
1. INTRODUCTION
Hexavalent Cr(VI), a toxic heavy metal pollutant, is derived from industrial waste and spent of electrochemical plating, leather tanning, textile, painting, metallurgical industries, and coal power plants, etc [1]. Cr(VI) toxicity is attributed to its water solubility, mobility, and high oxidizing ability in comparison to Cr(III) and Cr(IV) oxidative species [2]. Cr(VI) is reported to be carcinogenic and mutagenic due to the ability to produce reactive oxygen species [3]. In plants, Cr(VI) toxicity leads to poor germination, affects physiological processes such as photosynthesis, water relation, and nutrient deficiency [4,5]. Microbial toxicity of Cr(VI) has also been reported wherein biodiversity in chromate polluted sites has been severely affected [6,7]. Microbe-assisted phytoremediation of Cr(VI) using a combination of plant growth-promoting (PGP) Cr(VI) reducing bacteria residing in the rhizospheric region of phytoaccumulator plants is considered as an economical, viable, and green alternative to the use of physicochemical methods [8]. Many micro-organisms have been described that can remediate toxic Cr(VI) into non-toxic Cr species using different reduction mechanisms while many organisms are described that can tolerate Cr(VI) concentrations in soil but are unable to reduce into non-toxic variants [9–13]. Such Cr(VI) reducing bacteria have also been reported to produce secondary metabolites that also promote plant growth and immunity [8,14,15]. PGP substances produced by rhizospheric or endophytic bacteria play an important role in plant growth and developments as well as improving stress tolerance to heavy metals aiding phytoremediation in many accumulator plants [8,16]. Among the PGP products found to be positive in the isolates, indole acetic acid is important for improving nutrient absorption and lateral and adventitious root development in plants [17]. Siderophores are particularly important in chelating metals such as Fe for plants and reducing metal toxicity [18]. Organic acid byproducts of phosphate solubilization and mobilization by bacteria make insoluble phosphate available to plants [19]. Hydrogen cyanide (HCN) production act by keeping phytopathogens in control and biofilm exopolysaccharides biosorp metals and make them available for reduction while reducing cellular toxicity [20]. The effect of Cr(VI) toxicity on microbial growth and PGP production of rhizospheric bacteria in Vigna mungo system was studied to determine the effect of Cr(VI). It is not understood in previous experimental studies if Cr(VI) negatively affects the production of plant growth stimulating products due to inhibited or reduced microbial growth, and hence decreased PGP production or directly affect the PGP production without affecting growth. In this study, we study the effect of different Cr(VI) concentrations on microbial growth as well as indole acetic acid, siderophore, and phosphate solubilizing enzyme production. P15 bacterization of V. mungo seeds was performed to understand PGP and Cr(VI) remediation physiology in soil microenvironment.
2. MATERIALS AND METHODS
2.1. Microbial growth
Pseudomonas aeruginosa ATCC 15442 [21] was procured from Hi-Media, Mumbai and maintained on tryptone soyapeptone (TS) media at 37°C. For heavy metal experiments, the isolate was plated on tryptone soyapeptone agar (TSA) supplemented with 100 μg/ml Chromium (K2Cr2O7) by standard spread plate method. All media and chemicals used were purchased from HiMedia, India and Merck, India.
2.2. Minimum inhibitory concentration (MIC) and Diphenyl carbazide Assay
P15 was grown in successively increasing Cr(VI) concentrations in TSA medium and growth of plate recorded. Cr (VI) bioreduction ability was estimated using the 1,5 diphenyl carbazide method [15]. Briefly, TS media with varying concentrations of Cr (VI) was inoculated with 24 hours log culture (10% v/v). Aliquots of the sample at different time points were centrifuged and supernatant acidified using 6N H2SO4. 25% (w/v) 1, 5 diphenyl carbazide in acetone was added and change in color to dark pink detected by spectrophotometer (Thermo Scientific, Spectronics) at 540 nm.
