
1. INTRODUCTION
Rice (Oryza sativa L.) is a semi-aquatic monocotyledon domesticated annual crop. It is well-adapted to the Asian continent and has become a dominant crop over there because of the prevailing warm and humid conditions [1]. Its grain called caryopsis contains higher levels of carbohydrate content, vitamins, and minerals. Therefore, it is cultivated immensely in most Asian countries where about 90% of the people consume rice as a primary food in their daily lives. As the world’s population rises continuously, the rate of rice production has to be increased at a higher level because the population rate is predicted to be 9.7 billion in 2050 from the current 8 billion and it could peak at nearly 10.4 billion in the mid-2080s [2]. Already, rice yield stagnation has been initiated by Green Revolution technologies, and along with, the global warming caused by greenhouse gases [3] has become a hard challenge for rice scientists around the world [4]. The impact of drought stress on global rice production in rain-fed low and upland regions is very significant because of the decreasing water supply and increasing intensity of the changing climatic conditions. Growth of rice is hampered by limited leaf growth, reduced leaf area, leaf rolling, leaf drying, thickened leaf size, early senescence, stomatal closure, and cutinized layer on the leaf surface due to water stress [5–8]. Shah et al. [9] explored the development of drought-resistant rice varieties, focusing on the identification of genes that regulate responses to varying water conditions, and the usage of submergence-tolerant strains in flood-prone lowland areas. The study also underscores the important role of physiological, biochemical, and molecular adaptations in strengthening rice’s resilience to drought stress. The paper highlights the significance of marker-assisted breeding and farming practices in semi-arid and rainfed regions to cultivate more drought-resilient strains [9]. Han et al. [10] improved the crop yield of RoLe1 by increasing seed setting rates under moderate drought. Genomic evolution analysis identified a new beneficial allelic variant, proRoLe1−526 [10].
In Asia, approximately 42 million hectares of rice are affected by occasional or frequent drought stress [11]. The rate of yield loss depends on the growth stage, duration, and severity of drought stress. Particularly, significant yield loss is caused by the abortion of the fertilization process due to poor pollen grain germination and pollen tube elongation in the presence of drought stress at the time of flowering [12,13]. Various models and quantitative trait locus (QTL) mapping techniques are employed to identify the location of QTLs within marker intervals. QTL mapping serves as an effective method for uncovering genetic interactions related to disease resistance in plants. In rice, QTL mapping has been performed using F2, recombinant inbred lines, near iso genic lines (NILs), backcross populations, and double haploids to analyze the behavior of specific QTLs, their genetic expression related to grain yield, and additional yield-related traits such as biotic and disease resistance [14]. Fortunately, the continuous efforts of rice scientists of International Rice Research Institute (IRRI) are yielded many major QTL (qDTY 1.1, qDTY 2.1, qDTY 2.2, qDTY 2.3, qDTY 3.1, qDTY 3.2, qDTY 4.1, qDTY 6.1, qDTY 6.2, qDTY 9.1, qDTY 10.1, and qDTY 12.1) associated with increased grain yield in the presence of water scarcity. Among these, qDTY 1.1 [15], qDTY 2.2 [16–18], qDTY 3.1 [11,15], qDTY3.2 [19], qDTY4.1 [18], qDTY6.1 [15], and qDTY 12.1 [12,15,16,19–22] have been reported for its large effects under both lowland and direct seeded upland. In this study, rice varieties, ADT37 incorporated with qDTY3.1 and qDTY2.1 through conventional methods were evaluated for their efficiency of grain yield under nonflood conditions.
2. MATERIALS AND METHODS
2.1. Source of Rice Seeds
A small quantity of rice seeds of ADT37 from Tamilnadu Rice Research Institute, Aduthurai, Tamil Nadu state, and IR81869-B-B-195 from National Rice Research Institute, Cuttack, Odisha state were borrowed. In this study, the ADT37 rice variety was used as a female parent (recipient) for improving drought tolerance at the reproductive stage and IR81869-B-B-195 harboring qDTY3.1 and qDTY2.1 as a male parent (Donor).
