltration, leading to metastasis [30]. Furthermore, for the process to work, endothelial connections must be destroyed, which is accomplished by miR-939, which targets particular junctions, HUVECs (Human umbilical vein endothelial cells), which are responsible for enhanced permeability of a monolayered membrane. Simultaneously, aspartate beta-hydroxylase secretes pro-invasive endosomes that target Notch receptors, regulators, ligands, and MMPs, boosting invasion capabilities. Exosomes are also responsible for the transfer of exosomes such as miR-105 as a biomarker for metastasis and as a signal for early diagnosis. Only in particular microenvironments does cancer cell homing occur after dispersion. Immune cells and platelets aid the CTC (circulating tumor cell) while it is in circulation. It is worth noting that cancer cells have more EVs than normal cells. Metastatic cells include many proteins, including metatherian and ceruloplasmin. MMPs, which are responsible for extracellular matrix disintegration, are also renowned for their metastatic capacities. Exosomes are capable of helping malignancies evade the immune system, which is one of the characteristics of BC. They can elude T cells, macrophages, T regulatory cells, and dendritic cells. Members of NK group 2 [32] include NK cells, NKT cells, yeti delta t cells, and CD8+ alpha beta t cells, which all perform cytotoxic killing before activating T cells. When exposed to BC-derived exosomes, the efficiency of lymphocyte NKG2D receptor expression, as well as CD8+T, is greatly decreased. Exosomes containing miRNAs are capable of regulating tumor-associated macrophages, which help in angiogenesis, in addition to immunosuppression. This is accomplished by miR-23, which alters IL-4-mediated monocyte-derived macrophages as well as miR-565 and miR-660 [33].
The seed’s qualities are determined by the main soil and substances released by the tumor. It has been proposed that tumorigenic and stem cell characteristics are produced in cells during EMT. Stem cells are produced by fibroblasts, have a mesenchymal origin, and can migrate to other tissues [34]. Exosomes are present in at least mesenchymal stem cells from stem cells that exhibit features that inhibit angiogenesis. The stemness of the cell is attributed to components such as HMGB1 (High Mobility Group Box 1), which is regulated in cancer-associated fibroblasts through the HMGB1/TLR4 (Toll-like receptor-4) signaling pathway. The cell sometimes undergoes metastatic dormancy, which is only restored when oncogenic factors proliferate in the microenvironment. In the G0 phase, this is a condition of quiescence, not senescence. Cell death and suppressed cell activity caused by hTERT (human telomerase reverse transcriptase) extravasation also prevented cell death and suppressed cell activity [35]. The bones have the greatest chance of colonization, followed by the lungs, liver, and brain. BC cells carrying miR-105 exosomes have been demonstrated to cross the barrier between tight junctions and protein metastases to the lungs and brain [36]. Extracellular matrix breakdown and e-Cadherin-mediated transition from mesenchymal to epithelial cells are involved in lung cell invasion [36]. Meanwhile, colonization and expansion are the last stages of metastasis. One of the characteristics of metastatic illness is altered metabolism, which in the case of BC is accomplished with the assistance of miR-122, which examines glucose absorption by non-tumor cells such as brain astrocytes and lung fibroblasts [37]. He prepares the pre-metastatic niche by improving the soil’s congeniality. Meanwhile, the exosomes produced by the BC encourage enhanced cell division at the seeding sites. The living conditions in the pre-metastatic niche are regulated by several proteins and RNAs. miR-122, for example, modulates nutritional ability and is accompanied by long non-coding RNAs [38]. In certain instances, circulating tumor cells re-invade the tumor from the left side, a process is known as self-seeding [39]. There was also the idea that cells make different exosomes that have organ-specific integrins in them, which makes a good tumor microenvironment [40].
