
Tomato, Solanacearum lycopersicum, is the second most consumed plant organ worldwide. This berry, commonly mistaken to be a vegetable originated from the regions of South America, is the basic indigent for various raw, cooked, or processed foods. Thereby making it the most adopted occupational crop worldwide, either for local use or export [1]. China has been the primary producer of tomato accounting for about 31% of the total global production, followed by India and the United States of America [2]. Tomato belongs to the Solanaceae family, of the plant kingdom, which also comprises various other economically important crops such as the Solanum tuberosum (Potato), Capsicum annuum (Capsicum/Bell peppers), and Solanum melongena (Brinjal/Eggplant) [3]. Tomato constitutes numerous phytocompounds such as the Vitamins A and C, and lycopene, a pigment as well as an antioxidant capable of reducing the risk factors associated with cancer and neurodegenerative diseases [4]. The importance of tomato plant has also been linked to numerous evolutionary researches carried out to understand the metabolic and fruit development processes [5].
The depletion in quality and quantity of tomato crops has been occurring due to unviable cropping with monoculture coupled with expansion of crops to newer regions, leading to the occurrence of a wide range of infections [6]. Most common among such infections in the one caused by root-knot nematode, which is capable of causing severe damages to the plant leading to reduced yield. Globally, an approximation of about >$80 billion losses are incurred annually due to such infection [7]. The root-knot nematodes, which are primarily parasites, may also act specifically as vectors binding to other pathogenic organisms, either way resulting in disease aggregation. Nematodes comprise over 100 species, of which those belonging to the genus Meloidogyne, are associated with tomato crop pathology. Meloidogyne hapla, Meloidogyne naasi, Meloidogyne chitwoodi, and Meloidogyne fallax are commonly found in cold environment. However, Meloidogyne arenaria, Meloidogyne javanica, and Meloidogyne incognita are the most important root-knot nematodes known to cause severe damage in tomato crop in humid condition [8]. The root-knot nematodes adopt the endoparasitic behavior, allowing them to feed and reproduce within the plants and survive in the soil as egg mass and plant debris. During this event, the infective second-stage juveniles move to the root tip to form large spindle-shaped gall like structures. The size of these galls-like masses may vary and reach up to 15 mm in diameter [9].
Since the parasites such as root-knot nematodes cause a major loss in the agricultural system, an integrated use of pest, crop rotation, and biological practices is deemed to be a successful controlling strategy. The consequences of such exercises have been unsatisfactory since about 95% of spray pesticides fail to reach the target parasites and thereby causing a severe effect on the environment and humankind [10]. In addition, the tomato crops have been found to have different degrees of sensitivity toward each Meloidogyne species with the previous studies suggesting that each nematode species exhibited a different degree of parasitism on specific tomato cultivation [11]. While the M. hapla was reported to form small, distinct galls, the M. incognita and M. javanica produced large galls and huge root-swellings which made the infected plants to appear yellow, stunted, and wilt easily [12]. Furthermore, severe infection leads to a reduction of yield and ultimately killing the plants. Furthermore, the root-injuries caused by the nematodes may also act as the point of entry for infectious bacteria and fungi. Particularly in the wilt-resistant varieties, intake of necessary nutrients and water by the host plant is significantly reduced resulting in weak plants with low yields as their physiological processes are affected [13]. Various studies suggest the use of plant resistance inducers to mitigate these adversities and to increase the plant metabolic activity and development of defense mechanism against various parasites. Such an effort would also add value to our understanding of the interactions occurring naturally between plant and root-knot nematodes [14]. The current review provides with an overview of the root-knot nematodes, their mechanism of action against S. lycopersicum, and the possibilities of plant resistance that can act against these nematodes.
1.1. Morphology of Root-knot Nematodes
The root-knot nematodes belong to the genus Meloidogyne. These nematodes are typically elongated, triploblastic, unsegment, and bilaterally symmetrical with pseudocoelomate morphology. They exhibit sexual dimorphism, where the female nematode is up to 3mm long with a soft and pearl white colored body. The neck of a female protrudes anteriorly, while the excretory pore is situated anterior to the median bulb near the stylet base, with the stylet being shorter, and vulva as well as anus is located terminally surrounded by cuticular striae. Paired gonads represent the ovaries. In addition, six large unicellular rectal glands, responsible for gelatinous matrix production, are present in the posterior. Meanwhile, the male nematodes exhibit a vermiform morphology unlike the pear-shaped distended bodies of female nematodes. Male nematodes are shorter, being only 1.2–1.5 mm long. The body of the worm is thin and cylindrical shape [15] and male nematodes consist of a large labial disk that binds to medial lips to form a dorsoventral elongate lip region. Males have a delicate stylet and the developmental factor may influence the pattern shape [Figure 1] [16].
