
1. INTRODUCTION
The cultivated and wild pepper belongs to the Solanaceae family of the genus Capsicum. The bell or sweet peppers (Capsicum annuum L.) and the pungent or “bird’s eye pepper” (Capsicum frutescens L.) are the two widely grown species [1]. The world production of pepper (sweet and hot varieties) is estimated at 26,537 million tons, with China leading with an output of 7072 million tons representing 27% of the world production [2]. In Africa, Algeria is the lead producer (317,500 tons), followed by Tunisia with 280,000 tons [2]. Pepper is an important spice crop, highly cherished for its pungent flavor. Pungency comes from the capsaicinoids, alkaloid compounds (C18H22NO3) that abound only in the genus Capsicum [3]. Pepper fruit is enriched with vitamins A, C, and B2, potassium, phosphorus, and calcium, and in industries, pepper serves as an ingredient for many pharmaceuticals, for example., antioxidant and anticancer products; in food and cosmetics, the red pigment extract of ripe fruits serves as a natural coloring agent [1].
Pepper production in Sub-Saharan Africa is hindered by a number of biotic and abiotic factors. In Nigeria, a large amount of the crop comes from the semiarid area where drought and salinity are potential production constraints; thus, pepper is mostly grown under irrigation. Pepper is a salt-sensitive crop [4]. Salt may originate from several sources, such as seawater and salt strata. In the arid and semiarid regions, it originates from surface run-off and capillary movement. Poor source/quality of irrigation water may also constitute a source. Excess salt in the root zone inhibits water and nutrient absorption by the plant and may induce toxic effects [5,6]. Some saline soils contain sufficient salts that can suppress plant growth by upsetting osmotic potential and enzyme activities in plants [7-9].
The best management option for the production of crops under saline conditions has been the use of salt-tolerant varieties [5,10] and the use of the extracts of some plants [6,9]. Recently, however, researches have shown that arbuscular mycorrhizal fungi (AMF) are effective in alleviating the stress induced by salts on plants in saline soils [4,11-14]. The physiological and biochemical mechanisms of salt stress amelioration by AMF remain topical issues. Enhanced nutrient acquisition, especially immobile elements such as P, Cu, and Zn [4,13], has been attributed in part to the induced salt-tolerance by AMF-colonized plants. In recent studies, other potential mechanisms have been advocated, which include greater osmotic adjustment, reduced oxidative damage, selective toxic ion absorption like Na+, vacuolar compartmentalization of toxic ions, and increased production of antioxidative enzymes [7,14,15]. In spite of the substantial scientific evidence that AMF colonization alleviates salt stress, some researchers have explicitly stated that salt has negative effects on AMF [16,17]. Apart from salinity, root-knot nematodes (Meloidogyne spp.) are a serious threat to pepper production in the tropics [10,11,18].
Chemical control of plant-parasitic nematodes is quite efficient, but its associated drawbacks including environmental and health issues present a serious challenge. Recently, many authors have reported the effectiveness of AMF in the control of root-knot nematodes in several crops [19-22]. The various mechanisms attributed to nematode inhibition by AMF are still under debate [23,24]. However, induced systemic resistance/tolerance and histological, physiological, morphological, and biochemical changes associated with AMF inoculation have been implicated in nematode control and improved crop growth [25,26]. Galling of plant roots by root-knot nematodes under the saline condition could lead to growth and yield impairment if not checked [27,28]. There is a paucity of information on the salt stress ameliorating ability and root-knot disease control by AMF with both stress factors occurring simultaneously. The objective of this work was to evaluate the efficacy of AMF in the control of Meloidogyne incognita on sweet pepper exposed to salt stress.
2. MATERIALS AND METHODS
2.1. Experimental Site
Calabar is located in the tropical rainforest zone of Nigeria within latitude 5°00’ and 5°40’N and longitude 8°04’ and 8°62’E with an elevation of 70 m above sea level. The research was done in a screenhouse of the Faculty of Agriculture, University of Calabar, from January 7 to July 30, 2013. The environmental parameters of the screenhouse during the experimental period were as follows: average temperature range 28oC–31oC, relative humidity 69% to 81%, and sun hours 325 h/month to 371 h/month.