2.3. Heavy metal and Halotolerance
Heavy metal tolerance was determined for each isolate on TSA supplemented with 100 μg/ml of different heavy metals: ZnSO4, CdCl2, and Pb(NO3)2 in the absence and presence of 100 μg/ml Cr(VI). Plates were incubated at 37°C for up to 72 hours to measure growth. Halotolerance was similarly determined by incorporating different NaCl concentrations (0.5%–6%) in the absence and presence of 100 μg/ml Cr(VI) in TSA and growth recorded by visual observation.
2.4. Plant growth promontory (PGP) activities
All PGP activity was tested in the presence and absence of varying Cr(VI) concentrations.
2.4.1. Phosphate solubilization
Qualitative phosphate solubilization was checked on Pikovskaya’s agar medium by measuring the clearance zone [22]. The quantitative bioassay was carried out using Erlenmeyer flasks (100 ml) containing 10 ml of NBRIP broth medium inoculated with the 10% bacteria cell [23]. The flasks were incubated for 5 days at 28°C on a shaker at 120 rpm. Available phosphorus content in the filter supernatant, as well as control (supernatant obtained from no bacteria inoculation), was estimated using the vanado-molybdate colorimetric method by measuring the absorbance at a wavelength of 450 nm.
2.4.2. Indole acetic acid (IAA) production
Bacterial production of IAA was tested using tryptone broth containing 0.1% tryptophan. The medium was incubated with 10% log culture (1 × 107 cell/ml) for 2 days at 37°C with shaking. After centrifugation at 10,000 g for 10 minutes, 1 ml of supernatant was mixed with 2 ml of Salkowasky’s reagent and the optical density was measured at 550 nm [24].
2.4.3. Siderophore production
Siderophore production by the isolates was tested qualitatively using the Chroma Azurol Sulphonate (CAS) Media and quantitatively by CAS-Shuttle assay [25]. Succinate medium inoculated with log phase culture was incubated for 48 hours at 37ºC with constant shaking at 120 rpm. Centrifuged cell-free supernatant was mixed with 0.5 ml of CAS reagent, and the absorbance was measured at 630 nm against reference. Siderophore unit in aliquot was calculated by using following formula: [(Ar – As)/Ar] × 100, where Ar is the absorbance at 630 nm of reference (CAS assay solution+ uninoculated media) and As is the absorbance at 630 nm of the sample (CAS assay solution + supernatant) [26].
2.4.4. Ammonia production
Log culture of P15 was grown in peptone water at 37ºC for 2 days. After incubation, 1 ml of Nessler’s reagent was added into each tube and the development of yellow color indicates ammonia production [27].
2.4.5 Biofilm formation
P15 was streaked on Congo red agar media (0.08% congo red; 5% sucrose) in the absence and presence of 100 μg/ml Cr(VI) and incubated at 37ºC for 1 day. The appearance of the black precipitated colony was indicative of biofilm formation [28].
2.5. Plant bioassay
Experiment was performed to evaluate the toxic effects of Cr(VI) on seed germination and seedling development of V. mungo in the presence or absence of isolate [15]. Healthy seeds were firstly surface sterilized in a solution of 0.1% HgCl2 and then washed with sterile water. Log phase culture (108 CFU/ml) was used to inoculate sterile seeds while in control treatments, seeds were placed in sterile saline water in autoclaved Petri dishes. About 3 ml of varying K2Cr2O7 concentrations (100, 250, and 500 μg/ml) was added in seed containing Petri plates lined with Whatman filter paper. Petri plates were placed in room temperature for germination at 28°C for 7 days and moisture content maintained by adding 2 ml sterile water on alternate days. After 1 week, seedlings were harvested to determine shoot length and root length.
2.6. Statistical analysis
Experiments were performed in triplicates. Student’s t-test and ANOVA were used wherever applicable. Data were considered significant when p ≤ 0.05. Statistical analysis was carried out using GraphPad Prism software (version 6.0, USA).