7. DRUG RESISTANCE AND EXOSOMES
While treating cancer cells with exosomes is difficult enough due to tumor heterogeneity, another issue linked to cancer cells with exosomes is medication resistance. When one of the hallmarks of cancer is inhibited by the use of new, secondary pathways, drug resistance arises. This may happen as a result of epigenetic repression of tumor suppressor genes and proteins, a rise in the incidence of cancer stem cells, and limited drug penetrance [41]. Radiation treatment resistance arises when the remaining cells release exosomes, promoting cell proliferation. Because exosomes are responsible for the spread of drug and radiation resistance, they are unquestionably important when determining the course of treatment. While complete elimination of exosomes from the body would not be feasible, given that they are produced based on intracellular signaling, isolation of exosomes from the plasma can be quite helpful in diagnosing the nature of cancer and hence in deciding on a course of treatment. In the case of BC, which is heterogeneous in some cases, this process will have to be done in specific regions, based on the nature of the cells in that particular division. This is because while certain parts of cancer may be resistant to a certain drug, certain parts may be vulnerable to the drug. While the decision for the general course of treatment for heterogeneous cancers is quite challenging, using exosomes for diagnosis, for instance, resistance to trastuzumab, for example, is mediated by exosome proliferation in HER-2 positive cell lines. This is linked to cancer aggressiveness [42].
8. CLINICAL POTENTIAL
Exosomes, which were formerly thought to be cellular trash, are now recognized as essential biomarkers. As is well known, BC is a disease with a wide range of manifestations. This implies that the tumor has a wide range of cells, each with its own set of biomarkers, making it a difficult condition to treat. Exosomes are a crucial aspect of the microenvironment because they are responsible for cargo distribution and, since they are pleiotropic, include various variations that allow cells to be differentiated depending on their milieu [45]. Exosome cargo, including miRNA and proteins, is responsible for the regulation of the tumor microenvironment. Hence, studying the exosomes and their cargo can certainly be helpful in terms of diagnosis. This can include the general nature of the tumor, that is, HER2 expression as well as resistance to certain treatments, that is, trastuzumab resistance. Exosomes, which are an essential component of all fluid microenvironments in the body, are subjected to a liquid biopsy. The fluid contains both circulating tumor cells and extracellular vesicles. Exosomes may be released in a variety of ways. They can prove to be an even more efficient diagnostic tool as their contents change on the basis of the exact stage of cancer. Being an efficient part of all the phases of cancer, EMT transition, intravasation, extravasation, dormancy, stem-like nature, as well as characteristics exhibited during the initial phases of carcinogenesis, they can be a more efficient tool as opposed to physical evidence. Targeting exosomes for treatment can be a way to use the biochemical properties of cancer. It can prove to be an equally efficient method of diagnosis, if not more precise. This includes irradiation-induced senescence, cisplatin exposure, long-term hypoxia, and Tunica mien exposure [46]. Meanwhile, there have already been several advances made in the field of exosome usage for the treatment of BC. This mainly includes their role in the process of drug delivery. This includes cargo like miRNA and proteins. Delivery in exosomes can increase the uptake efficiency significantly. Further studies can be conducted to find the ideal source of exosomes for this purpose. Improvements in the delivery of the drug into the exosome, meanwhile, can also be improved, to further improve the process of drug delivery. Being an efficient part when it comes to treatment and a crucial part when it comes to diagnosis, exosomes certainly have significant clinical potential to study as well as treat breast cancer.
8.1. Applications of Exosomes in the Diagnosis with Breast Cancer
As the significance of exosomes in the process of treatment and diagnosis has been established, some examples of the application of exosomes in the process should also be discussed [Table 1]. Several proteins are found on the surface of surface exosomes. Tetracaines, which are abundant in exosomes, are among them. These proteins are transmembrane and signaling proteins found in the cytoplasm. CD9 works with the metalloprotease ADAM10 (A disintegrant and metalloproteinase-10), the annexin-1 protein, and the heat shock protein HSP70 [47]. The quantity of malignant tissue was directly related to the level of tetracaine CD82. CD24, fibronectin, and endothelial locus 1 were all prominent epithelial cell markers produced by circulating exosomes. HER-2 is another significant protein in certain instances of BC [42]. Meanwhile, glypican-1, a cancer-derived proteoglycan, is found on the surface of malignant cells, which is crucial in instances of BC [43]. In contrast to the control, CD47 expression was lower in BC [44].