2.2. Source of Materials
The pepper seeds (Capsicum annuum cv. Tatase) used in the experiment were obtained from the crop germplasm collection of the Crop Science Department, University of Nigeria, Nsukka. The pepper cultivar is susceptible to M. incognita [10]. The starter culture of AMF species Glomus mosseae FR113 (Nicol. and Gerd.) Gerd. and Trappe, Glomus deserticola FL912 (Trappe, Boss, and Menge), and Gigaspora gigantea VA 105 (Nicol. and Gerd.) were procured from the Department of Agronomy, University of Ibadan, Oyo State, Nigeria. The accessions were collected from France, Florida, and Virginia and cultured by the International Institute of Tropical Agriculture, Ibadan, Nigeria. The salt used for the trial was sodium chloride (NaCl) and was bought from a chemical shop in Calabar.
2.3. Soil Sample Collection and Analysis
A composite surface soil (0–15 cm) was collected from the University of Calabar Teaching and Research Farm and was analyzed for its physicochemical properties according to the methods of Tel and Rao [29]. The soil from this farm was used to fill plastic pots which served as the growth medium for the pepper plants.
2.4. Multiplication of Starter Culture of AMF and M. incognita Inoculum
The AMF starter culture which comprises pieces of roots of maize (Zea mays L.) plants, glomerospores, spores, and soil was increased in heat-sterilized soil. The soil was sterilized by heating in an earthen pot to a temperature of 100°C and maintained for an hour, allowed to cool for 5 d, and then planted with maize, and Hoagland’s solution low in phosphorus was used with the irrigation water for 3 months. The spore densities for G. mosseae, G. deserticola, and Gi. gigantea were 51, 49, and 48 spores/10 g of soil, respectively, determined based on the method of Gerdemann and Nicolson [30]. The stock culture of M. incognita kept on Talinum fruticosum (L.) Juss. (waterleaf) in the vicinity of the screenhouse was multiplied in Celosia argentea L. (Cock’s comb) in a sterilized soil. It served as an inoculum source.
2.5. Preparation of Nematode Inoculum
Roots of C. argentea heavily galled by M. incognita were carefully removed from the planting pots, washed with water, and later cut into pieces (1–2 cm segments) for egg extraction using the method of Hussey and Barker [31]. The galled root segments were shaken thoroughly for 4 min in a solution of 0.5% NaOCl in a 500 ml conical flask with the mouth of the flask tightly covered. The eggs in the solution were sifted by placing a 200-mesh sieve over a 500-mesh sieve. The eggs were carefully collected in a beaker by washing with a wash bottle, and the inoculum density was determined by counting with a stereomicroscope. The average of three counts gave approximately 500 eggs in 1 ml of suspension.
2.6. Nursery of Pepper Seedlings and Inoculation with AMF
Inoculation of pepper plants with the respective AMF species was done at the nursery stage. Sandy soil and poultry manure were mixed at the ratio of 3:1 and then heat-sterilized. Two and a half kilograms of the heat-sterilized soil admixture was placed in plastic baskets to which the top 5–6 cm layer was placed with the AMF inoculum. The pepper seeds were surface-sterilized with a 0.5% NaOCl solution (household bleach) and rinsed 3 times with distilled water. The seeds were planted in each basket by drilling which was later thinned to 25 plants per basket after emergence. Baskets that had seedlings with no AMF inoculation represented the control. The plants were watered appropriately. This inoculation method follows the procedure of Oyekanmi et al. [32].