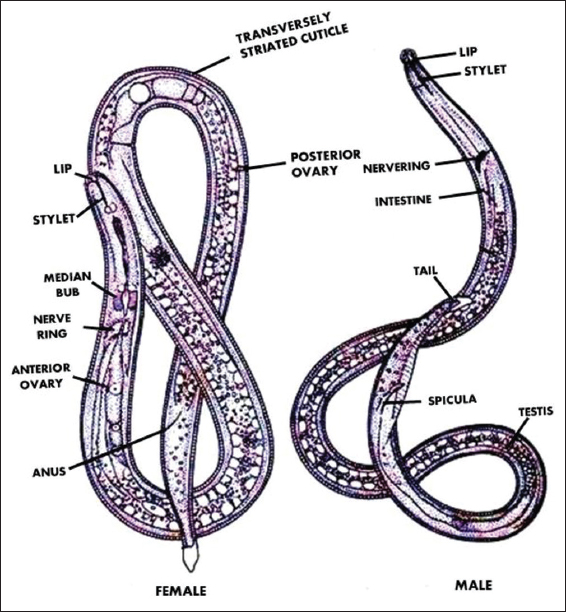 | Figure 1: Morphology of male and female root-knot nematodes. (Source: Handoo et al., 2004) [Click here to view] |
1.2. Life Cycle of Root-knot Nematodes
The lifecycle of all Meloidogyne spp. constitutes six stages [Figure 2] occurring within a duration of about 3–6 weeks. First stage is known as the egg stage, where the nematodes deposit over 1000 eggs in the form of a sticky mass which manages to survive on the soil surface as well as plant residues. In these masses, eggs are clustered to prevent loss of viability due to dehydration. After deposition, the eggs start to multiply to form a first juvenile stage. In the first juvenile stage (J1), coil-like structures appear inside the egg membrane and are characterized by the presence of a stylet. Hence, obtained first juvenile stage gives rise to an infective second-stage juvenile (J2) which hatches within 48 h. Among various nematodes, the Meloidogyne spp. juveniles are only such nematodes that inject esophagus gland secreted substances to apical meristem cells in an attempt to create feeding sites known as giant cells, which act as a channel to pass nutrients from the plant to nematode. Soon after, the second-stage juvenile (J2) becomes adult. In the adult stage, the male nematodes disappear from the roots, while the females swell on maturity producing several numbers of eggs, with or without the involvement of males. Interestingly, the male: female ratio in the nematode population, during this event of reproduction, is balanced generally. This can be attributed to the unique character of immature females to undergo metamorphosis into males, in an instance where the population density does not equate [17].
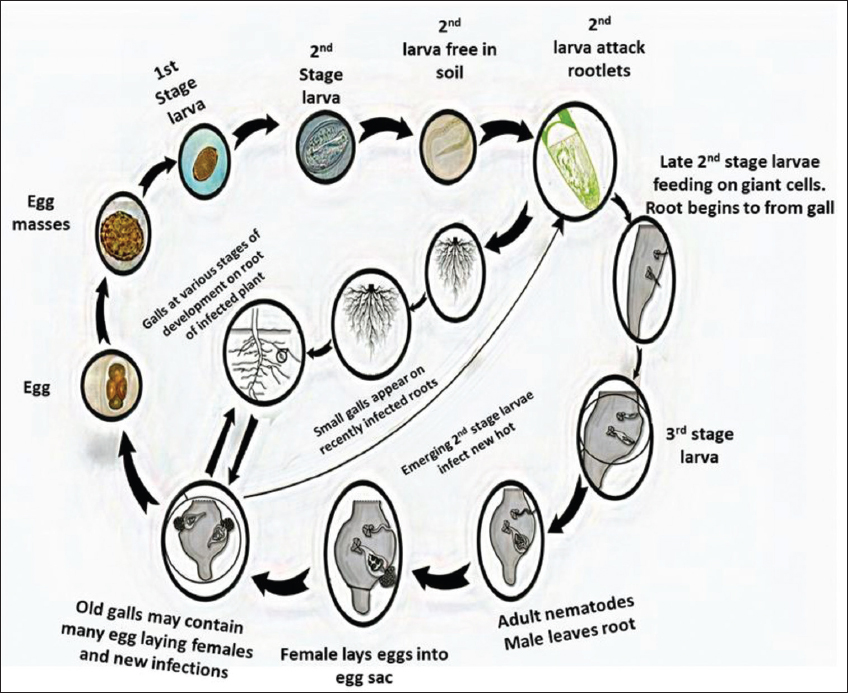 | Figure 2: Complete lifecycle of root-knot nematodes Julie (Source: Nicol et al., 2002) [Click here to view] |
1.3. Symptoms of Infected Plants
In an event of Meloidogyne spp. infection, the S. lycopersicum plants show stunted growth, wilting, yellowish of leaves, and death in severe cases, due to lack of nourishment. The infection of root-knot nematodes causes cell expansion, resulting in swallow or gall like structures as shown in Figure 3. Compared to the other Meloidogyne species, the M. hapla has been reported to form process smaller galls. The root-knot nematodes deprive the plants of their required nutrition thereby increasing their susceptibility other bacterial and fungal invasions. In addition, these plant parasitic nematodes which develop below the soil level are difficult to control by chemical means due to their vast quantities. Several experimental studies, carried out under plot, micro plot, and filed experiments, have suggested that the factors such as: (i) Inoculums type (s) and inoculation technique; (ii) extraction technique and efficiency in laboratory condition; (iii) inoculum density or nematode species and its population level; and (iv) variation in environment and soil type; affect the nematode growth population which is responsible to cause intense damage on tomato cultivation [18,19].