3. RESULTS AND DISCUSSION
3.1. P15 heavy metal and qualitative PGP characterization
Environmental strain P. aeruginosa ATCC 15542 (P15) was treated with media containing increasing concentration of Cr(VI). According to Table 1, the minimum inhibitory concentration of Cr(VI) was found to be 1,250 μg/ml on plate assay. The isolate also has the capability to cross tolerate other heavy metals such as CdCl2, Pb(NO3)2 and ZnCl2, in addition to 100 μg/ml Cr(VI). Tannery effluents are rich in brine and P15 also shows 2% halotolerance (NaCl) in the presence of 100 μg/ml Cr(VI) and 4% halotolerance in the absence of Cr(VI). Qualitative screening for the ability to produce plant growth-promoting products recorded that P15 was positive for ammonia, siderophore, indole acetic acid production biofilm formation even in the presence of 100 μg/ml Cr(VI).
3.2. Cr(VI) reduction and growth kinetics
Figure 1A shows growth kinetic profile of P15 isolate at increasing (100, 250, and 500 μg/ml) Cr(VI) concentration up to day 3 post-inoculation. The isolate shows 23% decrease in growth at 100 and 250 μg/ml Cr(VI), whereas 62% decreased growth is observed at 500 μg/ml Cr(VI) at day 1 post-inoculation compared to Cr(VI) non-amended media (p ≤ 0.001). By day 3, growth at 100 and 250 μg/ml Cr(VI) was found to be comparable to media with no Cr(VI) likely due to detoxification of Cr(VI) in the media while growth remained inhibited by 39% at 500 μg/ml Cr(VI) concentrations. As shown in Figure 1B at 100 μg/ml Cr(VI), bioreduction was found to be 98% within day 1 of growth. Cr(VI) reduction of 500 μg/ml Cr(VI) was found to be at a slower pace and 42% at day 1 and 83% was reduced at day 3, respectively, post inoculation.
 | Table 1: Cr(VI) MIC, cross heavy metal, halotolerance and plant growth promoting properties of Ps. aeruginosa ATCC 15542 (P15). [Click here to view] |
3.3. PGP production and biotoxic effect of Cr(VI)
Pseudomonas aeruginosa P15 was found to be positive for qualitative production of IAA, siderophore, ammonia production, and phosphate solubilization (Table 1). In order to study the effect of increasing Cr(VI) toxicity on growth and PGP production, P15 isolate was treated with 100 and 250 μg/ml Cr(VI) and the quantitative production of IAA, siderophore production, and phosphate solubilization was measured. Figure 2A shows no effect of 100 μg/ml Cr(VI) on growth or IAA production (p ≥ 0.05) with negative effect on growth (p ≤ 0.05) and IAA production (p = 0.01) at 250 μg/ml Cr(VI). Siderophore production was negatively affected in a concentration-dependent manner [Cr(VI) 100 μg/ml, p = 0.002; 250 μg/ml, p = 0.0009] even as growth was unaffected [Cr(VI) 100 μg/ml, p = 0.22; 250 μg/ml, p = 0.06] (Figure 2B). Phosphate solubilization was completely inhibited even at 100 μg/ml Cr(VI) (p ≤ 0.001) with no effect on growth (Figure 2C). Hence, plant growth-promoting activity of P15 was negatively affected by the presence of increasing Cr(VI) concentrations even though the MIC was recorded to be 1,250 μg/ml Cr(VI) and the isolate was capable of reducing Cr(VI) within few days.