Table 1: Various applications of exosome protein and its origin.
| Exosomal protein | Body fluid/origin | Isolation/detection method | Application | References |
|---|---|---|---|---|
| ADAM10, tetraspanin annexin 1 | BC cell line supernatants, serum, pleural effusions | Western blot | Diagnosis | [56] |
| CD24 | Serum | Sucrose gradient and ultracentrifugation | Early diagnosis | [57] |
| Del 1 | Plasma | ELISA (Enzyme-linked immune sorbent assay) | Diagnosis/monitoring | [58] |
| fibronectin | Plasma | ELISA | Early diagnosis | [58] |
| HER-2 | Serum BC cell line, culture supernatants | Bradford assay and ultracentrifugation | Trastuzumab Tumor aggressiveness and resistance | [42] |
| Annexin A2 | BC cell line supernatant | Atomic force, microscopy | Brain and lung metastasis | [22] |
| miR-21 | Plasma | Nano oligonucleotide probe | Diagnosis | [48] |
| miR-372 | Serum | Quantitative Taq Man microRNA assays | Diagnosis | [50] |
| miR-373 | Serum | Quantitative Taq Man microRNA assays | Diagnosis of triple-negative BC | [50] |
| miR-1246 | Plasma | RT-qPCR (real time-quantitative polymerase chain reaction) | Diagnosis | [49] |
| miR-105 | MCF-10A and MDA MB 231 cell culture | RT-PCR, western blot analysis | Diagnosis | [36] |
| lncRNA GAS5 | MDA-MB 231 cell culture supernatants | RT-qPCR | Marker of apoptosis induction | [51] |
| miR-233 | Human macrophages | RT-qPCR | Promotes BC invasion | [52] |
| ADAM10 | Fibroblast | Antibodies and qRT-PCR primers | BC cell mobilization and proliferation | [54] |
Meanwhile, microRNA, another essential identifying component, is part of the payload. These are non-coding, single-stranded RNAs that are responsible for post-translational regulation of their target mRNA and have three untranslated regions. According to studies, Mir–13070-3p, Mir–1246, Mir–6861-5p, miR–4634, and miR–6875-5p show good specificity, accuracy, and sensitivity in diagnosing BC. This does not include miR-21 [48] and miR-1246 [49], which are found in all exosomes. There are various key miRNAs, including miR-372, which is responsible for cyclin g2 expression in BRCA (Breast Cancer gene 1) type tumors, and miR-105, which represents the premetastatic to the metastatic stage [36], miR-373 [50], and others. In exosome miRNA, certain levels of the miRNA are also responsible for drug resistance. This feature is one of the reasons why BRCA is one of the worst malignancies. This comprises miR-100, miR-30a, and miR-222, all of which are chemosensitive.
Aside from miRNA, various other RNAs may be delivered by exosomes. Long non-coding RNA participates in a variety of activities, including chromatin architecture, protein translation, mRNA turnover, and gene transcription. The genesis of cancer is caused by dysregulation of this. Long non-coding RNAs also have a role in the spread of cell signals, which helps to control the microenvironments of surrounding malignancies. In this case, GAS5 [50] is an example because it inhibits cell division and promotes apoptosis [38].
In the meantime, numerous RNAs have been found in non-BRCA cell-derived exosomes [51]. This contains various biomarkers, such as miR223 (which is generated from macrophages and is cultured alongside BrCa cells and is responsible for monitoring the invasiveness of the BRCA cells [52]. Similarly, cancer-associated fibroblasts stimulate cancer cell motility and intravasation through ADAM10-rich exosomes, which are a key component of cancer signaling. This molecule is hypothesized to be an inhibitor of the metalloproteinase family, which is important in extracellular matrix maintenance CAFs may also communicate between cells, influencing BRCA development [54]. It is possible to figure out the type of cancer before you start treatment by looking at all of the exosome material, which helps because biomarkers are also in the right place [55].
The disadvantage of employing exosomes as a diagnostic tool for BC is that they have a limited life span outside the cell and are easily contaminated by extracellular vesicles [60]. Their separation is possible with the aid of 100,000g before sucrose gradient centrifugation. Following this, the cell has all forms of exosomes [59]. However, in the future, microfluidics may provide more efficient ways of isolating exosomes [61]. There is a device called the Ex-Chip that is good at collecting and labeling exosomes with a CD6 antibody and fluorescein [62].
