
1. INTRODUCTION
Allelopathy is the production of specific biomolecules by plant species (mostly secondary metabolites) that can induce suffering or benefit other plant species around it [1]. Allelopathy is the active or passive release of chemical substances by one plant into the environment by root exudation, leaching, volatilization, and decomposition. Allelochemicals are secondary metabolites and phytochemicals that correspond to the following classes: flavonoid, terpenoid, phenolic compound, organic cyanide, glucosinolate, saponin, alkaloid, and long chain fatty acid structures [2]. These are released as a result of enzymatic processes or as a result of environmental stress. Allelochemicals’ structure and concentration vary depending on biological and non-biological inputs, hence their targets and functions vary [3]. Green plants produce a wide range of compounds that are not involved in the primary metabolism of plants and hence are called secondary products or metabolites. The biosynthetic pathways of the secondary compounds as off-shoots of the primary metabolism elaborated these pathways of allelochemical(s) origin [4,5]. Competition is one of the most common ways of interaction between cultivated crops and their surrounding plants [6,7].
The plant inhibits the growth of other plants growing around it, a phenomenon called allelopathic effect. Allelopathic characteristics can be found in all parts of a plant, from the foliage and flowers to the roots, bark, soil, and mulch. Almost all allelopathic plants, especially during fall, store their protective chemicals in their leaves. These toxins can impact surrounding plants as the leaves fall to the ground and decompose. The allelopathic effect on plant growth and development depends on the compounds involved, their concentration and stability in the given environment, and plant response to the compounds or their microbial byproduct(s). The availability of biologically active concentrations of allelopathic compounds in the soil environment depends on the rates of input, absorption, and adsorption by seeds and roots, fixation in soil organic matter, leaching and microbial degradation, and persistence. In arid environments, the retention of inhibition is greater. Hence, they purist in the environment for a longer time. Allelopathic crops can help reduce noxious weeds and plant pathogens, enhance soil quality, and increase crop production when used as a cover crop, mulch, smother crop, green manure, or cultivated in rotational sequences. As a result, allelochemicals derived from allelopathic crops may be helpful in the creation of biological herbicides and pesticides [8]. Allelopathy has an alternative for developing eco-friendly agricultural practices, enhancing crop productivity, and maintaining ecosystem stability [9].
The tree is the dominant component in the agroforestry system. It impacts the related crops plants through leaf fall, exudates, and compounds produced at various stages of leaf litter decomposition. Inhibitory chemicals can be found in various plants and organs, including leaves, flowers, fruits, and buds. In agroforestry, where many crops are grown below the canopy of trees, allelopathic interactions are implicit. Phytochemicals present in tropical trees allelopathically retard the growth of associated crop species [10]. Coconut (Cocos nucifera L.), cashew (Anacardium occidentale L.), jackfruit (Artocarpus heterophyllus Lam.), mango (Mangifera indica L.), tamarind (Tamarindus indica L.), and teak (Tectona grandis L.f.) are examples of multipurpose trees that are often planted in agroforestry systems.
The allelochemical activity varies depending on the research methodology and operational processes [11]. The allelochemical released into the environment is usually not a single substance, and the amount of allelochemicals released varies depending on the circumstances. When Albuquerque et al. [12] investigated the allelopathic potential of plants, the type and amount of allelochemicals released by them were taken into account. In some instances, one allelochemical may not exhibit allelopathic action as a single component, but it may increase allelopathy when combined with additional allelochemicals.
Annona muricata is a coveted tropical tree and a wealth of phytochemical investigations has been conducted for this fruit plant. Annona muricata is a good source of bioactive compounds, such as polyphenols, acetogenins, and alkaloids [13,14]. In the A. muricata plant, polyphenols and acetogenins have been studied extensively. Annona muricata L., also known as soursop, graviola, guanabana, paw-paw, and sirsak, belongs to the Annonaceae family, which has approximately 130 genera and 2,300 species. Annona muricata has been shown to have a wide range of biological functions, in addition to being an essential food source and an indigenous medicinal plant. The anticancer, antiparasitic, and insecticidal effects of this plant have been determined to be the most promising in previous investigations. The fruit contains many nutrients such as vitamin C, calcium, dietary fiber, iron, protein, cholesterol, sodium sugars, unsaturated fats, etc. The fruit improves the immune system and avoids infections in the body.