3.4. Plant bioassay
In order to study if the decrease in PGP production of indole acetic acid, siderophore, and phosphate solubilization had an effect on plant growth in the presence and absence of Cr(VI), p15 inoculated V. mungo seed germination was studied. As shown in Figure 3, bacterization of seeds with P15 in the absence of Cr(VI) showed a significant increase in the shoot (22.4%) and root length (64.58%) hence showing that PGP effect was evident upon p15 seed treatment (p < 0.05). Concentration-dependent decrease in shoot and root length, respectively, was observed at 100 μg/ml (11.9%, 8.33%), 250 μg/ml (22.3%, 23.5%), and at 500 μg/ml (31.13%, 61.33%, p < 0.05) Cr(VI) concentrations, respectively. In P15 treated seeds at 100 μg/ml treatment, reversal of Cr(VI) toxicity in addition to enhanced growth in the shoot (13.11%) and root (8.3%) length was observed (p < 0.05). However, at both 250 and 500 μg/ml treatment, Cr(VI) toxicity reversal that is similar growth to control untreated seeds is seen without any increase in plant growth parameters. However, in comparison to untreated seeds at 250 and 500 μg/ml Cr(VI), an increase in shoot length but not root length was observed for P15 bacterized seeds (p < 0.05). Bioreduction ability of P15 led to decreased Cr(VI) toxicity to plants with no positive effects of PGP production at higher concentrations. It is likely that Cr(VI) induced repression of rooting hormone IAA production in P15 led to decreased root length. Shoot length increase was likely observed due to decreased Cr(VI) toxicity due to bioremediation potential of P15. This experiment is also indicative that PGP activity without Cr(VI) reduction and heavy metal tolerance abilities will be ineffective in microbe-assisted phytoremediation strategies.
4. DISCUSSION
This study highlights the role of chromate toxicity on micro-organisms by affecting the plant growth-promoting potential of environmental strains. Environmental strain P. aeruginosa ATCC 15542 can produce PGP products, reduce toxic Cr(VI), and help in plant growth restoration of V. mungo in Cr(VI) amended soils only at low Cr(VI) concentrations. This isolate showed minimum growth inhibitory concentration (MICG) of 1,250 μg/ml Cr(VI), halotolerance (2%–4%) as well as cross tolerance to different heavy metals but minimum PGP inhibitory concentration (MICPGP) of 100 μg/ml Cr(VI).
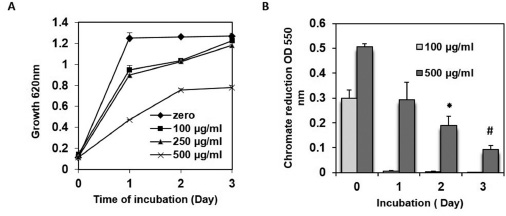 | Figure 1: (A) Growth kinetics of P. aeruginosa ATCC 15442 (P15) at different Cr(VI) concentrations 100, 250, 500 μg/ml) in TSB media up to 3 days post-inoculation (B) Chromium reducing ability at 100 and 500 μg/ml concentration of Cr(VI) in TSB media up to 3 days post-inoculation. Data are mean of three replicates with standard deviation. *p ≤ 0.05, # p ≤ 0.001 [Click here to view] |
 | Figure 2: Plant growth promotion by P. aeruginosa (P15) at different concentrations of Cr(VI) (0, 100, 250 μg/ml. (A) IAA production in tryptone broth with 0.1%tryptophan at 2 days post-inoculation. (B) Siderophore production in CAS media at 2 days post-inoculation. (C) Phosphate solubilization in NBRIP media. Data are mean of three replicates with standard deviation. * p ≤ 0.05, # p ≤ 0.001. [Click here to view] |
Microbial-derived PGP indirectly augments the bioremediation of pollutants as well as help plant growth in nutrient limiting and environmentally unsuitable conditions [8]. IAA, an intercellular communication signaling molecule, also functions as a root elongation hormone [29]. Siderophore production augments bioremediation processes by chelating iron and enhancing tolerance of micro-organisms to abiotic stresses such as pollutants [30]. Phosphate solubilizing organic acids produced by microbes make unavailable complex form of tricalcium phosphorus into products easily utilized by bacteria as well as plants and enhance their nutrition and immunity to environmental pollutants [19]. Yet high concentrations of environmental pollutant such as Cr(VI) also show microbial toxicity [6,7]. We observed that at >100 μg/ml concentrations of Cr(VI), PGP production of indole acetic acid, siderophore, and phosphate solubilization was negatively affected even as the isolate could reduce up to 500 μg/ml Cr(VI) levels to negligible measurable concentrations and grow in the presence 1,250 μg/ml Cr(VI). Hence, it is likely that Cr(VI) and its byproducts may be transcriptionally inhibiting the production of PGP in the presence of stress. Transcriptional regulation of genes in the presence of Cr(VI) stress has been studied to downregulate gene expression [31]. Hence, Cr(VI) concentration even as sub MIC levels shows biotoxic effect on the plant growth-promoting abilities of environmental micro-organisms without affecting their growth. Biotoxic effects of Cr(VI) on plant growth promotion have been reported for Cellulosimicrobium funkei (KM263188) isolated from Phaseolus vulgaris rhizosphere [32]. Heavy metals, such as copper (Cu), chromium (Cr), nickel (Ni), and cadmium, (Cd) were shown to reduce growth and PGP properties of P. aeruginosa and P. fluorescens [33] . Cr and Cd significantly reduced PGP activities, such as IAA, Hydrogen cyanide (HCN), siderophore, and P-solubilization compared to Cu and Ni.