2.7. Inoculation of Pepper Seedlings with M. incognita and Irrigation with Saline Water
Five-week-old pepper seedlings (1.5–2 cm average leaf width and 3–5 cm average leaf length) were transplanted to the screenhouse in the evening and inoculated with the root-knot nematode (M. incognita). Seventy-two (72) plastic pots were used for the experiment. Holes were made at the bottom of each pot for water drainage. Pots were labeled appropriately for easy identification and treatment application. Each of the labeled pots was filled with 2.5 kg of sterilized soil. The pepper seedlings were transplanted and inoculated by making three holes around each seedling and pouring 10 ml of the inoculum suspension containing 5,000 eggs near the roots. Uninoculated seedlings served as the control. Salinity levels of 3.24 and 6.06 dS/m were obtained by dissolving 31.68 and 63.50 g of NaCl, respectively, in 18 L of tap water. The electrical conductivity of the saline irrigation water (ECw) was measured with an electrical conductivity meter. Irrigation with saline water (150 ml/plant) commenced 2 weeks after transplanting on a daily basis. The control plants were irrigated with tap water with electrical conductivity (ECw) of 0.16 dS/m.
2.8. Experimental Design
It was a 2 × 3 × 4 factorial experiment fitted into a completely randomized design (CRD) with three replications. The first factor was nematode inoculation, with uninoculated plants serving as control. This factor was combined with the three salt levels and the three AMF species’ inoculation plus nonmycorrhizal plants as control. Thus, there were 24 treatment combinations with three replications amounting to 72 pots.
2.9. Data Collection, Statistical Analysis, and Evaluation of Mycorrhizal Colonization
Data were collected on plant height (cm) and the number of leaves per plant at 7 w after transplanting. At 7 w after transplanting, each plant was uprooted, and roots were washed thoroughly with flowing water. The numbers of galls and egg masses per root system were counted. The galling index was scored on a 0–5 scale rating as used by Taylor and Sasser [33], where 0 = no galls, 1 = 1 or 2 galls, 2 = 3 to 10 galls, 3 = 11 to 30 galls, 4 = 31 to 100 galls, and 5 = more than 100 galls per root system. According to Sassar et al. [34], 0 = immune, 1 = highly resistant, 2 = resistant, 3 = moderately susceptible, 4 = susceptible, and 5 = highly susceptible. Egg masses were stained with phloxine B (0.15 g/l) for 15 min according to the procedure by Daykin and Hussey [35] for easy counting. The fresh root was separated from the shoot, and fresh root weight was obtained by weighing with an electronic balance, while the dry shoot weight was obtained after oven drying in an envelope at 70°C for 48 hr. Data obtained from the study were statistically analyzed using the analysis of variance (ANOVA) for a three-factor factorial experiment in CRD with GenStat 10th Release Version statistical software. Means were compared using Duncan’s new multiple range test at the 5% probability level. There was a need to transform data on egg mass counts using square root transformation (x + 0.50) before statistical analysis as there were zero counts. The section of the roots colonized by AMF was determined by a grid-line intersecting method [36] after clearing the roots with KOH [37], roots stained with 0.05% trypan blue-lactophenol [38,39].
3. RESULTS
3.1. Soil Properties
The results of the soil analysis showed that the soil used for the study was loamy sand, with a pH of 5.80 (moderately acidic), electrical conductivity of the extract (ECe) = 0.13 dS/m, low total N (0.1%), exchangeable cations, ECEC, medium available P (24.63 mg/kg), and very high base saturation.
3.2. Gall Index/Root System
The result showed that the effects of AMF, salinity, and nematodes and their interactions on gall index were significant [Table 1]. No gall was found on the roots of pepper plants not inoculated with M. incognita [Table 2]. The plants irrigated with tap water without salt and uninoculated with AMF significantly (P ≤ 0.05) had the highest gall index (GI) of 5.00 in the presence of M. incognita and were rated highly susceptible. In both AMF-inoculated and uninoculated plants, irrigation with saline water significantly reduced root galling. However, among the mycorrhizal plants, those inoculated with G. deserticola and irrigated with 6.06 dS/m salt water significantly had the least root gall index of 3.00 and were rated moderately susceptible.