The production of agricultural crops in Tamil Nadu is still on a subsistence scale mainly due to traditional farming methods which may not be sufficient to cater to the needs of the growing population. In order to boost the present production status on sustainable basis, commercial cultivation with maintaining sustainability in agricultural production is a great challenge. One of the options is to promote and popularize agroforestry through scientific intervention using crop combinations from the indigenous plant resources. However, for selecting suitable crop combinations in agroforestry systems for better productivity, the compatibility among the woody and non-woody components has to be checked thoroughly. The present study, therefore, aims to analyze the inhibitory and stimulatory effects of aqueous extracts of traditional medicinal perennial species of A. muricata on seed germination, morphological, physiological, and biochemical activities of Vigna radiata and Eleusine coracana.
2. MATERIALS AND METHODS
The experiment was carried out in the Botanical garden of Rani Anna Government College for women, Tirunelveli, India. The study site is located at 8.7139°N latitude 77.7567°E longitude. Fresh mature leaves of A. muricata plant were collected, washed separately with several changes of sterile distilled water, and air dried for 7 days. They were blended with a sterile blender and sieved with a fine sterile cheese clothes to obtain fine powder. Cold water extract of the blended leaves of A. muricata was prepared separately by adding 10, 20, 30, 40, and 50 g of leaves of the powdered test plants separately to the conical flask containing 1,000 ml of sterile distilled water and allowing them to settle for 24 hours. The resulting solutions were filtered through sterile cheese cloth, which then gave the plant extract concentrations of 1%, 2%, 3%, 4%, and 5%, respectively. The plant extracts were put in sterile McCartney bottles and labeled accordingly, while the research lasted. All preparations were carried out in replicates of three each. Distilled water was used as control.
The viable seeds of V. radiata and E. coracana were sown in mud earthen pots measuring 10 cm in diameter and 10 cm in depth, packed with garden soil. Freshly prepared aqueous leaf concentrates of 1%, 2%, 3%, 4%, and 5% were sprayed on the surface of V. radiata and E. coracana seedlings to uniformly cover the entire seedling surface. The control pots were sprayed with distilled water. They repeated each procedure three times. After 10 days of seed sowing, germination was recorded. 15 days after sowing, the seedlings were rooted out and washed with tap water to remove any soil left on the roots. For biophysical analysis, the root length and shoot length were measured in centimeter. Biochemical parameters such as chlorophyll a and b (using Arnon’s method), carotenoid (using Krik and Allen’s method), carbohydrate (using Anthrone’s method), protein (using Lowery et al.’s method) [15], amino acid (using Moore and Stein’s method), and catalase (using Machly and Chance’s method) were examined in the seedlings.
2.1. Statistical Analysis
All the experiments were carried out in triplicates and carried out separately thrice with consistent findings. Data are defined as mean of replicates ± SD. Differences between treatments were then tested using one-way analysis of variance for each parameter under review, followed by Duncan’s test (p ≤ 0.05).
 | Figure 1: Allelopathic influence of A. muricata on the germination percentage of V. radiata and E. coracana seedlings (10 days after sown). [Click here to view] |
3. RESULTS AND DISCUSSION
Significant effects of the treatment on germination percentage were recorded with the interaction between A. muricata and annual crops such as V. radiata and E. coracana (Fig. 1). The germination percentage of V. radiata in 1% aqueous leaf extract of A. muricata was stimulated (3.16%) and inhibited (1.04%) in E. coracana over control. Reduction in germination percentage was noted with application of A. muricata leaf extracts of 2%, 3%, 4%, and 5%, respectively, on test crops compared to control. The stimulatory effect was recorded at 1% extract treatment on V. radiata only. The stimulatory effect of germination percentage has been attributed to test crops due to the allelochemicals of leaf extracts of tree species. Venkateshwarlu et al. [16] recorded 90% inhibition in seed germination of okra when treated with leaf extract of M. indica. Both stimulatory and inhibitory effects were noted by Sahoo et al. [17] on seed germination of annual crops, with the aqueous leaf leachate of M. indica. Inhibition of seed germination by allelochemicals has been attributed to suppressing the mitotic activities of young cells [18]. Inhibition of germination occurred due to the presence of chemicals in the leaf extract of A. muricata such as muricoreacina, murihexocina, kaempferol, quercetin 3-O-glucoside, quercetin 3-oneohesperidoside, quercetin 3-O-robinoside, quercetin-O-rutinoside, quercetin 3-O-a-rhamnosyl, robinetin, tangeretin, and taxifolin. Shahena et al. [19] reported that the aqueous extract of Wedelia significantly inhibited the growth of pulse seedlings with a more pronounced effect at higher concentrations. This supports our present findings.