2.2. Evaluation of NILs Under NonFlooding Conditions
Here, 32 rice lines of NIL-1 (F1-9.4.3.2) from BC3F2 generation which was incorporated with qDTY3.1 and qDTY2.1in the genetic background of ADT37 through marker-assisted backcrossing method (MABC) (data not shown) were evaluated at field level under moisture conditions. This experiment was carried out in an upland area located within the campus of Kandaswami Kandar’s College, Velur, Namakkal District, Tamil Nadu State, INDIA (11.1121°N 78.0044°E) during Rabi season, 2023–24. Before the commencement of this evaluation, seeds of parental line and NIL were kept at 50°C for 5 days to break dormancy. Seeds were surface sterilized with 70% alcohol for 2 minutes and 2% Clorox for 30 minutes. Then, seeds were kept for incubation at 35°C for 7 days under dark conditions. Germinated seeds were planted in trays for 2 weeks. A 21-day-old 32 seedlings of NIL-1 (F1-9.4.3.2.1-33) were transplanted to the rice field with flooding for 5 cm at a 20 × 20 cm distance. Fertilizers were applied at rates of Urea 50 kg/ha, TSP 62.5 kg/ha, and MOP 50 kg/ha at sowing time. Urea at a rate of 37.5 kg/ha was also applied as a top dressing after 2 and 4 weeks of planting [23]. After 10 days of seedling establishment, the experimental field was water irrigated to avoid water stagnation (nonflooding) for 5 days once until harvesting (Fig. 1).
 | Figure 1. Evaluation of rice lines of NIL-1 under nonflooding conditions. [Click here to view] |
After the maturation of seeds, plants were uprooted carefully and growth rate of rice plants of NIL-1 and parental line in terms of plant height (PH) (measured height from the base of the plant to the top of the latest spikelet on the panicle, excluding awn), root length (RL), flag leaf length (FLL) (measured length from the leaf base to the leaf tip of the fully expanded leaves), flag leaf width (FLW) (measured at the widest point of the leaf), number of tillers (NTs) (counted tillers at the maturity stage), panicle length (PL) (measured from the base of the lowest spikelet to the tip of the latest spikelet on the panicle, excluding awn), panicle dry weight (PDW), number of fertile seeds (NFSs) (counted filled grains in sampled panicles), number of chaff (NC), total number of spikelet (TNS) (counted total number of spikeletes in sampled panicles), 1,000 seeds weight (1,000 SW) (1,000 grains were counted from 5 plants of each replicate and weighed), Single seed length (SSL) (measured using Venire caliper), single seed width (SSW) (measured using Venire caliper), and total plant biomass (TPB) were measured [23]. Seed setting percentages (SS%) were calculated in these NILs, according to IRRI’s scale [24] (Scale 1-More than 80%; 3-61-80%; 5-41-60%; 7-11-40%; 9-Less than 11%).
In breeding programs, MABC is considered an effective tool, free from the ethical concerns associated with transgenic crops. Unlike genetically modified crops, this method relies on molecular markers [25]. MABC is extensively applied in crop improvement, particularly in rice, where it has been instrumental in developing new varieties with resistance to both biotic and abiotic stresses, including bacterial blight, blast, brown planthopper, green leafhopper, gall midge, viral infections, salinity, submergence, drought, cold, semi-dwarfism, and grain quality issues. This technique enables the efficient identification and selection of genes linked to these stresses. Additionally, it is well-accepted by rice farmers due to its ability to recover the recurrent parent genotype within three generations [26]. At the IRRI in the Philippines, rice scientists have identified several key DTY QTLs (qDTY1.1-qDTY12.1) for reproductive-stage drought tolerance, which have been incorporated into popular rice varieties such as IR64, MTU1010, Swarna, Sabitri, TDK1, and Vandana [27].
2.3. Statistical Analysis
Statistical analysis was done for these agronomic characters evaluated in this experiment using mean, standard deviation, and standard error.
3. RESULTS
In this study, we explored 32 NIL-1 rice lines, each incorporated with qDTY3.1 and qDTY2.1, under moisture conditions to assess their growth and water use efficiency. The water use efficiency of these 32 lines impacted the variation in their agronomic traits (Table 1 and Figs. 1 and 2).
 | Table 1. Statistical analysis of agronomic characters of rice lines of NIL-1 under nonflooding conditions. [Click here to view] |
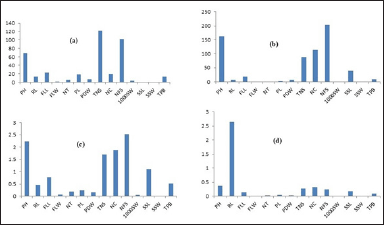 | Figure 2. Growth of improved rice lines incorporated with qDTY3.1 and qDTY2.1 under nonflooding conditions. (a) Mean value; (b) Variance; (c) Standard deviation; (d) Standard error. [Click here to view] |
3.1. Plant Height
For PH character, we noted variations from 47.0 cm (plant 24) to 90.1 cm (plant 10) with a 69.5 cm mean value. The values of variance, standard deviation, standard error, and coefficient variance % were noted to be 161.72%, 2.24%, 0.38%, and 0.03%, respectively.