 | Table 1: ANOVA for arbuscular mycorrhizal fungus (M), salinity (S), and Meloidogyne incognita (N) and their interactions for all the variables studied in sweet pepper. [Click here to view] |
 | Table 2: Effects of AMF and salinity on gall index per root system of pepper inoculated with or without Meloidogyne incognita. [Click here to view] |
3.3. Number of Egg Masses/Root System
The results showed that AMF, salinity, and nematodes and their interactions significantly (P ≤ 0.05) affected the number of egg masses produced per plant [Table 1]. The control plants without nematode inoculation had no egg mass on their roots [Table 3]. In both mycorrhizal and nonmycorrhizal plants, there was a significant (P ≤ 0.05) decrease in the number of egg masses produced by M. incognita with successive increases in the concentration of saline water. In general, inoculation of pepper plants with AMF significantly reduced egg mass production by M. incognita compared with the uninoculated plants. The lowest number of egg masses was observed in plants irrigated with 6.06 dS/m saline water and inoculated with G. deserticola in the presence of M. incognita infection. This was closely followed by G. mosseae-inoculated plants.
3.4. Fresh Root Weight
The result indicated that AMF, salinity, and nematodes and their interactions significantly (P ≤ 0.05) affected fresh root weight [Table 1]. With the exception of the highest level of salinity (6.06 dS/m), M. incognita infection significantly reduced the fresh root weight of pepper in both AMF-inoculated and uninoculated pepper plants compared with uninfected plants [Table 4]. However, at that salt level, nematode infection significantly reduced fresh root weight with G. deserticola inoculation compared with noninfected plants. Also, in the presence or absence of nematodes, successive increases in salinity level led to a significant decrease in fresh root weight of both AMF-inoculated and uninoculated plants. Relative to the nonmycorrhizal plants, plants inoculated with AMF at all salinity levels and with M. incognita inoculation significantly had higher fresh root weight except for those inoculated with Gi. gigantea which had similar results to the control without AMF at 3.24 d/Sm. Among the nematode-inoculated or uninoculated plants under salt stress, plants inoculated with G. deserticola produced significantly higher fresh root weight.
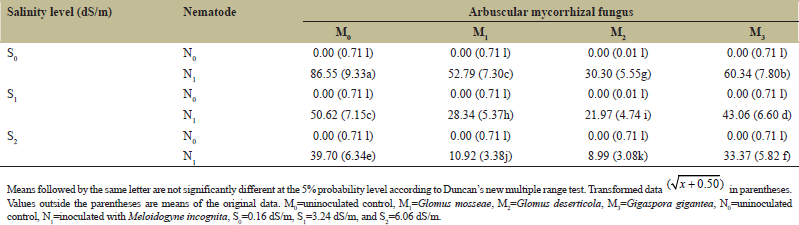 | Table 3: Effects of AMF and salinity on the number of egg masses per root system of pepper inoculated with or without Meloidogyne incognita. [Click here to view] |
 | Table 4: Effects of AMF and salinity on the fresh root weight (g plant-1) of pepper inoculated with or without Meloidogyne incognita. [Click here to view] |
3.5. Percentage Root Colonization
The effects of AMF, salinity, and nematodes and their interactions were significant (P ≤ 0.05) on percentage root colonization by AMF [Table 1]. No root colonization was observed in plants not inoculated with AMF [Table 5]. At salinity level of 0.16 dS/m, root-knot nematode infection significantly reduced root colonization by G. mosseae and G. gigantea but not that of G. deserticola. However, when pepper plants were irrigated with saline water nematode infection significantly reduced root colonization by G. deserticola. At all salinity levels in the presence or absence of nematodes, G. deserticola significantly had the highest percentage root colonization with the exception of G. mosseae-inoculated plants at 6.06 dS/m in the presence of M. incognita that did not differ from G. deserticola-inoculated plants.