When the test plants were treated with different concentrations of aqueous leaf extract of A. muricata, they showed reduction in the growth of the seedlings over control (Fig. 2). Only 1% extract treatment in V. radiata showed a stimulatory effect of 2.07% root length over control and 2.97% shoot length over control, respectively. The root lengths were more sensitive to allelochemicals than the shoot length, thereby ultimately affecting the plant growth [20]. The leaf extracts inhibited the seedling growth at different inhibition values. The inhibitory activity was test species, plant parts extract-dependent. In few cases, the growth stimulatory activity was also observed. Many researchers, around the world reported this type of growth inhibition by the aqueous extract of allelopathic plants [21,22].
The tannin from A. muricata L. leaves also inhibited the seedling growth of V. radiata and E. coracana. The inhibition effect is concentration-dependent. This means that the inhibition in the growth of the seedling took effect with increasing concentration. In the control, the root length and shoot length after 15 days of seed sowing of V. radiata was 2.41 and 11.29 cm and of E. coracana was 1.86 and 10.20 cm, respectively. Negative values were obtained in the treatments indicating that the seedling ceased to grow, which may have led to the impairment in the seedling’s metabolic activities, resulting in a decrease in their root and shoot length [23]. Cell division also stopped because no shoots grew in all the treatment. In the study of Arowosgabe et al. [24], similar concentrations of 2, 4, 6, 8, and 10 mg/ml were used in observing the allelopathic effect of Aloe ferox mill root extract on tomato. Morikawa et al. [25], in their study, reported the growth inhibitory activity of A. muricata.
Inhibition in the seedling growth of Echinochloa crus-galli was observed using the same concentrations. Phenolic compounds, like coumarin, had inhibitory effects on seedling growth and seed germination on both test seedlings. The chemicals involved in allelopathic interactions are present in virtually all plant parts including leaves, stems, fruits, roots, rhizomes, buds, and seeds [26,27].
The results (Figs. 3 and 4) showed that the pigment contents of the test crops are highly influenced by A. muricata leaf extracts. In V. radiata test seedlings recorded, 1% extract treatment pigments were stimulated (Fig. 3). At 1% extract, stimulations of chlorophyll a (5.56%), chlorophyll b (3.52%), and carotenoid (2.12%), respectively, were compared to control. However, in E. coracana (Fig. 4), there was no stimulation noted at 1% extract treatment. Between the two crops, more inhibition was observed in E. coracana. The inhibition was concentration-dependent. At 5% extract treatment, more inhibition was recorded in both crops. The inhibition of pigments in V. radiate, chlorophyll a (76.39%), chlorophyll b (68.31%), and carotenoid (51.85%), and in E. coracana, chlorophyll a (79.57%), chlorophyll b (70.51%), and carotenoid (74.80%), was more than that in the control. The allelochemicals of A. murricata, such as muricatacin, isoquinolines, anonaine, anoniin, coreximine, lipids, gentisic acid, lignoceric acid, stearic acid, genistein, glycitein, homoorientin, isoferulic acid, kaempferol, quercetin 3-O-glucoside, quercetin 3-oneohesperidoside, quercetin 3-O-robinoside, quercetin-O-rutinoside, quercetin 3-O-a-rhamnosyl, robinetin, tangeretin, taxifolin, vitexin, caffeic acid, hydroxycinnamic acid, gentisic acid, hydroquinone, gallic acid, tannin, flavonoids, phenolic compounds, etc., might inhibit the pigments of test seedlings. Allelochemicals have several effects on plant photosynthesis, including inhibiting or damaging the synthesis machinery and hastening the breakdown of photosynthetic pigments (Table 1). Photosynthetic pigment contents are reduced, which blocks the energy and electron transfer, reduces adenosine triphosphate (ATP) synthesis enzyme activity, inhibits ATP synthesis, and affects stomatal conductance and transpiration, all of which inhibit the photosynthetic process. The allelochemicals influence photosynthesis primarily via altering PS II function [28–30]. Poonpaiboonpipat et al. [31] found that a high concentration of essential oil from lemongrass (Cymbopogon citratus) leaves reduced the chlorophyll a and b and carotenoid content of barnyard grass and affected alpha-amylase activity in seeds, implying that essential oils interfere with photosynthetic metabolism.