8.2. Exosomes and BC Prognosis
Due to their versatility and pleiotropy, exosomes have the potential to be effective drug delivery vehicles. They may be utilized to distribute peptides as well as RNA variations because of their stability and excellent delivery efficiency. On the other hand, both tumor and non-tumor cells release a range of cytokines, growth factors, adhesion molecules, and extracellular matrix proteins, which mediate cell-to-cell communication within the tumor microenvironment and provide a favorable setting for cancer cell development and survival. A plethora of data has been gathered on the function that these chemicals and cell types play in cancer invasion and metastasis. As a result, evidence of their participation as new players in the crosstalk between cancer and normal cells in the tumor microenvironment is beginning to accumulate. Exosomes may act in an autocrine or paracrine way to enhance tumor-induced immune suppression, angiogenesis, or the creation of premetastatic niches [63]. Luga et al. have shown that cancer-associated fibroblasts (CAFs) produce exosomes that increase BC cell protrusive activity and motility in an autocrine manner through Wnt-planar cell polarity (PCP) signaling. Their findings, in particular, reveal a complex intercellular communication pathway in which CAF-secreted exosomes are internalized, modified, and loaded with Wnt11 into BC cells; exosomes are then released to activate an autocrine PCP signaling in BC cells, promoting cancer cell migration and invasive behavior [64]. Furthermore, Zhu et al. recently showed that Mesenchymal stem cell (MSC) released exosomes are capable of robustly activating VEGF pathways in tumor cells through activation of Erk1/2 kinases, resulting in improved angiogenesis and tumor development in vivo [65]. Peinado et al. recently demonstrated that cancer-derived exosomes modulate the crosstalk between malignant cells and the bone marrow microenvironment; they reported, for the first time, that metastatic melanoma cells release exosomes capable of “educating” bone marrow progenitors, inducing mobilization that supports tumor vasculogenesis, invasion, and metastasis via activation of the Mesenchymal-epithelial transition factor (MET) receptor tyrosine kinase. They discovered that MET-activated signaling proteins are expressed in highly metastatic melanoma derived-exosomes, and that the transfer of the exosomal receptor tyrosine kinase MET from tumor derived-exosomes to bone marrow progenitor cells increases metastasis in vivo. These findings imply that trained bone marrow cells maintain their phenotypic state following engraftment into a new host [66]. Many studies have shown that exosomes that come from cancer cells play a role in activating signal transduction pathways that are important for cancer cell growth and survival.
Exosome-mediated treatment has been used to treat diseases such as cerebral ischemia. This is accomplished using curcumin-loaded exosomes. Natural compounds are most effective when combined with exosomes. Polyphenols, for example, may be utilized to lower the risk of and treat certain types of cancer. Exosomes are often isolated using exosomes from stem cells, which have a higher quantity of exosomes and the capacity to expand in vivo settings, as well as a high ability to differentiate and are immune system free [67]. They may, for example, be employed to limit the expression of miR-142-3p and miR-150, resulting in increased anti-tumor immunity and apoptosis. Several additional chemicals, such as shikonin, may be delivered to the pre-adipocyte milieu to inhibit the growth of tumor cells [68]. Meanwhile, removing exosomes may prove to be an effective technique for slowing the spread of some cancers. For example, in the case of HER-2 positive tumors, removing HER-2 positive exosomes may prevent the development of HER-2 positive BC [69]. Inhibition of miR-1246 and CCNG2 mRNA could be used as therapeutic targets for BC, which could slow tumor growth and treatment resistance [70-72].
8.3. Future for Exosome Targeting in Cancer Diagnosis and Prognosis
Improvements in techniques for isolating exosomes, most likely from stem cells, and improved diagnostic and therapeutic treatment of BC.
Because cancer cells are diverse, a greater knowledge of cargo packing is required, as well as a better understanding of early detection.
Cell lines and animal models have improved diagnostics and medicinal applications.
Improved knowledge of biomarkers and their responses to standard therapy
9. CONCLUSION
BC has undoubtedly been a tough illness to treat due to its heterogeneity, high likelihood of recurrence in roughly 5 years, and high metastasis rate. One approach to addressing metastatic illness and BC is to get a better knowledge of exosomes, which are important participants in intercellular communication. When we investigate the origins and activities of exosomes, we may discover several diagnostic and therapeutic uses for extracellular vesicles. This involves knowledge of the cargo in exosomes and its influence on the tumor’s microenvironment. Furthermore, the categorization of BC based on nature may aid in the development of a therapeutic technique for the condition. Analyzing the role of exosomes in each stage of metastasis may aid in the identification of a location for early detection and effective therapy. Exosomes may be used in the future to help detect and treat cancer. They could also be used as biomarkers or delivery vehicles for drugs.
10. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
11. FUNDING
No funding received for this particular study.
12. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
13. ETHICAL APPROVALS
This particular study has been approved by University ethical committee.
14. DATA AVAILABILITY
No previous data available for this study.
15. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Fang J, Chen F, Liu D, Gu F, Chen Z, Wang Y. Prognostic value of immune checkpoint molecules in breast cancer. Biosci Rep 2020;40:BSR20201054. [CrossRef]
2. Pentel DM, Gould SJ. Exosomes. Ann Rev Biochem 2019;88:487-514. [CrossRef]
3. Akhtar M, Haider A, Rashid S, Al-Nabet AD. Paget's “Seed and Soil“theory of cancer metastasis:An idea whose time has come. Adv Anat Pathol 2019;26:69-74. [CrossRef]
4. Lee G, Romo Bucheli DE, Madabhushi A. Adaptive dimensionality reduction with semi-supervision (AdDReSS):Classifying multi-attribute biomedical data. PloS One 2016;11:e0159088. [CrossRef]
5. Eliyatk?n N, Yalç?n E, Zengel B, Akta?S, Vardar E. Molecular classification of breast carcinoma:From traditional, old-fashioned way to a new age, and a new way. J Breast Health 2015;11:59-66. [CrossRef]
6. Harbeck N, Penault-Llorca F, Cortes J. Breast cancer. Nat Rev Dis Primers 2019;5:66. [CrossRef]
7. Rakha EA, Reis-Filho JS, Baehner F, Dabbs DJ, Decker T, Eusebi V, et al. Breast cancer prognostic classification in the molecular era:The role of histological grade. Breast Cancer Res 2010;12:207. [CrossRef]
8. Heng YJ, Lester SC Tse GM, Factor RE, Allison KH, Collins LC, et al. The molecular basis of breast cancer pathological phenotypes. J Pathol 2017;1:375-91. [CrossRef]
9. Sheffield BS, Kos Z, Asleh-Aburaya K, Wang XQ, Leung S, Gao D, et al. Molecular subtype profiling of invasive breast cancers weakly positive for estrogen receptor. Breast Cancer Res Treat 2016;155:483-90. [CrossRef]
10. Nagao T, Kinoshita T, Hojo T, Tsuda H, Tamura K, Fujiwara Y. The differences in the histological types of breast cancer and the response to neoadjuvant chemotherapy:The relationship between the outcome and the clinicopathological characteristics. Breast 2012;21:289-95. [CrossRef]
11. Jeppesen DK, Fenix AM, Franklin JL, Higginbotham JN, Zhang Q, Zimmerman LJ, et al. Reassessment of exosome composition. Cell 2019;177:428-45. [CrossRef]
12. Stevens-Hernandez CJ, Bruce LJ. Reticulocyte maturation. Membranes (Basel) 2022;12:311. [CrossRef]
13. Kalluri R. The biology and function of exosomes in cancer. J Clin Investig 2016;126:1208-15. [CrossRef]
14. Baietti MF, Zhang Z, Mortier E, Melchior A, Degeest G, Geeraerts A, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol 2012;14:677-85. [CrossRef]
15. Giordano C, Gelsomino L, Barone I, Panza S, Augimeri G, Bonofiglio D, et al. Leptin modulates exosome biogenesis in breast cancer cells:An additional mechanism in cell-to-cell communication. J Clin Med 2019;8:1027. [CrossRef]
16. Chairoungdua A, Smith DL, Pochard P, Hull M, Caplan MJ. Exosome release of b-catenin:A novel mechanism that antagonizes Wnt signaling. J Cell Biol 2010;190:1079-91. [CrossRef]
17. Jabbari N, Akbariazar E, Feqhhi M, Rahbarghazi R, Rezaie J. Breast cancer-derived exosomes:Tumor progression and therapeutic agents. J Cell Physiol 2017;235:6345-56. [CrossRef]
18. Ahmed SF, Das N, Sarkar M, Chatterjee U, Chatterjee S, Ghosh MK. Exosome-mediated delivery of the intrinsic C-terminus domain of PTEN protects it from proteasomal degradation and ablates tumorigenesis. Mol Ther 2015;23:255-69. [CrossRef]
19. Chen WX, Cai YQ, Lv MM, Chen L, Zhong SL, Ma TF, et al. Exosomes from docetaxel-resistant breast cancer cells alter chemosensitivity by delivering microRNAs. Tumor Biol 2014;35:9649-59. [CrossRef]
20. Soboci?ska AA, Czarnecka AM, Szczylik C. Mechanisms of angiogenesis in neoplasia. Postepy Hig Med Dosw 2016;70:1166-81.