Pseudomonas aeruginosa has until recently been considered a medically important opportunistic pathogen until reports of the rhizospheric and environmental P. aeruginosa as pathogenic or beneficial strains were published [34,35]. 75% Cr(VI) bioreduction at 400 mg/l and tolerance to several heavy metals have been reported for P. aeruginosa CCTCC AB93066 strain [36]. Recently, the role of Pseudomonas sp. (strain CPSB21, capable of Cr6+ bioreduction), in phytoremediation of Cr stress in sunflower plants (but not tomato plants) showed enhanced growth parameters, nutrient uptake, and increase in oxidative stress tolerance [37].
 | Figure 3: Seed bacterization of V. mungo with P. aeruginosa (P15) affects shoot and root length development at day 7 in the absence (control) and presence of different Cr(VI) concentrations (0, 100, 250, and 500 μg/ml). Multiple comparisons among treatments were made using ANOVA with Tukey’s multiple comparison test. Data are mean of three replicates with standard error of mean (SEM). *p ≤ 0.05 of P(15) inoculated in comparison to uninoculated seeds. [Click here to view] |
The use of Cr reducing and PGP producing P. aeruginosa strain OSG41 isolated from rhizospheric soil of mustard plant showed enhanced dry matter accumulation, nodule formation, grain yield, and protein in chickpea compared to non-inoculated plants while decreasing Cr uptake by the plant [38]. The effect of plant growth-promoting P. aeruginosa inhibiting germination of seeds of Zea mays and Triticum aestivum has also been reported [39]. Hence, PGP producing heavy metal stress-tolerant and Cr(VI) reducing bacteria which can enhance growth and reverse Cr toxicity in V. mungo can be used as potentially useful bioinoculant for soil remediation as well as biofertilizers (remedifertilization) for microbe-assisted phytoremediation.
5. CONCLUSION
Plant growth-producing P. aeruginosa ATCC15442 (P15) showed reduction in indole acetic acid, siderophore, and phosphate solubilizing enzymes at sub-inhibitory concentrations of Cr(VI). Hence, the presence of heavy metals such as Cr(VI) affect the beneficial properties of rhizospheric microflora. The Cr(VI) reducing ability of P15 however showed a complete reversal of Cr(VI) toxicity in V. mungo plants even when PGP activity was hindered. Selection and subsequent use of concurrently heavy metal remediating and plant growth-promoting substance producing bacteria are promising isolates to be used for sustainable enhancement of soil productivity.
AUTHOR CONTRIBUTION
VK, RAO, and PDU contributed to design, experimentation, and data acquisition. VK and SDK contributed to the concept, design, data analysis, interpretation, and manuscript preparation.
ACKNOWLEDGMENTS
Financial support from University Grant Commission, New Delhi (357996) to Vineet Kumar is gratefully acknowledged.