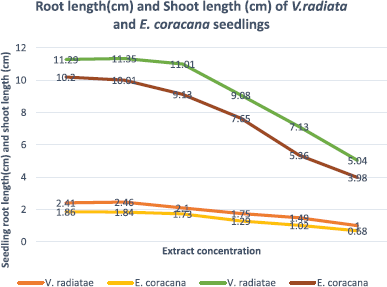 | Figure 2: Allelopathic influence of A. muricata on root length (cm) and shoot length (cm) of V. radiata and E. coracana seedlings (15 DAS). [Click here to view] |
 | Figure 3: Allelopathic influence of A. muricata on Chl a, Chl b, and carotenoid (mg/g fr. wt.) contents of V. radiata seedlings (15 DAS). [Click here to view] |
 | Figure 4: Allelopathic influence of A. muricata on Chl a, Chl b, and carotenoid (mg/g fr. wt.) contents of E. coracana seedlings (15 DAS). [Click here to view] |
A decrease in chlorophyll and carotenoid content in response to A. muricata allelochemicals indicates that it has a negative impact on photosynthetic efficiency. The observed effects are similar to the effects of essential oils, such as Eucalyptus citriodora and Artemisia scoparia, which reduce chlorophyll content and thus affect photosynthetic activity [32–34]. Scott [35], in his study, found that chlorophyll a, b, and carotenoid are key pigments of chloroplasts of plants that capture light energy. However, it is unclear whether the drop in chlorophyll concentration is related to a decreased de novo production or increased degradation. Nevertheless, the results showed a decline in carotenoid content. The chlorophyll content of V. radiata and E. coracana was reduced by the aqueous extract of A. muricata. The presence of allelochemicals in A. muricata was responsible for the decreased chlorophyll concentration observed in this study. Frabboni et al. [36] and Ojija et al. [37] have reported similar findings. Chlorophyll photooxidation takes place due to a lack of carotenoid [38,39]. Reduced seed germination in V. radiata and E. coracana might be caused by the allelopathic stress of different extract concentrations resulting from different abnormalities in metabolic activities and cell division abnormalities due to the effect of A. muricata (Figs. 5 and 6).
The biochemicals such as protein, amino acid, carbohydrate, peroxidase, and catalase of V. radiata and E. coracana seedlings were studied. In V. radiata (Table 2), amino acid (89.90%) and carbohydrate (71.09%) were reduced highly, followed by catalase (65.83%), protein (68.96%), and peroxidase (54.54%) at 50% concentration treatment of A. muricata, whereas lower concentration (1%) extract treatment stimulation of biochemicals as noticed. The allelochemicals of A. murricata, such as lignoceric acid, stearic acid, genistein, glycitein, homoorientin, isoferulic acid, kaempferol, quercetin 3-O-glucoside, quercetin 3-oneohesperidoside, quercetin 3-O-robinoside, quercetin-O-rutinoside, quercetin 3-O-a-rhamnosyl, robinetin, etc., inhibited the synthesis of biochemicals of test seedlings. In E. coracana (Table 3) seedlings, at all extract treatments only inhibition was noted. For the lower concentration (1%) extract treatment, very less amount of inhibition was observed. Protein (0.96%), amino acid (4.59%), carbohydrate (2.51%), peroxidase (0.74%), and catalase (1.85%) were recorded over control. The inhibition of the test seedlings was different in the same extract treatment. From this experiment, it was observed that the inhibition or stimulation of seedling growth, germination, and biochemicals were different in different species by the influence of allelochemicals of A. muricata. Allelochemicals affect the production, functions, contents, and activities of various enzymes in diverse ways. Peroxidase and catalase can be suppressed by taxifolin, vitexin, caffeic acid, hydroxycinnamic acid, gentisic acid, hydroquinone, gallic acid, tannin, flavonoids, and phenolic compounds, which can also reduce the synthesis of amylase and acid-phosphatase in the endosperm. Sofi et al. [40] have thoroughly documented the link between root length and plant failure to compete and search for water and minerals from the soil. Their works support the present findings.