 | Figure 3: The effect of root-knot nematodes causes (a) wilt, (b) yellowish of leaves and (c) gall formation in Solanacearum lycopersicum species (Source: Atamian et al., 2012) [Click here to view] |
1.4. Host Identification through Chemosensory Perception
The root-knot nematodes recognize surrounding habitat through chemosensory impression or through root exudate signals. The tomato plants are capable of producing various chemical gradients across the root nodules, attracting the parasite invasion [20]. A range of phytochemicals have been identified as chemotactic elicitors in Meloidogyne incognita two second-stage juvenile (J2) larvae. The J2 were found to adapt their behavioral responses post-exposure to phytochemicals such as ethephon (Eth), potato root diffusate, and salicylic acid, supporting the existence of long-term habituation traits mediated by the serotonergic-dependent neurotransmission. Understanding of such host-parasite interactions will help develop a pragmatic approach to protect the tomato crops from nematode invasions [21].
1.5. Attractants and Repellants
The J2 root-knot nematode larvae target mostly the root tip of a host. In an attempt to ward off any such infection, the host plants develop unique chemical and physical signals including change (s) in temperature, pH, iron, and carbon dioxide concentrations in the root tips. Such phenomena may either attract or repel the parasite. In addition, carbohydrates, phenolic compounds, chelating compounds, amino acids, and redox potentials of the plant may also regulate the root-knot nematode behavior [22]. The mechanistic intricacies underlying the chemical signaling responsible for such behaviors are poorly understood. Yet, a few studies suggest that when any of the above signaling molecules of chemical nature come in contact with the amphidal duct root-knot nematode, which provides a satisfactory information about root-knot nematode sensory perception, because this is where the phytohormones trigger receptors for induction and secretion of signals by a diffusion process [23]. In Arabidopsis thaliana it was observed that the suppression of ethylene synthesis attracted more M. hapla nematodes toward root tip, whereas an overexpression showed the least attractive capability in the same root-knot nematode species. Indicating that the ethylene and by products of ethylene related pathways repelled the parasites [24]. Based on proven evidences, various studies have been carried out to identify root metabolites detected in root exudates of tomato plants [Table 1]. Of which, methyl salicylate, zeatin, quercetin, specific to tomato root, were found to attract M. incognita J2 larvae, while the metabolites of the ethylene pathway attracted M. hapla, M. incognita, and M. javanica. Meanwhile, palmitic acid and linoleic acid, 1-octanol, trans-cinnamic acid, and p-Coumaric acid showed excellent repellent activity in M. incognita, as other root tip exudates were repellent to J2s, by inducing a quiescence response due to which >80% of the M. incognita nematodes lost motility [32].