3.2. Root Length
RL of 32 rice lines varied from 8.0 cm (plant 21) to 19.8 cm (plant 1) with a mean value of 13.5 cm. Here, we noted variance (6.9), standard deviation (0.46), standard error (2.64), and coefficient variance % (0.06%).
3.3. Flag Leaf Length
The length of the flag leaf of 32 rice lines varied from 14.0 cm (plant 4) to 31.5 cm (plant 16) with a mean value of 23.0 cm. The level of variance, standard deviation, standard error, and coefficient variance % of these lines were 19.06%, 0.77%, 0.13%, and 0.03%, respectively.
3.4. Flag Leaf Width
In the case of FLW, we noted variations in the range of 0.7 cm (plant 19)—1.5 cm (plants 1 and 6) with a mean value of 1.2 cm. The level of variance, standard deviation, standard error, and coefficient variance % of these lines were 0.14%, 0.06%, 0.01%, and 0.05%, respectively.
3.5. Number of Tiller
In 32 rice lines, a NTs ranged from 4 (plants 5, 9, 24, and 25) to 8 (plants 29,31, 32) with mean value, variance, standard deviation, standard error, and coefficient variance % of 6.1%, 1.30%, 0.20%, 0.03%, and 0.03%, respectively.
3.6. Panicle Length
In 32 rice lines, the length of the panicle was noted in a range of 16.0 cm (plant 1)–21.5 cm (plant 4) with mean value, variance, standard deviation, standard error, and coefficient variance % of 19.0 cm 2.02%, 0.25%, 0.04%, and 0.01%, respectively.
3.7. Panicle Dry Weight
The rate of the dry weight of panicle in 32 rice lines varied from 4.57 g (plant 9) to 8.24 (plant 33) with mean value, variance, standard deviation, standard error, and coefficient variance % of 6.6 g 6.8%, 0.16%, 0.02%, and 0.02%, respectively.
3.8. Number of Spikelet (NS)
In the case of a TNS, variation was from 102 (plant 22) to 149 (plant 3) with mean value, variance, standard deviation, standard error, and coefficient variance % of 122.1%, 88.2%, 1.7%, 0.28%, and 0.01%, respectively, among 32 rice lines.
3.9. Number of Chaff
A NC of 32 rice lines varied in the range of 4 (plant 30)—54 (plant 25) with mean value, variance, standard deviation, standard error, and coefficient variance % of 19.9%, 114.11%, 1.88%, 0.32%, and 0.09%, respectively.
3.10. Number of Fertile Seed
A NFSs of 32 rice lines was in the range of 72 (plant 24)—124 (plant 2) with mean value, variance, standard deviation, standard error, and coefficient variance % of 102.2%, 204.2%, 2.52%, 0.24%, and 0.02%, respectively.
3.11. 1,000 Seed Weight
In case of 1,000SW of 32 rice lines, we found variation in the range of 3.24 g (plants 2, 4, 5, 6, 7, 8, 9, 10, 13, 15, 16, 18, 19, 20, 21, 25, 27, 29, 30)—4.0 g (plants 11, 12, 17, 22, 24, 26, 31, 33) with mean value, variance, standard deviation, standard error, and coefficient variance % of 3.5 g, 0.09%, 0.05%, 0.008%, and 0.01%, respectively.
3.12. Length of Single Seed (LSS)
LSS of 32 rice lines ranged from 0.6 cm (plant 1, 2, 3, 4, 5, 6, 7, 8, 9,11, 12, 13, 14,16, 17, 18, 19, 20, 21, 23, 24, 25, 26, 28, 29, 30, 31, 32) to 0.7 cm (plant 10, 5, 22, 27, 33) with mean value, variance, standard deviation, standard error and coefficient variance % of 0.68 cm, 38.93%, 1.10%, 0.19%, and 0.02%, respectively.
3.13. Single Seed Width
The width of a single seed of 32 rice lines varied from 0.2 cm (plant 6, 7, 14, 15, 16, 17) to 0.4cm (plant 32) with mean value, variance, standard deviation, standard error, and coefficient variance % 0.28 cm, 0.001%, 0.006%, 0.001%, and 0.02%, respectively.
3.14. Total Plant Biomass
The TPB of 32 rice lines was noted in the range of 5.6 g (plant 1)—19.8 g (plant 7) with mean value, variance, standard deviation, standard error, and coefficient variance % of 13.4 g, 8.95%, 0.52%, 0.09%, and 0.03%, respectively. Rice lines with enhanced grain yield and their association with other agronomic traits under nonflooding conditions (Table 2).