CONFLICT OF INTEREST
The authors declared that they have no conflict of interest.
REFERENCES
1. Saha R, Nandi R, Saha B. Sources and toxicity of hexavalent chromium. J Coord Chem 2011;64(10):1782–806. CrossRef
2. Bagchi D. Cytotoxicity and oxidative mechanisms of different forms of chromium. Toxicology. 2002;180(1):5–22. CrossRef
3. Cohen MD, Kargacin B, Klein CB, Costa M. Mechanisms of chromium carcinogenicity and toxicity. Crit Rev Toxicol 1993;23(3):255–81. CrossRef
4. Oliveira H. Chromium as an environmental pollutant: insights on induced plant toxicity. J Bot 2012;2012:1–8. CrossRef
5. Stambulska UY, Bayliak MM, Lushchak VI. Chromium(VI) toxicity in legume plants: modulation effects of rhizobial symbiosis. BioMed Res Int 2018;2018:1–13. CrossRef
6. Shi W, Bischoff M, Turco R, Konopka A. Long-term effects of chromium and lead upon the activity of soil microbial communities. Appl Soil Ecol 2002;21(2):169–77. CrossRef
7. Al-Battashi H, Joshi SJ, Pracejus B, Al-Ansari A. The geomicrobiology of chromium (VI) pollution: microbial diversity and its bioremediation potential. Open Biotechnol J 2016;10(1):379–89. CrossRef
8. Kumar V, Omar R, Kaistha SD. Phytoremediation enhanced with concurrent microbial plant growth promotion and hexavalent chromium bioreduction. J Bacteriol Mycol Open Access 2016;2(6):1–3. CrossRef
9. Thatoi H, Das S, Mishra J, Rath BP, Das N. Bacterial chromate reductase, a potential enzyme for bioremediation of hexavalent chromium: a review. J Environ Manag 2014;146:383–99. CrossRef
10. Kanmani P, Aravind J, Preston D. Remediation of chromium contaminants using bacteria. Int J Environ Sci Technol 2012;9(1):183–93. CrossRef
11. Batool R, Yrjälä K, Hasnain S. Impact of environmental stress on biochemical parameters of bacteria reducing chromium. Braz J Microbiol 2014;45(2):573–83. CrossRef
12. Joutey NT, Sayel H, Bahafid W, El Ghachtouli N. Mechanisms of hexavalent chromium resistance and removal by microorganisms. Rev Environ Contam Toxicol 2015;233:45–69. CrossRef
13. Ramírez-Díaz MI, Díaz-Pérez C, Vargas E, Riveros-Rosas H, Campos-García J, Cervantes C. Mechanisms of bacterial resistance to chromium compounds. Biometals 2008;21(3):321–32. CrossRef
14. Ahemad M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J Genet Eng Biotechnol 2015;13(1):51–8. CrossRef
15. Sagar S, Dwivedi A, Yadav S, Tripathi M, Kaistha SD. Hexavalent chromium reduction and plant growth promotion by Staphylococcus arlettae strain Cr11. Chemosphere 2012;86(8):847–52. CrossRef
16. Rajkumar M, Sandhya S, Prasad MNV, Freitas H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol Adv 2012;30(6):1562–74. CrossRef
17. Gangwar S, Singh VP. Indole acetic acid differently changes growth and nitrogen metabolism in Pisum sativum L. seedlings under chromium (VI) phytotoxicity: implication of oxidative stress. Sci Hortic 2011;129(2):321–8. CrossRef
18. Rajkumar M, Ae N, Prasad MNV, Freitas H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol 2010;28(3):142–9. CrossRef
19. Ahemad M. Phosphate-solubilizing bacteria-assisted phytoremediation of metalliferous soils: a review. 3 Biotech 2015;5(2):111–21. CrossRef
20. Pastorella G, Gazzola G, Guadarrama S, Marsili E, Gugadarrama S, Marsili E. Biofilms: applications in bioremediation. In: Lear G, Lewis G (eds.). Microbial biofilms: current research and applications. Caister Academic Press; 2012. p. 73–98.