Table 1: Compound of root metabolites involved in attraction and repletion of root-knot nematodes.
| Sl. No | species/synthetic chemicals | Root metabolites | Nematodes affected | Effect | Reference |
|---|---|---|---|---|---|
| 1. | Tomato S. lycopersicum | Methyl salicylate | M. incognita | Attractant | [25] |
| 2. | Tomato S. lycopersicum | Zeatin | M. incognita | Attractant | [26] |
| 3. | Tomato S. lycopersicum | Quercetin | M. incognita | Low concentration act as an attractant and higher concentration as a repellent | [26] |
| 4. | Tomato S. lycopersicum and Rice Oryza sativa | Small lipophilic molecules | M. incognita and Heterodera glycine | Repellent | [27] |
| 5. | Tomato S. lycopersicum and barrel clover Medicago truncatula | Metabolites of ethylene pathway | M. hapla, M. javanica, and M. incognita | Ethylene signaling attract more toward hosts compared to wild type | [28] |
| 6. | Synthetic chemicals | Isoamyl alcohol,1- butanol and 2-butanone | M. incognita | Attractant | [29] |
| 7. | Synthetic chemicals | Salicylic acid | M. incognita | Attractant | [30] |
| 8. | Synthetic chemicals | Dopamine | Radophus Similis | Attractant | [30] |
| 9. | Ricinus communis | Palmitic acid and linoleic acid | M. incognita | Repellent | [31] |
| 10. | Synthetic chemicals | 1-octanol | M. incognita | Repellent | [29] |
| 11. | Marigold Tagetespatula, Pepper Capsicum annuum, and Soybean Glycine max | Root exudate compounds | M. incognita and Heteroderaglycines | Repellent to root knot nematode | [32] |
| 12. | Pea Pisum sativum, Snap bean Phaseolus vulgaris, and Alfalfa Medicago sativa | Root tip exudate | M. incognita | Repellent to J2s; induced quiescence response, > 80% of the nematodes lost motility | [32] |
| 13. | Synthetic chemicals | Protocatechuic acid, umbelliferone, caffeic acid, ferulic acid, luteolin, daidzein, genistein, Kaempferol, quercetin, myricetin | Radopholus similis | Repellent andnematicidal | [30] |
| 14. | Synthetic chemicals | Trans-cinnamic acid; p-Coumaric acid | M. incognita | Repellent | [33] |
| 15. | Synthetic chemicals | Salicylic acid, ethephon, vanillic acid, gibberellic acid, indole-3-acetic acid,6-dimethylallylamino purine, mannitol, arginine and lysine | M. incognita | Attractants | [33] |
| 16. | Synthetic chemicals | Salicylic acid, methyl jasmonate, ethephon, indole-3-acetic acid, mannitol | Globodera pallida | Attractants | [33] |
M. incognita: Meloidogyne incognita, Solanacearum lycopersicum: S. lycopersicum.
1.6. Hatching Stimulation
Most of the nematode hatching mechanisms are focused during their parasitic venture, particularly occurring between the host and root-knot nematodes lifecycle. In general, few nematodes depend on the particular host system for their hatching mechanism. The injected juveniles process less energy leads to starvation and finally to death (M. javanica and M. incognita spp.) whereas, few other nematodes enclose dormant eggs that are capable of surviving for decades to reactivate the host-specific active stimulants. There by suggesting the use of hatching stimulants, in the absence of host, to obtain a promising reduction of nematode population. For example, the use of 4-Hydroxyl benzene ethanol to induce hatching in M. incognita; root-exudates derived stimulants as hatching inhibitors of M. hispanica, M. hapla, and M. arenaria has resulted in providing protection to the tomato crops by inhibiting the J2 from penetrating the plant [Table 2] [34].
Table 2: Suppression of root-knot nematode hatching against root metabolites.