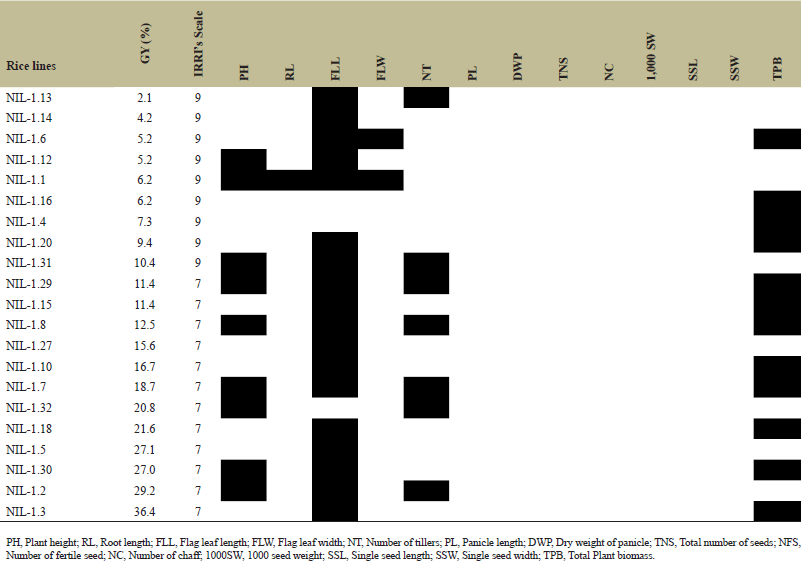 | Table 2. Rice lines with increased grain yield and their linkage with other agronomic characters under nonflooding conditions (Black box indicates the correlation of morphological character with grain yield). [Click here to view] |
4. DISCUSSION
Over the past two decades, many mega rice varieties such as IR64, MTU1010, Swarna, Sabitri, TDK1, PB1, and Vandana have been improved for drought tolerance at reproductive stage incorporating major drought yield QTLs (qDTY 1.1, qDTY 2.1, qDTY 2.2, qDTY 2.3, qDTY 3.1, qDTY 3.2, qDTY 4.1, qDTY 6.1, qDTY 6.2, qDTY 9.1, qDTY 10.1, and qDTY 12.1) [12,15–17,19–21, 25–30] through MABC method worldwide. In the present study, 32 rice lines of NIL-1 incorporated with qDTY3.1 and qDTY2.1 were evaluated under moisture conditions to check their growth and water use efficiency.
In this evaluation, 14 plants (plants 1, 2, 3, 7, 8, 10, 21, 22, 23, 26, 29, 30, 31, 32) were taller than the parental line (ADT37) (78.1 cm) in range of 1.08%–15.29%. A higher rate of PH indicates the water use efficiency of drought-tolerant rice lines and it gets sufficient water and promotes the cell elongation process very fast under stress conditions [31]. Growth of RL is increased in 15 rice lines (plants 1, 2, 5, 6, 9, 11, 15, 18, 25, 26, 27, 28, 29, 31, 32) from 1.3% (plant 27) to 43.3% (plant 1) over the parental line (13.82 cm). Elongation of RL in drought-tolerant rice lines increases due to well adaptation in soil by production of abscisic acid in response to insufficient water [32] and it leads to increased grain yield [33]. In 25 rice lines (plants 1, 2, 3, 6, 7, 8, 10, 11, 12, 13, 15, 16, 18, 19, 20, 21, 22, 23, 26, 27, 28, 29, 30, 31), FLL is increased from 0.96% (plant 29) to 51.4% (plant 16) when compared to the parental line (20.8 cm). In 14 rice lines (plants 1, 6, 7, 8, 9, 12, 13, 16, 17, 21, 23, 24, 28, 31), the rate of FLW increased over the parental line (1.4 cm) in the range of 7.1% (plants 1 and 6)–14.3% (plant 17). In previous studies, the increased rate of flag leaf area along with leaf relative water and leaf pigment content has played a significant role in the drought tolerance of rice lines [6,7,34]34]. Supportively, Kumar et al. [35] reported that the length and width of flag leaves become longer and wider, respectively, in drought-tolerant rice varieties under drought conditions. NT is increased in 24 rice lines (plant 1, 2, 6, 7, 10, 11, 12, 14, 15, 16, 17, 18, 20, 21, 22, 23, 26, 27, 28, 29, 30, 31, 32) from 20.0% (plant 1, 2, 10, 11, 12, 18, 20, 23, 28, 30) to 60.0% (plant 29, 31, 32) over the parental line (5). The NTs increases with the increasing rate of plant growth and photosynthesis in drought-tolerant genotypes promoting the NTs in the presence of sufficient water [8,36,37]. In 17 rice lines (plant 2, 3, 4, 6, 7, 10, 15, 16, 18, 19, 23, 26, 28, 29, 30, 31, 32), PL is increased from 2.16% (plant 23) to 16.21% (plant 4) over the parental line (18.5 cm). The incident of decreased number of panicles and PL occurs significantly in drought-intolerant rice genotypes under drought stress conditions [38]. PDW is found to be increased in 6 rice lines (plant 2, 4, 18, 19, 31, 32) which is superior to the parental line (7.33 gms) in the range of 6.54% (plant 32)—12.41% (plant 33). The rate of NS is higher in 6 rice lines (plant 2, 3, 5, 23, 25, 32) than that of the parental line (130) in the range of 0.7% (plant 23, 25)—14.61% (plant 3). In these, 4 rice lines (plant 23, 24, 25, 26) showed more NS sterility over the parental line (34) up to 58.82% (plant 25). Spikelet sterility is associated with the inability of pollen grains to germinate on dried stigma due to the interruption of phloem loading to ovules by lack of sufficient water [39].