3.6. Plant Height
The effects of AMF, salinity, and nematodes and their interactions on plant height were significant [Table 1]. Irrespective of the salinity level and AMF species, M. incognita inoculation significantly reduced the height of pepper plants compared with the nematode-free plants [Table 6]. In both mycorrhizal and nonmycorrhizal plants, increase in salinity level in the presence or absence of M. incognita resulted in a significant decrease in plant height. In nematode-inoculated plants, at all salt levels, plants inoculated with AMF were significantly taller than their uninoculated counterparts. However, in all cases, G. deserticola-inoculated plants were significantly taller than others followed by G. mosseae-inoculated plants.
 | Table 5: Effects of salinity and Meloidogyne incognita on percentage (%) root colonization of pepper plants by different AMF species. [Click here to view] |
 | Table 6: Effects of AMF and salinity on plant height (cm) of pepper inoculated with or without Meloidogyne incognita. [Click here to view] |
3.7. Number of Leaves
The results showed that only the individual effects of AMF, salinity, and nematodes and the interaction between AMF and nematodes were significant on the number of leaves produced by pepper plants [Table 1]. AMF inoculation significantly (P ≤ 0.05) increased the number of leaves relative to the nonmycorrhizal plants [Table 7]. However, G. deserticola-inoculated plants produced a significantly higher number of leaves ahead of G. mosseae-inoculated plants. Successive increases in salt level significantly reduced the number of leaves. Furthermore, in both mycorrhizal and nonmycorrhizal plants, nematode inoculation significantly reduced the number of leaves. However, among AMF-inoculated plants in the presence of M. incognita, G. deserticola-inoculated plants produced a significantly higher number of leaves.
 | Table 7: Effects of AMF and salinity on the number of leaves per plant (NL) of pepper inoculated with and without Meloidogyne incognita. [Click here to view] |
3.8. Dry Shoot Weight
The effects of AMF, salinity, and nematodes and their interactions were significant on the dry shoot weight of pepper [Table 1]. In both mycorrhizal and nonmycorrhizal plants, nematode inoculation significantly reduced dry matter accumulation in the shoot at all salinity levels with the exception of nonmycorrhizal plants at 3.24 dS/m and Gi. gigantea-inoculated plants at 6.06 dS/m [Table 8]. In some cases, there was a significant reduction in shoot dry matter as salinity level was increased from 0.16 dS/m (control) to 6.06 dS/m in both mycorrhizal and nonmycorrhizal plants in the presence or absence of M. incognita. In all cases, G. deserticola-inoculated plants in the presence or absence of nematodes at all salinity levels accumulated the highest dry matter in shoot compared with the other AMF species.
4. DISCUSSION
Evidence from this study showed that salinity significantly reduced plant height, fresh root weight, shoot dry weight, and the number of leaves in pepper. Salinity reduces the growth and development of plants [4,6,12]. These findings agree with Abdel Latef and Chaoxing [14], who reported that salinity stress significantly reduced assimilate partitioning to root, stem, leaf, and leaf area of tomato compared with the control plants. They attributed these to direct effects of ion toxicity or indirect effects of saline ions resulting in soil/plant osmotic imbalance. Salinity also affected nematode activities. It was observed that where there was nematode inoculation, a higher level of salt-reduced nematode activities. Root galling was reduced at the higher salt level. A similar observation was made by Hamdy et al. [28]. This could be due to the reduction of growth and development of the host plant which leads to the failure of the host tissue to keep pace with the nutritional demands of the nematodes [36]. This finding corroborates the report of Edongali and Ferris [27] and Hamdy et al. [28] that increase in salinity decreased M. incognita reproduction and root galling in susceptible varieties of tomato and okra, respectively.
 | Table 8: Effects of AMF and salinity on dry shoot weight (g/plant) of pepper inoculated with or without Meloidogyne incognita. [Click here to view] |
High levels of salinity suppressed root colonization by arbuscular mycorrhiza fungi. This is in agreement with the reports by several researchers [16,17,37]. They reported the negative effects of salinity, stating that the rate of spore germination, hyphal colonization of root, and hyphal growth of the fungus were impeded. Pepper seedlings with AMF significantly had reduced effects of salts. This was obvious with the fact that the pepper plants inoculated with AMF had higher fresh root weight and shoot dry weight and were taller. This conforms to the reports of several researchers who reported enhanced growth attributes and yield by AMF-inoculated plants under saline stress compared to the nonmycorrhizal plants. They stated that plant growth and biomass suffered a setback under salt stress. This may likely be due to nutrient deficiency and waste of energy to alleviate the toxic effects of salt [12,22,40].