| Sl. No. | Plant species/synthetic chemicals | Root metabolites | Nematodes affected | Effect | Reference |
|---|---|---|---|---|---|
| 1. | Welsh onion Allium fistulosum | 4-Hydroxyl benzene ethanol | Meloidogyne incognita | Hatching inhibitor | [35] |
| 2. | Sticky nightshade Solanum sisymbriifolium | Root exudate compounds | Root-knot nematodes | Hatching inhibitor of Meloidogyne hispanica, Meloidogyne hapla and Meloidogyne arenaria; J2s not able to penetrate the plant and highly resistant | [36] |
| 3. | Sticky nightshade Solanum sisymbriifolium | Root exudate compounds | Root-knot nematodes | Hatching inhibitor of Meloidogyne javanica, Meloidogyne arenaria and Meloidogyne hapla; J2s not able to penetrate the plant and highly resistant | [37] |
| 4. | Sticky nightshade Solanum sisymbriifolium | Root exudate compounds | Globodera pallida | Hatching stimulants, reduced number of J2s per cyst and decreased population | [38] |
| 5. | Sticky nightshade Solanum sisymbriifolium and Nightshade Solanum nigrum | Root exudate compounds | Globodera rostochiensis and Globodera pallida | Hatching stimulants, reduced number of J2s per cyst and decreased population | [39] |
| 6. | Nightshade Solanum nigrum | Root exudate compounds | Globodera tabacum | Hatching stimulants | [40] |
1.7. Mode of Action of Root Metabolites in Root Knot Nematodes
Plants employ a defense mechanism to elude from pathogenic nematodes. Such plants generally consist of different proteinase inhibitors, which provide a suitable defense mechanism against those pathogens. These proteinase inhibitors act on nematode proteases, causing starvation due to nutrition depletion. The nematode surfaces consist of various small signaling molecules, such as the Microbe-Associated Molecular Patterns/Nematode-Associated Molecular Patterns (NAMP), which have a root surface pattern recognition receptor (PRR) [41]. On binding to the PRR, triggering of the pattern or NAMP triggered immunity (PTI) occurs [Figure 4] and thereby initiates the Mitogen-activated protein kinase cascade to eventually activate the plant genes responsible for defense. Countering this, the nematodes secrete an effectors molecule, as caroside, along with the effector molecules to hijack the plant cellular system [42]. About time, sensor proteins such as the nucleotide binding leucine-rich repeat proteins are also put into action, while the pathogen detection would have stimulated the R-gene activation to produce the resistance protein. Soon after activation, the resistance protein causes an ion-flux, involving potassium and hydrogen ions to initiate a hypersensitive response (HR) [43]. This HR causes oxidative burst by producing reactive oxygen species mediated cell death to trigger the deposition of lignin and cellulose around the surrounding cells in an attempt to contain the pathogen infection. Such an event is considered to be a short-term response to infection by the plant. The HR also triggers other types of effectors-triggered immunity inducing systemic acquired resistance enhancing the immunity in plants [44]. In an event of failure of HR, activation of hormonal signaling pathway begins, which is a parasite species specific response. While the necrotrophic parasites induce the jasmonic acid pathway [45], the biotrophic parasites will initially induce the salicylic pathway, which further prompts different hormonal pathways and to activate pathogen-related defense gene(s) of the plant [46]. Following this, PR protein expression is upregulated to act against the nematode infection, in combination with various other protein factors.
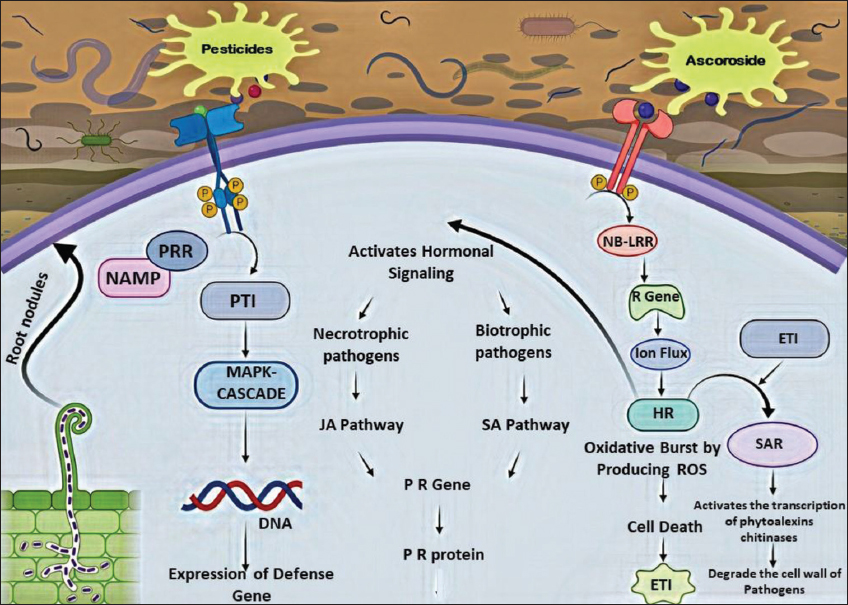 | Figure 4: Mechanism of action root metabolites against root-knot nematodes (Source: Hyong et al., 2016) [Click here to view] |
1.8. Role of RNA Interference (RNAi) in Inhibiting Root-knot Nematode Effectors
RNAi has been an important tool for gene controlling, with an aim to analyze the function of different genes by inhibiting their activity. Such techniques have been demonstrated in various organisms including the root-knot nematodes [47]. The nematodes intake expressible double-stranded RNA molecules, showing a systemic RNAi response in root-knot nematodes as mechanistically explained in Figure 5. Here, the interaction of the particular nematode effector gene(s) with the dsRNA will remains crucial form the infection to be initiated [48]. Studies reveal the importance of controlling root-knot nematodes through host induced gene silencing for a better understanding of possible knockdown on the nematodes. This is also known to control the damage or yield loss in S. lycopersicum species. The functional analysis of these root-knot nematode genes depends on development of a reverse genetic tool acquired by these obligate parasites [49]. Many studies suggest that the RNAi genes are involved in silencing of dicer and pasha genes of M. incognita juveniles using siRNAs [50]. Urwin et al., 2002, studied the plant-parasitic cyst nematode and concluded that octopamine induces the FITC uptake through the secretory system [51]. Iqbal et al., 2016, have explained the behavioral and infectivity variations in the J2 of M. incognita and reported the RNAi influence on such plant-parasitic nematodes [52].