In drought-tolerant genotypes, the rate of grain yield increases with an increasing rate of photosynthesis due to stomatal opening, increased turgor pressure, leaf gas exchange, and CO assimilation under lack of sufficient water [5,34,40]. In this study, NFS is increased in 22 rice lines (plants 1, 2, 3, 4, 5, 6, 7, 8, 10, 12, 13, 15, 16, 18, 19, 20, 27, 29, 30, 31, 32) when compared to parental line (96) in the range of 2.08% (plant 13)—36.45% (plant 3). In 13 plants (plant 1, 3, 7, 11, 12, 14, 17, 22, 23, 24, 26, 28, 31), rate of 1,000SW is increased over the parental line (3.24 g) in the range of 11.41% (plant 1, 2, 7, 24, 23, 28)—23.45% (plant 11, 12, 14, 17, 22, 26, 31, 33). Increased 1,000SW over the parental line indicates the drought tolerance ability of rice lines under drought stress conditions [41]. Moreover, increased activity of antioxidant enzymes such as Peroxidase, Catalase, and Superoxide dismutase in drought-tolerant rice genotypes during drought stress leads to increase grain weight by preventing reactive oxygen species accumulation in plant tissues [42,43].
In the case of SSL, 4 plants (plant 10, 15, 22, 27) were noted for having a higher rate of seed length up to 16.7% when compared to the parental line (0.6 cm). For SSW, we noted only one plant (plant 32) with a wider than parental line (0.3 cm) at the rate of 33.4%. In 18 rice lines (plant 3, 4, 6, 7, 9, 12, 13, 15, 16, 18, 19, 22, 23, 24, 26, 29, 30), TPB is increased when compared to parental line (12.0 g) in the range of 3.87% (plant 13, 29)—53.48% (plant 7). Plant biomass is associated with increased leaf area and fast photosynthesis rate under drought conditions [44,45].
In this study, the rate of SS% in the improved lines with qDTY3.1 and qDTY2.1 is increased in the range of 2.1%–36.4% under nonflooding conditions with IRRI’s scale 7 and 9. Moreover, an increased rate of fertile seed is detected to be linked with other trait of plants such as PH in plant 1, 2, 7, 8, 12, 29, 30, 31, 32, FLL in plants 1, 2, 3, 5, 6, 7, 8, 10, 12, 13, 14, 15, 18, 20, 27, 29, 30, and 31, NTs in plant 1, 2, 7, 8, 29, 31, and 32 and TPB in plant 3, 4, 6, 7, 8, 10, 15, 16, 18, 20, 27, 29, and 30. Among these, many rice lines having increased FLL are associated with grain yield trait followed by TPB rather than PH and NT (Table 2). Moreover, water use efficiency of rice lines influences on expression of agronomic traits differentially among 32 rice lines, for instance, increased rate of PH, FLL, NT, and TPB in plant 7, 8, and 29; PH, FLL, and NT in plant 1, 2, and 31; PH and FLL in plant 12; PH, FLL, and TPB in plant 30; PH and NT in plant 32; FLL and TPB in plant 3; FLL and TPB in plant 6; FLL and TPB in plant 10, 15, 18, 20, 27. It indicates the variations in the adaptive potential of qDTY3.1 and qDTY2.1 in the genetic background of the modern rice variety (ADT37). Very recently, it has been reported that the expression of microRNA (miRNA) affects various transcription factors and traits linked with drought tolerance [46]. In a previous study also, it has been pointed out that two miRNAs (miR2919 and miR156k) regulate the signaling gene of cytokinin and brassinosteriod growth hormone by targeting the genes in the major drought-responsive qDTYs such as qDTY3.1, qDTY1.1, qDTY12.1 associated with drought tolerance at reproductive stage [47]. Marker-assisted breeding, particularly using the backcross method, has been extensively used for introgressing genes governing resistance/tolerance biotic as well as abiotic stresses in rice [48]
5. CONCLUSION
The incorporation of qDTY3.1 and qDTY2.1 into the widely cultivated ADT37 rice variety has successfully improved drought tolerance, particularly at the flowering stage. The study revealed significant variation across agronomic traits among the 32 rice lines, with notable improvements in traits such as PH, RL, NTs, and TPB linked to increased grain yield. These drought-tolerant lines demonstrated up to 36.4% higher yield under nonflooding conditions, making them valuable for low-water farming systems. The enhanced water-use efficiency and adaptability of these lines offer practical benefits for farmers, particularly in drought-prone regions of Southern India, by providing higher yields, reduced reliance on irrigation, and increased economic returns, contributing to overall food security. For future breeding programs, targeting the genetic markers associated with root development, tiller production, and biomass accumulation will be critical for enhancing rice resilience against drought.