21. Gopal SK, Greening DW, Hanssen EG, Zhu HJ, Simpson RJ, Mathias RA. Oncogenic epithelial cell-derived exosomes containing Rac1 and PAK2 induce angiogenesis in recipient endothelial cells. Oncotarget 2016;7:19709-22. [CrossRef]
22. Maji S, Chaudhary P, Akopova I, Nguyen PM, Hare RJ, Gryczynski I, et al. Exosomal annexin II promotes angiogenesis and breast cancer metastasis. Mol Cancer Res 2017;15:93-105. [CrossRef]
23. Tadokoro H, Umezu T, Ohyashiki K, Hirano T, Ohyashiki JH. Exosomes derived from hypoxic leukemia cells enhance tube formation in endothelial cells. J Biol Chem 2013;288:34343-51. [CrossRef]
24. Kruger S, Abd Elmageed ZY, Hawke DH, Wörner PM, Jansen DA, Abdel-Mageed AB, et al. Molecular characterization of exosome-like vesicles from breast cancer cells. BMC Cancer 2014;14:44. [CrossRef]
25. King HW, Michael MZ, Gleadle JM. Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 2012;12:421. [CrossRef]
26. Gross JC, Chaudhary V, Bartscherer K, Boutros M. Active Wnt proteins are secreted on exosomes. Nat Cell Biol 2012;14:1036-45. [CrossRef]
27. Dovrat S, Caspi M, Zilberberg A, Lahav L, Firsow A, Gur H, et al. 14-3-3 and b-catenin are secreted on extracellular vesicles to activate the oncogenic Wnt pathway. Mol Oncol 2014;8:894-911. [CrossRef]
28. Lin R, Wang S, Zhao RC. Exosomes from human adipose-derived mesenchymal stem cells promote migration through the Wnt signaling pathway in a breast cancer cell model. Mol Cell Biochem 2013;383:13-20. [CrossRef]
29. Luga V, Zhang L, Viloria-Petit AM, Ogunjimi AA, Inanlou MR, Chiu E, et al. Exosomes mediate stromal mobilization of autocrine wnt-pcp signaling in breast cancer cell migration. Cell 2012;151:1542-56. [CrossRef]
30. Harris DA, Patel SH, Gucek M, Hendrix A, Westbroek W, Taraska JW. Exosomes released from breast cancer carcinomas stimulate cell movement. PLoS One 2015;10:e0117495. [CrossRef]
31. Ribelles N, Santonja A, Pajares B, Llácer C, Alba E. The seed and soil hypothesis revisited:Current state of knowledge of inherited genes on prognosis in breast cancer. Cancer Treat Rev 2014;40:293-9. [CrossRef]
32. Wen SW, Sceneay J, Lima LG, Wong CS, Becker M, Krumeich S, et al. The biodistribution and immune suppressive effects of breast cancer-derived exosomes. Cancer Res 2016;76:6816-27. [CrossRef]
33. Rong L, Li R, Li S, Luo R. Immunosuppression of breast cancer cells mediated by transforming growth factor-b in exosomes from cancer cells. Oncol Lett 2016;11:500-4. [CrossRef]
34. Baroni S, Romero-Cordoba S, Plantamura I, Dugo M, D'Ippolito E, Cataldo A, et al. Exosome-mediated delivery of miR-9 induces cancer-associated fibroblast-like properties in human breast fibroblasts. Cell Death Dis 2016;7:e2312. [CrossRef]
35. Riches A, Campbell E, Borger E, Powis S. Regulation of exosome release from mammary epithelial and breast cancer cells-a new regulatory pathway. Eur J Cancer 2014;50:1025-34. [CrossRef]
36. Zhou W, Fong MY, Min Y, Somlo G, Liu L, Palomares MR, et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 2014;25:501-15. [CrossRef]