 | Table 1: Structure of some of the allelochemicals produced by A. muricata. [Click here to view] |
 | Figure 5: Allelopathic effect of A. muricata on seedling growth of V. radiata. [Click here to view] |
 | Figure 6: Allelopathic effect of A. muricata on seedling growth of E. coracana. [Click here to view] |
 | Table 2: Allelopathic potential of A. muricata on protein, amino acid, carbohydrate, peroxidase, and catalase (mg/g fr. wt.) constituents of V. radiata. [Click here to view] |
 | Table 3: Allelopathic potential of A. muricata on protein, amino acid, carbohydrate, peroxidase, and catalase (mg/g fr. wt.) constituents of E. coracana. [Click here to view] |
All phenolic acids can impact the integrity of DNA and RNA [41,42]. Many phenols and alkaloids, as well as ferulic acid and cinnamic acid, can limit protein synthesis. In barnyard grass co-cultured with allelopathic rice or treated with rice-produced phenolic acids, the expression levels of miRNAs related to plant hormone signal transduction, p53 signaling pathways, nucleotide excision repair, and the peroxisome proliferator-activated receptor were increased [43]. According to Kato-Noguchi et al. [44], the rice allelochemicals momilactone A and B may prevent Arabidopsis seeds from germinating by preventing the breakdown of the storage proteins and cruciferina. These studies support the present findings. Allelochemicals released by donor plants affect receiver plants, and receiver plants respond to the donor plants by changing gene expression. Most phenolic allelochemicals can stimulate Indole-3-Acetic Acid (IAA) oxidase activity and inhibit the reaction of peroxidase with IAA, bound gibberellic acid or IAA to influence endogenous hormone levels [45]. Other plant species’ growth and development are inhibited by extracts from all parts of the plant.
4. CONCLUSION
The present investigation showed that aqueous leaf extract of A. muricata at different concentration levels inhibited seedling growth of V. radiata and E. coracana. However, a low concentration (1%) stimulated germination, seedling length, pigments, starch, protein, amino acid, carbohydrate, peroxidase, and catalase contents of the V. radiata seedlings only. Therefore, the study indicated the beneficial and harmful effects of A. muricata. This plant contains phytochemicals, primarily acetogenins, alkaloids, and phenols. These allelochemicals highly affect the crops. Annona muricata allelochemicals play an important role in the investigation of appropriate farming systems, as well as in the control of weeds, diseases and insects, the alleviation of continuous cropping obstacles, and allelopathic cultivar breeding. Allelochemicals can also be used as ecologically friendly herbicides, fungicides, insecticides, and plant growth regulators, making them extremely useful in sustainable agriculture. Between the two test crops, more inhibition was observed in E. coracana than V. radiata seedlings at 5% aqueous leaf extract treatment. This experiment concluded that the inhibition or stimulation of the seedlings was concentration-dependent and varied in different species. Detailed field studies need to be undertaken to ascertain whether the inhibitory effects noted in the pot culture experiments are expressed in the field too.
5. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
6. FUNDING
There is no funding to report.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Mbagwu FN. The allelopathic effects of crude water extracts of Annona muricata on common weed. Int J Nat Appl Sci 2006;2(4):341–4. CrossRef
2. Oracz K, Bouteau HE, Farrant JM, Cooper K, Belghazi M, Job C, et al. ROS production and protein oxidation as a novel mechanism for seed dormancy alleviation. Plant J 2007;50:452–65. CrossRef
3. Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM. Allelopathy and exotic plant invasion: from molecules and genes to species interaction . Science 2003;301:1377–80. CrossRef
4. Duke SO, Dayan FE, Rimando AM, Schrader KK, Aliotta G, Oliva A, et al. Chemicals from nature for weed management. Weed Sci 2002;50:138–51. CrossRef
5. Duke SO, Dayan FE. Modes of action of phytotoxins from plants. In: Reigosa MJ, Pedrol N, González L (eds.). Allelopathy: A Physiological Process with Ecological Implications, Springer, Amsterdam, The Netherlands, pp 511–36, 2006.
6. He HB, Wang HB, Fang CX, Lin ZH, Yu ZM, Lin WX. Separation of allelopathy from resource competition using rice/barnyard grass mixed-cultures. PLoS One 2012;7:e37201; doi:10.1371/journal.pone.0037201 CrossRef
7. An Y, Ma Y, Shui J. Switchgrass (Panicum virgatum L.) plants and switchgrass residue reduce the biomass and density of associated weeds. Acta Agric Scand 2013;63:107–13. CrossRef
8. Khanh T, Chung M, Xuan T, Tawata S. The exploitation of crop allelopathy in sustainable agricultural production. Agron Crop Sci 2005;191:172–84. CrossRef
9. Scavo A, Restuccia A, Mauromicale G. Allelopathy: principles and basic aspects for agroecosystem control. J Sustain Rev 2018;28:47–101. CrossRef
10. Jacob J, Sreekumar KM, Rekha P. Allelopathic effects of leaf leachates of multipurpose trees on vegetables. Allelopathy J 2001;19:507–16.