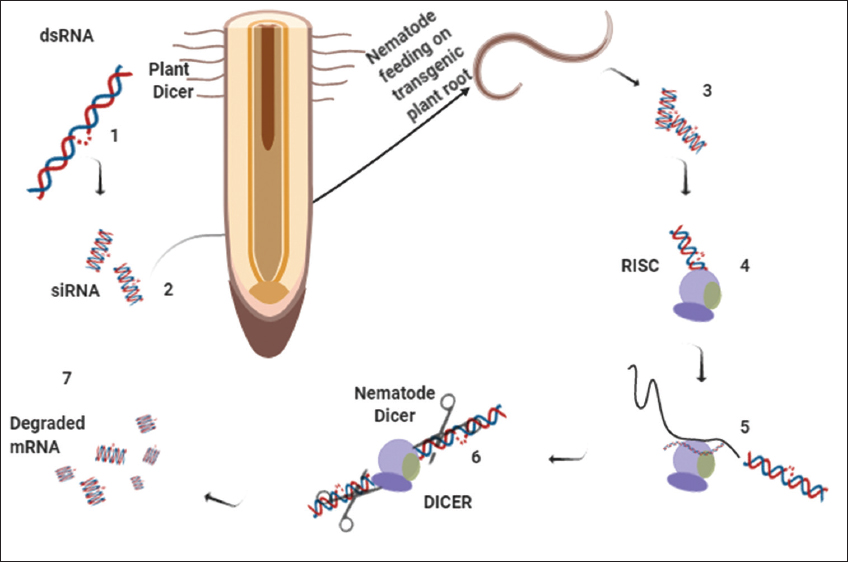 | Figure 5: Mechanism of RNA interference (RNAi), double-stranded RNA (dsRNA) is processed by plant dicer enzyme (1) into (2) once this dsRNA was uptake by these nematodes during feeding (3) once they uptake dsRNA, the nematodes execute dsRNA to shorter interference RNA (4) and also unwind the sense and antisense strands. The RISC complex is filled with antisense strand interacts with the corresponding mRNA in nematodes (5), this makes mRNA to cleave by RISC (6) and leads to degradation of mRNA (7) (Source: Agrawal et al., 2003)[53] [Click here to view] |
1.9. Biological Control
Various bacteria and fungi are also involved in the inhibition of root-knot nematodes. The nematophagous fungi involve in toxin production, behave as endoparasites and/or also capable of trapping the nematodes. Aspergillus spp. and Trichoderma spp. inhibit the tomato plant affecting M. incognita effectively [54]. P. lilacinum fungi (strain 251) were reported to mitigate M. javanica and M. incognita infections apart from reducing gall formation in tomato crop [55]. The bacteria also inhibit root-knot nematodes efficiently [56]. Dababat and Sikora, 2007, concluded that the Fusarium oxysporum Fo162 was capable to induce resistance in tomato plant against M. incognita [57]. Thereby implying that biological control is another important technique to be considered for the control of parasitic infections of S. lycopersicum.
2. CONCLUSION
The economic consequence faced globally due to crop loss through nematode infections is relatively high. These nematodes are unique due to the characteristic feature wherein they are difficult to manage or suppress on introduction to cropland. Various researches are being carried out to understand such nematodes, morphologically, as well as behaviorally, in an attempt to eradicate them. To overcome this, it is essential to understand the mechanistic pathways supporting the nematodes to affect a particular crop, apart from realizing the species-specific damages caused in the host. The imperative use of molecular approaches, such as RNAi, as an alternative to the chemical means can be considered for further evaluation due to their cumulative impact on sustainable as well as profitable agricultural practices.
3. ACKNOWLEDGMENT
The authors acknowledge the authorities of JSS Academy of Higher Education and Research, Mysore, Karnataka, India, for the intellectual and infrastructural support.