37. Suetsugu A, Honma K, Saji S, Moriwaki H, Ochiya T, Hoffman RM. Imaging exosome transfer from breast cancer cells to stroma at metastatic sites in orthotopic nude-mouse models. Adv Drug Deliv Rev 2013;65:383-90. [CrossRef]
38. Feng T, Zhang P, Sun Y, Wang Y, Tong J, Dai H, et al. High throughput sequencing identifies breast cancer-secreted exosomal LncRNAs initiating pulmonary pre-metastatic niche formation. Gene 2019;710:258-64. [CrossRef]
39. Huang H, Zheng X, Cai C, Yao Z, Lu S, Meng X, et al. Exosomes derived from breast cancer lung metastasis subpopulations promote tumor self-seeding. Biochem Biophys Res Commun 2018;503:242-8. [CrossRef]
40. Hoshino A, Costa-Silva B, Shen TL, Rodrigues G, Hashimoto A, Mark MT, et al. Tumour exosome integrins determine organotypic metastasis. Nature 2015;527:329-35. [CrossRef]
41. Holzer K, Ori A, Winkler J, Eiteneuer E, Breuhahn K, Beck M, et al. Nup155 is linked to the p53 pathway in HCC. Zeitschrift Für Gastroenterol 2014;52:3-20. [CrossRef]
42. Bose R, Kavuri SM, Searleman AC, Shen W, Shen D, Koboldt DC, et al. Activating HER2 mutations in HER2 gene amplification negative breast cancer. Cancer Discov 2013;3:224-37. [CrossRef]
43. Ciravolo V, Huber V, Ghedini GC, Venturelli E, Bianchi F, Campiglio M, et al. Potential role of HER2-overexpressing exosomes in countering trastuzumab-based therapy. J Cell Physiol 2012;227:658-67. [CrossRef]
44. Melo SA, Luecke LB, Kahlert C, Fernandez AF, Gammon ST, Kaye J, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015;523:177-82. [CrossRef]
45. Kibria G, Ramos EK, Lee KE, Bedoyan S, Huang S, Samaeekia R, et al. A rapid, automated surface protein profiling of single circulating exosomes in human blood. Sci Rep 2016;6:36502. [CrossRef]
46. Cazzoli R, Buttitta F, Di Nicola M, Malatesta S, Marchetti A, Rom WN, et al. microRNAs derived from circulating exosomes as noninvasive biomarkers for screening and diagnosing lung cancer. J Thorac Oncol 2013;8:1156-62. [CrossRef]
47. Ju SM, Jo YS, Jeon YM, Pae HO, Kang DG, Lee HS, et al. Phosphorylation of eIF2a suppresses cisplatin-induced p53 activation and apoptosis by attenuating oxidative stress via ATF4-mediated HO-1 expression in human renal proximal tubular cells. Int J Mol Med 2017;40:1957-64. [CrossRef]
48. Josson S, Gururajan M, Sung SY, Hu P, Shao C, Zhau HE, et al. Stromal fibroblast-derived miR-409 promotes epithelial-to-mesenchymal transition and prostate tumorigenesis. Oncogene 2015;34:2690-9. [CrossRef]
49. Lee JH, Kim JA, Kwon MH, Kang JY, Rhee WJ. In situ single-step detection of exosome microRNA using a molecular beacon. Biomaterials 2015;54:116-25. [CrossRef]
50. Hannafon BN, Trigoso YD, Calloway CL, Zhao YD, Lum DH, Welm AL, et al. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res 2016;18:90. [CrossRef]
51. Eichelser C, Stückrath I, Müller V, Milde-Langosch K, Wikman H, Pantel K, et al. Increased serum levels of circulating exosomal microRNA-373 in receptor-negative breast cancer patients. Oncotarget 2014;5:9650-63. [CrossRef]
52. Oguz G. Identification of Breast Cancer Stemness Genes as a Biomarker for Breast Cancer Progression, Doctoral Dissertation. Singapore:National University of Singapore;2019.