21. Wang Y, Li C, Gao C, Ma C, Xu P. Genome sequence of the nonpathogenic Pseudomonas aeruginosa strain ATCC 15442. Genome Announcements. 2014;2(2). CrossRef
22. Pikovskaya R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948;17:362–70.
23. Nautiyal CS. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett 1999;170(1):265–70. CrossRef
24. Patten CL, Glick BR. Bacterial biosynthesis of indole-3-acetic acid. Canad J Microbiol 1996;42(3):207–20. CrossRef
25. Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem 1987;160(1):47–56. CrossRef
26. Payne SM. Detection, isolation, and characterization of siderophores. Methods Enzymol 1994;235:329–44. CrossRef
27. Wani PA, Khan MS, Zaidi A. Chromium-reducing and plant growth-promoting Mesorhizobium improves chickpea growth in chromium-amended soil. Biotechnol Lett 2008;30(1):159–63. CrossRef
28. de Castro Melo P, Ferreira LM, Filho AN, Zafalon LF, Vicente HIG, de Souza V. Comparison of methods for the detection of biofilm formation by Staphylococcus aureus isolated from bovine subclinical mastitis. Braz J Microbiol 2013;44(1):119–24. CrossRef
29. Spaepen S, Vanderleyden J, Remans R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev 2007;31(4):425–48. CrossRef
30. Tak HI, Ahmad F, Babalola OO. Advances in the application of plant growth-promoting rhizobacteria in phytoremediation of heavy metals. Rev Environ Contam Toxicol 2013;223:33–52. CrossRef
31. Zablon HA, VonHandorf A, Puga A. Highlight Article: Chromium exposure disrupts chromatin architecture upsetting the mechanisms that regulate transcription. Exp Biol Med 2019;244(9):752–7. CrossRef
32. Karthik C, Arulselvi PI. Biotoxic effect of chromium (VI) on plant growth-promoting traits of novel Cellulosimicrobium funkei strain AR8 isolated from Phaseolus vulgaris rhizosphere. Geomicrobiol J 2016;34(5):434–42. CrossRef
33. Effect of Heavy metals on Growth and PGPR activity of Pseudomonads. October. 2013;2(5):286–90.
34. Radó J, Kaszab E, Petrovics T, Pászti J, Kriszt B, Szoboszlay S. Characterization of environmental Pseudomonas aeruginosa using multilocus sequence typing scheme. J Med Microbiol 2017;66(10):1457–66. CrossRef
35. Illakkiam D, Shankar M, Ponraj P, Rajendhran J, Gunasekaran P. Genome sequencing of a mung bean plant growth promoting strain of P. aeruginosa with biocontrol ability. Int J Genom 2014;2014:123058. CrossRef
36. Kang C, Wu P, Li Y, Ruan B, Zhu N, Dang Z. Estimates of heavy metal tolerance and chromium(VI) reducing ability of Pseudomonas aeruginosa CCTCC AB93066: chromium(VI) toxicity and environmental parameters optimization. World J Microbiol Biotechnol 2014;30(10):2733–46. CrossRef
37. Gupta P, Rani R, Chandra A, Kumar V. Potential applications of Pseudomonas sp. (strain CPSB21) to ameliorate Cr6+ stress and phytoremediation of tannery effluent contaminated agricultural soils. Sci Rep 2018;8(1):4860. CrossRef
38. Oves M, Khan MS, Zaidi A. Chromium reducing and plant growth promoting novel strain Pseudomonas aeruginosa OSG41 enhance chickpea growth in chromium amended soils. Eur J Soil Biol 2013;56:72–83. CrossRef
39. Tiwari P, Singh JS. A plant growth promoting rhizospheric Pseudomonas aeruginosa strain inhibits seed germination in Triticum aestivum (L) and Zea mays (L). Microbiol Res 2017;8(2). CrossRef