4. AUTHOR’S CONTRIBUTION
NB, CD, LKC, and VCV developed the concept. BS, CS, SKP, CD, and NB wrote the review.
5. FUNDING
There is no funding to report.
6. CONFLICTS OF INTEREST
The authors report no conflicts of interest in this work.
REFERENCES
1. Sun S, Luo C, Wei Q, Zhang J. International research status analysis of tomato molecular breeding based on literature survey. Am J Biosci 2016;4:53-7. [CrossRef]
2. Kumar P, Chauhan RS, Grover RK. Economics analysis of tomato cultivation under poly house and open field conditions in Haryana, India. J Appl Natl Sci 2016;8:846-8. [CrossRef]
3. Knapp S, Bohs L, Nee M, Spooner DM.
4. Sundaramoorthy S, Balabaskar P. Biocontrol efficacy of
5. Quinet M, Angosto T, Yuste-Lisbona FJ, Blanchard-Gros R, Bigot S, Martinez JP,
6. Nicol J, Turner D, Coyne L, Den Nijs L, Hockland S, Maafi Z. Current Nematode Threats to World Agriculture. Berlin, Germany:Springer Science Business Media;2011. 21-43. [CrossRef]
7. Seid A, Fininsa C, Mekete T, Decraemer W, Wesemael WM. Tomato (
8. Back MA, Haydock PP, Jenkinson P. Disease complexes involving plant parasitic nematodes and soil borne pathogens. Plant Pathol 2002;51:683-97. [CrossRef]
9. Gharabadiyan F, Jamali S, Komeili HR. Determining of root-knot nematode (
10. Aktar MD, Sengupta D, Chowdhury A. Impact of pesticides use in agriculture:Their benefits and hazards. Interdiscipl Toxicol 2019;1:1-12. [CrossRef]
11. El-Sappah AH, Islam MM, El-Awady HH, Yan S, Qi S, Liu J,
12. Ladner DC, Tchounwou PB, Lawrence GW. Evaluation of the effect of ecologic on root knot nematode, meloidogyne incognita, and tomato plant,
13. Bartlem DG, Jones MG, Hammes UZ. Vascularization and nutrient delivery at root-knot nematode feeding sites in host roots. J Exp Bot 2014;65:1789-98. [CrossRef]
14. Dutta KT, Papolu KP, Banakar P, Choudhary D, Sirohi A, Rao U. Tomato transgenic plants expressing hairpin construct of a nematode protease gene conferred enhanced resistance to root-knot nematodes. Front Microbiol 2015;6:260. [CrossRef]
15. Garcia LE, Sanchez-Puerta MV. Characterization of a root-knot nematode population of
16. Sirca S, Urek G, Karssen G. First report of the root-knot
17. Chukwurah PN, Poku SA, Yokoyama A, Takeda A, Shishido M, Nakamura I. Mitigating root knot nematode propagation on transgenic tobacco via in planta hairpin RNA expression of
18. Berliner J, Ganguly AK, Kamra A, Sirohi A, Pathak H. Effect of elevated carbon dioxide on life cycle of root-knot nematode,
19. Morgan JB, Connolly EL. Plant-soil interactions:Nutrient uptake. Nat Educ Knowl 2013;4:2.
20. Perry RN, Nematol JR. An evaluation of types of attractants enabling plant-parasitic nematodes to locate plant roots. Russ J Nematol 2005;13:83-8.
21. Fleming TR, Maule AG, Fleming CC. Chemosensory responses of plant parasitic nematodes to selected phytochemicals reveal long-term habituation traits. J Nematol 2017;49:462-71.