Inoculation of pepper seedlings with AMF reduced galling and egg mass production. This agrees with Borowicz’s [41] findings where he listed various results of many workers confirming the reduction in the reproduction of sedentary nematodes due to AMF inoculation. The decrease in the fecundity was attributed to physiological changes and/or physical or chemical barriers leading to the unattractiveness of the roots for easy penetration by plant-parasitic nematodes. The inhibition in root galls observed on pepper seedlings could have occurred as a result of competition between the nematode and the symbiont for infection sites, though other factors such as higher concentration of lignin and phenols could be involved [19,24]. Zhang et al. [19] recorded fewer galls and egg masses in cucumber plants inoculated with AMF than the uninoculated plants. This was attributed to the failure of the nematode to penetrate the root and initiate the formation of giant cells as well as impairment of the nematode’s development [23,42,43].
However, among the AMF species, G. deserticola was the most effective in reducing root-knot nematode infection and alleviation of salt stress in pepper plants. There are various reports on the variation in the efficacy of different AMF in nematode control and salinity amelioration [21,44]. Mohammed and Mittra [44] observed that an isolate of a stress-adapted AMF species, G. deserticola, was the most effective in alleviating salinity and heavy metal stresses as well as enhancement of Solanum melongena and Sorghum sudanense growth and dry matter yield compared to other AMF species. This is in line with the result of this trial. In general, this trial has illustrated the possibility of managing the abiotic stress factor induced by salinity and the biotic factor caused by M. incognita on pepper production with proper selection and inoculation of the symbiont, AMF. Under saline soils, appropriate and sufficient AMF propagules may be lacking, thus the need for inoculation of nursery seedlings with efficient AMF species.
5. CONCLUSION
AMF inoculation significantly reduced root galling and egg masses by M. incognita and also enhanced growth and dry matter production of sweet pepper plants in a simulated salinity condition. This study revealed that the most efficient AMF species in salinity amelioration and reduction in nematode infectivity was G. deserticola followed by G. mosseae. In conclusion, G. deserticola and G. mosseae could be effectively utilized as eco-friendly management bioagents in pepper production where salinity and M. incognita are constraints.
6. ACKNOWLEDGMENTS
The authors are grateful to Mrs. Giwa of the University of Ibadan for providing the starter culture of AMF and the University of Calabar for providing research facilities for the work to be carried out.
7. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All data generated and analyzed are included within this research article.
12. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Seleshi D. Evaluation of Elite Hot Pepper Varieties (Capsicum species) for Growth, Dry Pod Yield and Quality under Jimma Condition, Southwest Ethiopia. M.Sc Thesis (Horticulture), Jimma University; 2011.
2. Food and Agriculture Organization. Food and Agricultural Commodities Production. Rome, Italy: Food and Agriculture Organization of the United Nations Corporate Document Repository; 2010.
