
1. INTRODUCTION
Gibberellic acid (GA3) is a well-recognized plant growth-promoting hormone preferably produced by Fusarium species. Among the Fusarium species, Fusarium fujikuroi is the most frequently used in the production of GA3 biotechnologically [1]. GA3 was first isolated from F. fujikuroi, a predominant causal pathogen of bakane disease of rice. Previously, it was thought that F. fujikuroi was the only species able to produce GA3 [2]. But later, it was reported that Fusarium proliferatum and Fusarium verticillioides also produced GA3 [3,4]. Therefore, it has been considered that F. proliferatum, F. fujikuroi, and F. verticillioides produce GA3 that is related to bakanae disease development. The most important thing, the pathogen’s virulence is associated with GA3 and the virulence depends on excessive secretion of GA3. GA3 is not necessary for the growth and development of fungi but it contributes to the virulence mechanism of fungi [5]. Fusaric acid (FA) is a secondary metabolite (SM) as well as a mycotoxin produced by Fusarium species, and was first recovered from Fusarium heterosporum. FA has also been produced by different types of Fusarium species including F. proliferatum, F. verticillioides, F. fujikuroi, Fusarium subglutinans, Fusarium sambucinum, Fusarium crookwellense, Fusarium oxysporum, F. heterosporum, and Fusarium solani [6]. Among the FA-producing Fusarium species, F. fujikuroi, F. verticillioides, F. proliferatum, and F. subglutinans were found to be involved with rice bakanae disease [7].
Different Fusarium species have been produced GA3 and FA, the most prevalent and oldest known SMs [6,8]. A number of Fusarium species have already been reported for causing bakanae disease of rice [9] and some of them were studied for GA3 and FA production. The other Fusarium species causing bakanae disease of rice need to be investigated for GA3 and FA production. Fusarium species have species-specific SM profiles [2,10]. Therefore, it was essential to find out which strain of Fusarium species produced GA3 and FA in bakanae-infected plants.
The role of GA3 and FA in bakanae symptoms development has been reported previously. GA3 promotes plant elongation [3] and FA has an important role in stunting [4]. FA was also reported for Fusarium wilt in tomato [11]. The exact function of GA3 and FA in the development of disease symptoms is not yet verified, as the specific symptoms caused by the pathogen depend on the production of GA3 and FA, amount of inoculums level, and environmental factors [12,13]. In fact, the SMs profiles of Fusarium species depended on the geographical locations [14,15]. The GA3 and FA-producing Fusarium species causing rice bakanae disease have been reported in several countries including Malaysia [2,4] and India [3]. To the best of our knowledge, previous to our investigations, there are no reports on GA3 and FA production by Fusarium species obtained from bakanae-diseased rice plants in Bangladesh. In fact, bakanae disease has already been reported as an emerging threat to rice production in Bangladesh [16].
The present study, therefore, was conducted to explore 18 Fusarium strains associated with bakanae disease in Bangladesh for GA3 and FA production in vitro, to detect and quantify GA3 and FA produced by the tested strains of Fusarium species, and to confirm the role of GA3 and FA in bakanae disease development.
2. MATERIALS AND METHODS
2.1. Collection and Identification of Fusarium Strains from Bakanae Samples
Diseased rice plants with typical bakanae symptoms such as elongation, lanky, yellowish leaves, stunted and wilted plants were taken from various rice-growing locations in Bangladesh during 2019–2022. Fusarium strains were isolated from bakanae infected rice plants. The isolated strains were identified though morphological and molecular identification methods described by Husna et al. [17].
2.2. Virulence Assay for Fusarium Strains
In total, 121 Fusarium strains belonging to five species were assayed for the evaluation of their pathogenic behavior. Among them, 18 highly virulent Fusarium strains were selected for further analysis through virulence assay in the plant house method described by Husna et al. [17].