6. ACKNOWLEDGMENT
The authors are thankful to the TRRI, Aduthurai, Tamilnadu state, NRRI, Cuttack, Odisha state and Kandaswami Kandar’s College, Velur, Namakkal District, Tamilnadu for providing rice seeds and necessary facilities for conducting the present investigation.
Dembitsky VM, Gloriozova TA, Poroikov VV, Koola MM. QSAR Study of Some Natural and Synthetic Platelet Aggregation Inhibitors and their Pharmacological Profile. J Appl Pharm Sci. 2022; 12(05):039–058. Doi: https://doi.org/10.7324/JAPS.2022.120503
7. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. CONFLICTS OF INTEREST
The authors report no conflicts of interest in this work.
10. FUNDING
There is no funding to report.
11. DATA AVAILABILITY
Research data is available with the authors and shall be provided upon request.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Horie T. Encyclopedia of rice. Noubunkyo (Nousangyosonbunkakyokai, Rural Culture Association Japan), Tokyo, Japan, p 3, 2018 (in Japanese).
2. Gu D, Andreev K, Dupre ME. Major trends in population growth around the world. China CDC Weekly 2021 Jul 7;3(28):604; CrossRef
3. Wilson CA, Mitchell JF. A doubled CO2 climate sensitivity experiment with a global climate model including a simple ocean. J Geophys Res Atmos 1987 Nov 20;92(D11):13315–43; CrossRef
4. Horie T, Shiraiwa T, Homma K, Katsura K, Maeda S, Yoshida H. Can yields of lowland rice resume the increases that they showed in the 1980s? Plant Prod Sci 2005 Jan 1;8(3):259–74; CrossRef
5. Zhu R, Wu F, Zhou S, Hu T, Huang J, Gao Y. Cumulative effects of drought–flood abrupt alternation on the photosynthetic characteristics of rice. Environ Exp Bot 2020 Jan 1;169:103901; CrossRef
6. Mishra SS, Panda D. Leaf traits and antioxidant defense for drought tolerance during early growth stage in some popular traditional rice landraces from Koraput, India. Rice Sci 2017 Jul 1;24(4):207–17; CrossRef
7. Hussain HA, Hussain S, Khaliq A, Ashraf U, Anjum SA, Men S, et al. Chilling and drought stresses in crop plants: implications, cross talk, and potential management opportunities. Front Plant Sci 2018 Apr 10;9:393; CrossRef
8. Quampah A, Wang RenMing WR, Shamsi IH, Ghulam Jilani GJ, Zhang Qiao ZQ, Hua ShuiJin HS, et al. Improving water productivity by potassium application in various rice genotypes. Int J Agric Biol 2011;13(1):9–17.
9. Shah B, Yadav SP, Shrestha M, Khadka A, Dahal K, Neupane B, et al. Drought tolerance in rice (Oryza sativa L.): impact, performance and recent trends. Selcuk J Agric Food Sci 2024;38(1):169–81.
10. Han S, Wang Y, Li Y, Zhu R, Gu Y, Li J, et al. The OsNAC41-RoLe1-OsAGAP module promotes root development and drought tolerance in upland rice. Mol Plant 2024 Sep 2;17(10):1573–93.