3. Bosland PW, Votava EJ. Peppers, Vegetables and Spices Capsicum. New York: CABI Publishing; 2000. p. 198.
4. Kaya C, Ashraf M, Sonmez O, Aydemirs S, Tuma LA, Cullu AM. The influence of arbuscular mycorrhizal colonization on key growth parameters and fruit yield of pepper plants grown at high salinity. Sci Hortic 2009;121:1-6. CrossRef
5. Chartzoulakis K, Klapaki G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci Hortic 2000;86:247-60. CrossRef
6. Abdel Latef AA, Abu Alhmad MF, Kordrostami M, Abo-Baker AE, Zakir A. Inoculation with Azospirillum lipoferum or Azotobacter chroococcum reinforces maize growth by improving physiological activities under saline conditions. J Plant Growth Regul 2020;39:1293-306. CrossRef
7. He Z, He C, Zhang Z, Zou Z, Wang H. Changes of antioxidative enzymes and cell membrane osmosis in tomatoes colonized by arbuscular mychorrhiza under NaCl stress. Colloid surfaces B. Biointerfaces 2007;59:128-33. CrossRef
8. Shiyab SM, Shantawi MA, Shibli RA. Growth, nutrient acquisition, and physiological responses of hydroponic grown tomato to sodium chloride salt induced stress. J Plant Nutr 2013;36:665-76. CrossRef
9. Osman MS, Badawy AA, Osman AI, Abdel Latef AA. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J Plant Growth Regul 2020;40:1245-56. CrossRef
10. Udo IA, Uguru MI, Ogbuji RO. Pathogenicity of Meloidogyne incognita Race 1 on Nigerian pepper (Capsicum spp) lines. Glob J Agric Sci 2005;4:23-7. CrossRef
11. Bernardo BF, Garita SA, Arango MC, Ripodas JI, Saparrat MC, Ruscitti MF. Arbuscular mycorrhizal fungi against false root-knot nematode activity in Capsicum annum; physiological responses in plants. Biocontrol Sci Technol 2020;31:119-31. CrossRef
12. Giri B, Mukerji K. Mycorrhizal inoculant alleviates salt stress in Sesbania aegypticca and Sesbania granndiflora under field conditions. Evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004;14:307-12. CrossRef
13. Al-Karaki GN. Nursery inoculation of tomato with arbuscular myccorhizhal fungi and performance under irrigation with saline water. Sci Hortic 2006;109:1-7. CrossRef
14. Abdel Latef AH, Chaoxing H. Effect of arbuscular mycorrhizal fungus on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci Hortic 2011;127:228-33. CrossRef
15. Ruiz-Lozano JM. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress: New perspectives for molecular studies. Mycorrhiza 2003;13:309-17. CrossRef
16. Hirrel MC. The effect of sodium and chloride salts on the germination of Gigaspora margarita. Mycology 1981;73:610-7. CrossRef
17. Juniper S, Abbott L. Vesicular arbuscular mycorrhiza and soil salinity. Mycorrhiza 2006;4:45-57. CrossRef
18. Sowley EN, Kankam F, Adomako J. Management root-knot nematode (Meloidogyne spp) on sweet pepper (Capsicum annum L.) with Moringa (Moringa oleifera Lam) leaf powder. Arch Phytopathol Plant Prot 2014;47:1531-8. CrossRef
19. Zhang L, Zhang J, Christie P, Li X. Effect of inoculation with the arbuscular mycorrhizal fungus Glomus intraradices on root knot nematode Meloidogyne incognita in Cucumber. J Plant Nutr 2009;32:967-79. CrossRef
20. Odeyemi IS, Afolami SO, Sosanya OS. Effects of Glomus mosseae (Arbuscular mycorrhizal fungus) on host-parasite relationship of Meloidogyne incognita on four improved cowpea varieties. J Plant Prot Res 2010;50:25-32. CrossRef
21. Udo IA, Uguru MI, Ogbuji RO. Comparative efficacy of arbuscular mycorrhizal fungi in combination with bioformulated Paecilomyces lilacinus against Meloidogyne incognita on tomato in two Ultisols of south eastern Nigeria. Biocontrol Sci Technol 2013;23:1083-97. CrossRef
22. Sharma IP, Sharma AK. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhizal and PGPR against root-knot nematodes. Symbiosis 2017;71:175-82. CrossRef
23. Gera Hol. WH, Cook R. An overview of arbuscular mycorrhizal fungi nematode interactions. Basic Appl Ecol 2005;6:489-503. CrossRef
24. Campos MA. Bioprotection by arbuscular mycorrhizal fungi in plant infected with Meloidogyne nematodes: A sustainable alternative. Crop Prot 2020;135:105203. CrossRef
25. Vos C, Schouteden N, van Tuinen D, Chatagnier O, Elsen A, de Waele D, et al. Mycorrhiza induced resistance against the root-knot nematode Meloidogyne incognita involves priming of defense gene response in tomato. Soil Biol Biochem 2013;60:45-54. CrossRef
26. Vos, CM, Tesfashun AN, Panis B, de Waele D, Elsen A. Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl Soil Ecol 2012;61:1-6. CrossRef
27. Edongali E, Ferris H. Varietal response of tomato to the interaction of salinity and Meloidogyne incognita infection. J Nematol 1982;14:57-62.