53. Guo J, Duan Z, Zhang C, Wang W, He H, Liu Y, et al. Mouse 4T1 breast cancer cell-derived exosomes induce proinflammatory cytokine production in macrophages via miR-183. J Immunol 2020;205:2916-25. [CrossRef]
54. Koldemir O, Özgür E, Gezer U. Accumulation of GAS5 in exosomes is a marker of apoptosis induction. Biomed Rep 2017;6:358-62. [CrossRef]
55. Shimoda M, Jackson HW, Khokha R. Tumor suppression by stromal TIMPs. Mol Cell Oncol 2016;3:e975082. [CrossRef]
56. Piombino C, Mastrolia I, Omarini C. The role of exosomes in breast cancer diagnosis. Biomedicines 2021;9:312. [CrossRef]
57. Galindo-Hernandez O, Villegas-Comonfort S, Candanedo F, González-Vázquez MC, Chavez-Ocaña S, Jimenez-Villanueva X, et al. Elevated concentration of microvesicles isolated from peripheral blood in breast cancer patients. Arch Med Res 2013;44:208-14. [CrossRef]
58. Rupp AK, Rupp C, Keller S, Brase JC, Ehehalt R, Fogel M, et al. Loss of EpCAM expression in breast cancer-derived serum exosomes:Role of proteolytic cleavage. Gynecol Oncol 2011;122:437-46. [CrossRef]
59. Moon PG, Lee JE, Cho YE, Lee SJ, Jung JH, Chae YS, et al. Identification of developmental endothelial locus-1 on circulating extracellular vesicles as a novel biomarker for early breast cancer detection. Clin Cancer Res 2016;22:1757-66. [CrossRef]
60. Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics 2017;7:789-804. [CrossRef]
61. Dong X, Bai X, Ni J, Zhang H, Duan W, Graham P, et al. Exosomes and breast cancer drug resistance. Cell Death Dis 2020;11:987. [CrossRef]
62. Ko J, Carpenter E, Issadore D. Detection and isolation of circulating exosomes and microvesicles for cancer monitoring and diagnostics using micro-/nano-based devices. Analyst 2016;141:450-60. [CrossRef]
63. Taylor DD, Gercel-Taylor C. Exosomes/microvesicles:Mediators of cancer-associated immunosuppressive microenvironments. Semin Immunopathol 2011;33:441-54. [CrossRef]
64. Luga V, Zhang L, Viloria-Petit AM, Ogunjimi AA, Inanlou MR, Chiu E, et al. Exosomes mediate stromal mobilization of autocrine wnt-pcp signaling in breast cancer cell migration. Cell 2012;151:1542-56. [CrossRef]
65. Zhu W, Huang L, Li Y, Zhang X, Gu J, Yan Y, et al. Exosomes derived from human bone marrow mesenchymal stem cells promote tumor growth in vivo. Cancer Lett 2012;315:28-37. [CrossRef]
66. Peinado H, Aleckovic M, Lavotshkin S, Matei I, Costa-Silva B, Moreno-Bueno G, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through met. Nat Med 2012;18:883-91. [CrossRef]
67. Zhao Z, Yang Y, Zeng Y, He M. A microfluidic ExoSearch chip for multiplexed exosome detection towards blood-based ovarian cancer diagnosis. Lab Chip 2016;16:489-96. [CrossRef]
68. Ohno S, Takanashi M, Sudo K, Ueda S, Ishikawa A, Matsuyama N, et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther 2013;21:185-91. [CrossRef]
69. Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science 2015;348:69-74. [CrossRef]
70. Marleau AM, Chen CS, Joyce JA, Tullis RH. Exosome removal as a therapeutic adjuvant in cancer. J Transl Med 2012;10:134. [CrossRef]
71. Gorczynski RM, Zhu F, Chen Z, Kos O, Khatri I. A comparison of serum miRNAs influencing metastatic growth of EMT6 vs 4THM tumor cells in wild-type and CD200R1KO mice. Breast Cancer Res Treat 2017;162:255-66. [CrossRef]
72. Rana S, Zöller M. Exosome target cell selection and the importance of exosomal tetraspanins:A hypothesis. Biochem Soc Trans 2011;39:559-62. [CrossRef]