11. Peng SL, Wen J, Guo QF. Mechanism and active variety of allelochemicals. Acta Bot Sin 2004;46:757–66.
12. Albuquerque MB, Santos RC, Lima LM, Melo Filho PDA, Nogueira RJMC, Câmara CAG, et al. Allelopathy, an alternative tool to improve cropping systems. Rev Agron Sust Dev 2010;31:379–95. CrossRef
13. Fernandez C, Santonja M, Gros R, Monnier Y, Chomel M, Baldy V, et al. Allelochemicals of Pinus halepensis as drivers of biodiversity in Mediterranean open mosaic habitats during the colonization stage of secondary succession. J Chem Ecol 2013;39:298–311. CrossRef
14. Coria-Téllez AV, Montalvo-Gónzalez E, Yahia EM, Obledo-Vázquez EN. Annona muricata: a comprehensive review on its traditional medicinal uses. Arab J Chem 2016;5:662–91.
15. Lowery. O.H, Rosenbrough, N.J, Farr. A.L., Randall. R. J. “Protein measurement with the folin phenol reagent”. J Biol Chem 1951;193:265–27. CrossRef
16. Venkateshwarlu G, Ravindra V, Challa P. Mangiferin: an allelopathin from mango (Mangifera indica L.) leaves. Allelopathy J 2001;8(2):221–4.
17. Sahoo UK, Jeeceelee L, Vanlalhriatpuia K, Upadhyaya K, Lalremruati JH. Allellopathic effects of leaf leachate of Mangifera indica L. on initial growth parameters of few home garden food crops. World Appl Sci J 2010;10(12):1438–47.
18. Rice EL. Allelopathy. 2nd edition, Academic Press, Orlando, FL, 1984.
19. Shahena S, Rajan M, Chandran V, Mathew L. Allelopathic effect of Wedelia trilobata L., on the germination and growth of Cicer arietinum, Vigna unguiculata, and Vigna radiata seedlings. J App Biol Biotech 2021;9(2):93–114.
20. Murimwa JC, Rugare JT, Mabasa S, Mandumbu R. Allelopathic effects of aqueous extracts of sorghum (Sorghum bicolor L. Moench) on the early seedling growth of Sesame (Sesamum indicum L.) varieties and selected weeds. Int J Agron 2019;2019:1–12. CrossRef
21. Mangao AM, Arreola SLB, Gabriel EVS, Salamanez KC. Aqueous extract from leaves of Ludwigia hyssopifolia (G. Don) exell as potential bioherbicide. J Sci Food Agric 2020;100:1185–94. CrossRef
22. Islam AKMM, Hasan MM, Yeasmin S, Abedin MA, Kader MA, Rashid MH, et al. Bioassay screening of tropical tree sawdust for allelopathic proper ties and their field performance against paddy weeds. Fundam Appl Agric 2019;4(3):906–15. CrossRef
23. Abu-Romman S, Shatnawi M, Shibli R. Allelopathic effects of spurge (Euphorbia hierosolyminata) on wheat (Triticum durum). AEJAES 2010;7(3):298–302.
24. Arowosgabe S, Wintola OA, Afolayan AJ. Phytochemical constituents and allelopathic effect of Aloe ferox Mill. root extract on tomato. JMPR, 2012;6(11):2094–9. CrossRef
25. Morikawa CIO, Miyaura R, de Lourdes Tapia y Figueroa M, Rengifo Salgado EL, Fujii Y. Screening of 170 Peruvian plant species for allelopathic activity by using the sandwich method. Weed Biol Manage 2012;12:1–11. CrossRef
26. Qin B, Perry LG, Broeckling CD, Du J, Stermitz FR, Paschke MW, et al. Phytotoxic allelochemicals from roots and root exudates of leafy spurge (Euphorbia esula L.). Plant Signal Behav 2006;1:323–7. CrossRef
27. Rudrappa T, Quinn WJ, Stanley-Wall NR, Bais HP. A degradation product of the salicylic acid pathway triggers oxidative stress resulting in down-regulation of Bacillus subtilis biofilm formation on Arabidopsis thaliana roots. Planta 2007;226:283–97. CrossRef
28. Yu JH, Zhang Y, Niu CX, Li JJ. Effects of two kinds of allelochemicals on photosynthesis and chlorophyll fluorescence parameters of Solanum melongena L. seedlings. Chin J Appl Econ 2006;17:1629–32.