28. Hamdy FA, Gad SB, Abo-El-Heer EA, Aldosari A. Influence of three rates of salinity on okra plants (Abelmoschus esculentus L.) infected with root-knot nematode in vivo. Arch Phytopathol Plant Prot 2019;52:277-87. CrossRef
29. Tel DA, Rao P. Automated and Semi-automated Methods for Soil Plant Analysis. Manual Series No.7 IITA, Ibadan, Nigeria; 1982.
30. Gerdemann JW, Nicolson TH. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans Br Mycol Soc 1963;46:235-44. CrossRef
31. Hussey RS, Barker KR. Comparison of methods of collecting inocula of Meloidogyne spp including a new technique. Plant Dis Rep 1973;57:1025-8.
32. Oyekanmi EO, Coyne DL, Fagade OE, Osonubi O. Improving root-knot nematode management of two soybean genotypes through the application of Bradyrhizobium japonicum, Trichoderma pseudokoningii and Glomus mosseae in full factorial combinations. Crop Prot 2007;26:1006-12. CrossRef
33. Taylor AL, Sasser JN. Biology, Identification and Control of Root-knot Nematodes (Meloidogyne Species). Raleigh, NC: Department of Plant Pathology, North Carolina State University and US Agency for International Development; 1978. p. 111.
34. Sassar JN, Carter CC, Hartman KM. Standardization of Host Suitability Studies and Reporting of Resistance to Root-knot Nematode. Raleigh, NC, USA, A Cooperative Publication of the Department of Plant Pathology, North Carolina State University and the United States Agency for International Development; 1984.
35. Daykin ME, Hussey RS. Staining and histopathology techniques. In: Barker KR, Carter CC, Sasser JN editors. An Advanced Treatise on Meloidogyne. Vol. 2. Raleigh: North Carolina University Graphics; 1985. p. 39-48.
36. Mote UN. Effect of chlorogenic acid on hatching and infectivity of root-knot nematode on tomato. Int Nematol Netw Newslett 1988;5:5-6.
37. Jahromi F, Aroca R, Porcel R, Ruiz-lozano J. Influence of salinity on the in-vitro development of Glomus intradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plant. Microb Ecol 2008;15:45-53. CrossRef
38. Giovanneti M, Mosse B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infections in roots. N Phytol 1980;84:489-500. CrossRef
39. Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 1970;55:158-61. CrossRef
40. Zuccarini P, Okurowska P. Effects of mycorrhizal colonization and fertilization on growth and photosynthesis of sweet basil under salt stress. J Plant Nutr 2008;31:497-513. CrossRef
41. Borowicz VA. Do arbuscular mycorrhiza fungi alter plant pathogens relations? Ecology 2001;82:3057-68. CrossRef
42. Flor-Peregrin E, Azcon R, Martos V, Vedejo-Lucas S, Talavera M. Effects of dual inoculation of mycorrhiza and endophytic, rhizospheric or parasitic bacteria on root-knot disease of tomato. Biocontrol Sci Technol 2014;24:1122-36. CrossRef
43. Vos C, Geerinckx KK, Mkandawire R, Panis BS, de Waele D, Elsen A. Arbuscular mycorrhizal fungi affect both penetration and further life development of root -knot nematodes in tomato. Mycorrhiza 2012;22:157-63. CrossRef
44. Mohammed A, Mittra B. Effects of inoculation with stress-adapted arbuscular mycorrhizal fungus Glomus deserticola on growth of Solanum melogena and Sorghum sudanes Staph. Seedling under salinity and heavy metal stress conditions. Arch Agron Soil Sci 2013;59:173-83. CrossRef