11. Venuprasad R, Dalid CO, Del Valle M, Zhao D, Espiritu M, Sta Cruz MT, et al. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor Appl Genet 2009 Dec;120:177–90; CrossRef
12. Barnabás B, Jäger K, Fehér A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 2008 Jan;31(1):11–38; CrossRef
13. Kumar A, Dixit S, Ram T, Yadaw RB, Mishra KK, Mandal NP. Breeding high-yielding drought-tolerant rice: genetic variations and conventional and molecular approaches. J Exp Bot 2014 Nov 1;65(21):6265–78; CrossRef
14. Redona ED, Mackill DJ. Quantitative trait locus analysis for rice panicle and grain characteristics. Theor Appl Genet 1998 May;96:957–63.
15. Vikram P, Swamy BM, Dixit S, Singh R, Singh BP, Miro B, et al. Drought susceptibility of modern rice varieties: an effect of linkage of drought tolerance with undesirable traits. Sci Rep 2015 Oct 13;5(1):14799.
16. Sandhu N, Singh A, Dixit S, Sta Cruz MT, Maturan PC, Jain RK, et al. Identification and mapping of stable QTL with main and epistasis effect on rice grain yield under upland drought stress. BMC Genet 2014 Dec;15:1–5; CrossRef
17. Sandhu N, Torres RO, Sta Cruz MT, Maturan PC, Jain R, Kumar A, et al. Traits and QTLs for development of dry direct-seeded rainfed rice varieties. J Exp Bot 2015 Jan 1;66(1):225–44; CrossRef
18. Swamy BPM, Ahmed HU, Henry A, Mauleon R, Dixit S, Vikram P, et al. Genetic, physiological, and gene expression analyses reveal that multiple QTL enhance yield of rice mega-variety IR64 under drought. PLoS One 2013 May 8;8(5):e62795; CrossRef
19. Yadaw RB, Dixit S, Raman A, Mishra KK, Vikram P, Swamy BM, et al. A QTL for high grain yield under lowland drought in the background of popular rice variety Sabitri from Nepal. Field Crops Res 2013 Mar 20;144:281–7; CrossRef
20. Bernier J, Kumar A, Ramaiah V, Spaner D, Atlin G. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci 2007 Mar;47(2):507–16; CrossRef
21. Dixit S, Swamy BM, Vikram P, Ahmed HU, Sta Cruz MT, Amante M, et al. Fine mapping of QTLs for rice grain yield under drought reveals sub-QTLs conferring a response to variable drought severities. Theor Appl Genet 2012 Jun;125:155–69; CrossRef
22. Mohd Ikmal A, Noraziyah AA, Wickneswari R, Oyebamiji YO. Sub1 and qDTY 3.1 improved tolerance of rice (Oryza sativa L.) lines to drought and submergence stresses. Euphytica 2023 Jun;219(6):60; CrossRef
23. Ranawake AL, Amarasingha UG, Dahanayake N. Agronomic characters of some traditional rice (Oryza sativa L.) cultivars in Sri Lanka. J Univ Ruhuna 2013 Oct 12;1(1):3–9; CrossRef
24. International Rice Research Institute (IRRI). Annual report for 1975. International Rice Research Institute, Los Banos, Philippines, p 29, 2002.
25. Oladosu Y, Rafii MY, Samuel C, Fatai A, Magaji U, Kareem I, et al. Drought resistance in rice from conventional to molecular breeding: a review. Int J Mol Sci 2019 Jul 18;20(14):3519.
26. Bapela T, Shimelis H, Tsilo TJ, Mathew I. Genetic improvement of wheat for drought tolerance: progress, challenges and opportunities. Plants 2022 May 18;11(10):1331.
27. Varshney RK, Bohra A, Yu J, Graner A, Zhang Q, Sorrells ME. Designing future crops: genomics-assisted breeding comes of age. Trends Plant Sci 2021 Jun 1;26(6):631–49.