29. Weir TL, Park SW, Vivanco JM. Biochemical and physiological mechanism mediated by allelochemicals. Curr Opin Plant Biol 2004;7:472–9. CrossRef
30. Wang CM, Chen HT, Li TC, Weng JH, JhanYL, Lin SX. The role of pentacyclic triterpenoids in the allelopathic effects of Alstonia scholaris. J Chem Ecol 2014;40:90–8. CrossRef
31. Poonpaiboonpipat T, Pangnakoran U, Suvunnamek U, Teerarark M, Charoenying P, Laosinwatanna C. Phytotoxic effect of essential oil from Cymbopogan citratus and its physiological effects of banyard grass. Indian Crop Prod 2003;41:403–7. CrossRef
32. Batish RD, Setia N, Singh HP, Kohli RK. Phytotoxicity of lemon-scented eucalypt oil and its potential use as a bioherbicide. Crop Prot 2004;23:1209–14. CrossRef
33. Singh HP, Batish DR, Setia N, Kohli RK. Herbicidal activity of volatile essential oils from Eucalyptus citriodora against Parthenium hysterophorus. Ann Appl Biol 2005;146:89–94. CrossRef
34. Kaur S, Singh HP, Mittal S, Batish DR, Kohli RK. Phytotoxic effects of volatile oil from Artemisia scoparia against weeds and its possible use as a bioherbicide. Ind Crops Prod 2010;32:54–61. CrossRef
35. Scott P. Physiology and behaviour of plants. John Wiley & Sons Inc., Hoboken, NJ, pp 20–1, 2008.
36. Frabboni T, Tarantino D, Petruzzi F, Disciglio G. Bio-herbicidal effects of oregano and rosemary essential oils on chamomile (Matricaria chamomilla L.) crop in organic farming system. Agronomy 2019;9(9):475. CrossRef
37. Ojija F, Arnold SEJ, Treydte AC. Bio-herbicide potential of naturalised Desmodium uncinatum crude leaf extract against the invasive plant species Parthenium hysterophorus. Biol Invasions 2019;21:3641–53. CrossRef
38. Dankov K, Busheva M, Stefanov D, Apostolova EL. Relationship between the degree of carotenoid depletionand function of the photosynthetic apparatus. J Photochem Photobiol B 2009;96:49–56. CrossRef
39. 39.Siyar. S, Majeed A., Muhammad Z.,. Ali .H., and Inayat. N., “Allelopathic effect of aqueous extracts of three weed species on the growth and leaf chlorophyll content of bread wheat,” Acta Ecologica Sinica, vol. 39, no. 1, pp. 63–68, 2019. CrossRef
40. Sofi PA, Djanaguiraman M, Siddique KHM, Prasad PVV. Reproductive fitness in common bean (Phaseolus vulgaris L.) under drought stress is associated with root length and volume. Indian J Plant Physiol 2018;23(4):796–809. CrossRef
41. Zeng RS, Luo SM, Shi YH, Shi MB, Tu CY. Physiological and biochemical mechanism of allelopathy of secalonic acid F on higher plants. Agron J 2001;93:72–9. CrossRef
42. Li ZH, Wang Q, Ruan X, Pan CD, Jiang DA. Phenolics and plant allelopathy. Molecules 2010;15:8933–52. CrossRef
43. Fang C, Li Y, Li C, Li B, Ren Y, Zheng H, et al. Identification and comparative analysis of microRNAs in barnyardgrass (Echinochloa crus-galli) in response to rice allelopathy. Plant Cell Environ 2015;38:1368–81. CrossRef
44. Kato-Noguchi H, Ota K. Biological activities of rice allelochemicals momilactone A and B. J Rice Res 2013;1, 108–12. CrossRef
45. Yang QH, Ye WH, Liao FL, Yin XJ. Effects of allelochemicals on seed germination. Chin J Ecol 2005;24:1459–65.