22. Kihika R, Murungi LK, Coyne D, Nganga M, Hassanali A, Teal PE,
23. Kirwa HK, Murungi LK, Beck JJ, Torto B. Elicitation of differential responses in the root-knot nematode
24. Diez JA, Dusenbery DB. Repellent of root-knot nematodes from exudate of host roots. J Chem Ecol 1989;15:2445-55. [CrossRef]
25. Murungi LK, Kirwa H, Coyne D, Teal PE, Beck JJ, Torto B. Identification of key root volatiles signaling preference of tomato over Spinach by the root knot nematode
26. Dyer S, Weir R, Cox D, Cheseto X, Torto B, Dalzell JJ. Ethylene Response Factor (ERF) genes modulate plant root exudate composition and the attraction of plant parasitic nematodes. Int J Parasitol. 2019;49:13-4. [CrossRef]
27. Dutta TK, Khan MR, Phani V. Plant-parasitic nematode management via bio fumigation using brassica and non-brassica plants:Current status and future prospects. Curr Opin Plant Biol 2019;17:17-32. [CrossRef]
28. Cepulyte R, Danquah WB, Bruening G, Williamson VM. Potent attractant for root-knot nematodes in exudates from seedling root tips of two host species. Sci Rep 2018;8:10847. [CrossRef]
29. Shivakumara TN, Dutta TK, Rao U. A novel in vitro chemo taxis bioassay to assess the response of
30. Wuyts N, Swennen R, De Waele D. Effects of plant phenyl propionic pathway products and selected terpenoids and alkaloids on the behavior of the plant-parasitic nematodes
31. Dong L, Xiaolin L, Huang C, Lu Q, Yao Y, Liu T. Reduced
32. Zhao X, Schmitt M, Hawes MC. Species-dependent effects of border cell and root tip exudates on nematode behavior. Phytopathology 2000;90:1239-45. [CrossRef]
33. Farnier K, Bengtsson M, Becher PG, Witzell J, Witzgall P, ManduricS. Novel bioassay demonstrates attraction of the white potato cyst nematode
34. Clarke AJ, Shepherd AM. Picrolonic acid as a hatching agent for the potato cyst nematode,
35. Li T, Wang H, Xia X, Cao S, Yao J, Zhang L. Inhibitory effects of components from root exudates of Welsh onion against root knot nematodes. PLoS One 2018;13:e0201471. [CrossRef]
36. Meyer SL, Nyczepir AP, Rupprecht SM, Mitchell AD, Martin PA, Brush CW. Tall fescue Jesup (max-Q):
37. Dias MC, Conceiçao IL, Abrantes I, Cunha MJ.
38. Timmermans BG, Vos J, Stomph TJ, Van Nieuwburg J, Van Der Putten PE. Growth duration and root length density of
39. Schenk H, Driessen RA, Gelder RD. Elucidation of the structure of solanoeclepin A, a natural hatching factor of potato and tomato cyst nematodes, by single-crystal X-ray diffraction. Croat Chem Acta 1999;72:593-606.
40. La Mondia JA. Hatch and reproduction of
41. Choi HW, Klessig FD. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol 2016;16:232. [CrossRef]
42. Teixeira MA, Wei L, Kaloshian I. Root-knot nematodes induce pattern-triggered immunity in
43. Dhandaydham M, Charles L, Zhu H, Starr LJ, Huguet T, Cook DR,
44. Niu J, Liu P, Liu Q, Chen C, Guo Q, Yin J,
45. Nahar K, Kyndt T, De Vleesschauwer D, Hofte M, Gheysen G. The jasmonate pathway is a key player in systemically induced defense against root knot nematodes in rice. Plant Physiol 2011;157:305-16. [CrossRef]
46. Matthews FB, Beard H, Brewer E, Kabir S, MacDonald HM, Youssef MR.
47. Lilley CJ, Davies LJ, Urwin PE. RNA interference in plant parasitic nematodes:A summary of the current status. Parasitology 2012;139:630-40. [CrossRef]
48. Njezic B, De Sutter N, Moens M. Interaction of
49. Youssef RM, Kim KH, Haroon SA, Matthews BF. Posttranscriptional gene silencing of the gene encoding aldolase from soybean cyst nematode by transformed soybean roots. Exp Parasitol 2013;134:266-74. [CrossRef]
50. Dalzell JJ, McVeigh P, Warnock ND, Mitreva M, Bird DM, Abad P,
51. Urwin PE, Lilley CJ, Atkinson HJ. Ingestion of double-stranded RNA by preparasitic juvenile cyst nematodes leads to RNA interference. Mol Plant Microbe Interact 2002;15:747-52. [CrossRef]
52. Iqbal S, Fosu-Nyarko J, Jones MG. Attempt to silence genes of the RNAi pathways of the root-knot nematode,
53. Agrawal N, Dasaradhi PV, Mohmmed A, Malhotra P, Bhatnagar RK, Mukherjee SK. RNA interference:Biology, mechanism, and applications. Microbiol Mol Biol 2003;67:657-85. [CrossRef]
54. Goswami BK, Mittal A. Management of root-knot nematode infecting tomato by
55. Kiewnick S, Sikora RA. Biological control of the root-knot nematode
56. Manakau R. Biological control of nematode pests by natural enemies. Ann Rev Phytopathol 1980;81:415-40. [CrossRef]
57. Dababat AA, Sikora RA. Induced resistance by the mutualistic endophyte,