28. Sandhu N, Jain S, Kumar A, Mehla BS, Jain R. Genetic variation, linkage mapping of QTL and correlation studies for yield, root, and agronomic traits for aerobic adaptation. BMC Genet 2013 Dec;14:1–6; CrossRef
29. Sandhu N, Dixit S, Swamy BP, Raman A, Kumar S, Singh SP, et al. Marker assisted breeding to develop multiple stress tolerant varieties for flood and drought prone areas. Rice 2019 Dec;12:1–6; CrossRef
30. Dhawan G, Kumar A, Dwivedi P, Gopala Krishnan S, Pal M, Vinod KK, et al. Introgression of qDTY1. 1 governing reproductive stage drought tolerance into an elite basmati rice variety “Pusa Basmati 1” through marker assisted backcross breeding. Agronomy 2021 Jan 21;11(2):202; CrossRef
31. Fukai S, Pantuwan G, Jongdee B, Cooper M. Screening for drought resistance in rainfed lowland rice. Field Crops Res 1999 Nov 1;64(1-2):61–74; CrossRef
32. Kim Y, Chung YS, Lee E, Tripathi P, Heo S, Kim KH. Root response to drought stress in rice (Oryza sativa L.). Int J Mol Sci 2020 Feb 22;21(4):1513; CrossRef
33. Uga Y, Sugimoto K, Ogawa S, Rane J, Ishitani M, Hara N, et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat Genet 2013 Sep;45(9):1097–102; CrossRef
34. Farooq M, Kobayashi N, Wahid A, Ito O, Basra SM. 6 strategies for producing more rice with less water. Adv Agron 2009 Jan 1;101(4):351–88; CrossRef
35. Kumar S, Tripathi S, Singh SP, Prasad A, Akter F, Syed MA, et al. Rice breeding for yield under drought has selected for longer flag leaves and lower stomatal density. J Exp Bot 2021 Jun 22;72(13):4981–92; CrossRef
36. Gupta SD, Singh A, Singh P, Kewat RN. Effect of drought stress on carbohydrate content in drought tolerant and susceptible chickpea genotypes. IJCS 2018;6(2):1674–6.
37. Samynathan R, Krishnan M, Venkidasamy B, Subramanian U, Sankaran S, Thiruvengadam R, et al. Comparative transcriptional analysis of genetically superior tea cultivars provides insights into variations in metabolite profiles and biological activities. Sci Hortic 2024 Aug 1;334:113308; CrossRef
38. Bassuony NN, Hassan HM, El-Hissewy AA. Sensitivity of panicle characters of rice (Oryza sativa L.) to drought stress and their assosiation with grain yield. J Plant Prod 2015 Jun 1;6(6):1047–62; CrossRef
39. Wu Y, Fang W, Peng W, Jiang M, Chen G, Xiong F. Sucrose transporter in rice. Plant Signal Behav 2021 Nov 2;16(11):1952373; CrossRef
40. Gupta A, Rico-Medina A, Caño-Delgado AI. The physiology of plant responses to drought. Science 2020 Apr 17;368(6488):266–9; CrossRef
41. Mukamuhirwa A, Persson Hovmalm H, Ortiz R, Nyamangyoku O, Prieto–Linde ML, Ekholm A, et al. Effect of intermittent drought on grain yield and quality of rice (Oryza sativa L.) grown in Rwanda. J Agron Crop Sci 2020 Apr;206(2):252–62; CrossRef
42. Wei X, Cang B, Yu K, Li W, Tian P, Han X, et al. Physiological characterization of drought responses and screening of rice varieties under dry cultivation. Agronomy 2022 Nov 15;12(11):2849; CrossRef
43. Zhong H, Tang W, Li Z, Sonne C, Lam SS, Zhang X, et al. Soil Geobacteraceae are the key predictors of neurotoxic methylmercury bioaccumulation in rice. Nature Food 2024 Apr 11:5:301–11; CrossRef
44. Bunnag S, Pongthai P. Selection of rice (Oryza sativa L.) cultivars tolerant to drought stress at the vegetative stage under field conditions. Am J Plant Sci 2013 Aug 21;4(09):1701; CrossRef
45. Tang W, Bai X, Zhou Y, Sonne C, Wu M, Lam SS, et al. A hidden demethylation pathway removes mercury from rice plants and mitigates mercury flux to food chains. Nat Food 2024 Jan;5(1):72–82; CrossRef
46. Kaur S, Seem K, Kumar D, Kumar S, Kaundal R, Mohapatra T. Biogenesis to functional significance of microRNAs under drought stress in rice: recent advances and future perspectives. Plant Stress 2024 Mar 24;12:100447; CrossRef
47. Kumar D, Ramkumar MK, Dutta B, Kumar A, Pandey R, Jain PK, et al. Integration of miRNA dynamics and drought tolerant QTLs in rice reveals the role of miR2919 in drought stress response. BMC Genomics 2023 Sep 6;24(1):526; CrossRef
48. Dwivedi P, Ramawat N, Dhawan G, Gopala Krishnan S, Vinod KK, Singh MP, et al. Drought tolerant near isogenic lines (NILs) of Pusa 44 developed through marker assisted introgression of qDTY2. 1 and qDTY3. 1 enhances yield under reproductive stage drought stress. Agriculture 2021 Jan 14;11(1):64.