2.3. Production of GA3 and FA
2.3.1. Fusarium strains
Based on virulence assay, 18 Fusarium strains were identified as highly virulent and found to be associated with bakanae disease of rice. These strains were used in this study for analysis of GA3 and FA production (Table 1).
2.3.2. Medium preparation
In this study, Czapek-Dox medium was used to stimulate GA3 and FA production by Fusarium strains [18]. For 1 L Czapek-Dox medium preparation, the composition was as follows: NaNO3: 2.0 g, Sucrose: 30.0 g, K2HPO4: 1.0 g, FeSO4 7H2O: 0.01 g, KCl: 0.5 g, MgSO4 7H2O: 0.5 g and distilled water :1 l.
2.3.3. Inoculum preparation
The Fusarium strains were cultured on PDA plates for 7 days at 25°C ± 1°C with a 12:12 hours light:dark cycle. After that, the plates were immersed in 5 ml sterile distill water and spread with a glass rod (hockey stick shape). The conidial suspensions were pooled. The concentration was measured with a hemocytometer and adjusted to 1 × 105 conidia/ml.
2.3.4. Inoculation
A 1 ml suspension of conidia from each strain was added into 100 ml of sterilized Czapek-Dox media. For GA3 production, strains were grown for 5 days in Czapek-Dox medium and shaken at 150 rpm, 30°C in a 12:12 hour light:dark cycle. Besides, for FA production, strains were allowed to grow for 10 days in Czapek-Dox medium and shaken at 180 rpm, 30°C in a 12:12 hour light: dark cycle by using a shaker [19]. The Czapek-Dox media containing 1 ml of sterile distilled water was used as a control.
2.3.5. Extraction, detection and quantification of GA3
2.3.5.1. Chemicals and reagents for GA3
GA3 standard (Sigma Chemical Co., USA), methanol, acetone, phosphoric acid, potassium hydroxide, ethyl acetate, hydrochloric acid (purity> 37%), and ultra-pure water (deionized) were purchased. All these chemicals were HPLC grade.
2.3.5.2. Extraction of GA3
The GA3 was extracted from Fusarium strains according to the method described by Husna et al. [20]. In brief, the fungal mycelial mat was discarded from Czapek-Dox medium and the filtrate pH was corrected to 2.5 with 1N HCL. Then, the filtrate was extracted with equal volumes of ethyl acetate using a separatory funnel. After rotary evaporation (50°C and 10 rpm) of ethyl acetate, the suspended residue was dissolved in 5 ml of acetone and kept at 4°C for ultra-performance liquid chromatography (UPLC) analysis.
2.3.5.3. Preparation of GA3 standard solution
GA3 in powder form was used as a standard for making stock solutions. Standard stock solution (1,000 μg/ml) was prepared in 5 ml methanol and kept at 8°C for further use. Five working standard solutions; 5, 50, 100, 250, and 500 μg/g were prepared in methanol and filtered by 0.2 μm polytetrafluorethylene filter. Finally, the filtrates were preserved at 4°C before use in UPLC.
2.3.5.4. UPLC analysis for GA3
In UPLC analysis, quantification of GA3 was performed according to the method described by Husna et al. [20]. In brief, a C18 column (reversed-phase) was used for chromatographic separations. Methanol (20%) with 10 mM H3PO4 and pH 2.3 KOH was used as the mobile phase [21]. Then, the mobile phase solution was filtrated. The samples were run at 0.1 ml/minute for 10 minutes. The injected rate was 10 μl/sample. GA3 was identified by comparing its retention time and UV spectrum to the standard GA3 sample. For GA3 quantification, the retention durations and peak heights of the samples were compared to those of GA3 standards using a calibration curve.
 | Table 1. Collection, isolation and identification Fusarium strains. [Click here to view] |
2.3.6. Extraction, detection, and quantification of FA
2.3.6.1. Chemicals and reagents for FA
FA standard (ACROS ORGANICS, 99%), methanol, ethanol, phosphoric acid, ethyl acetate, hydrochloric acid (purity> 37%), and ultra-pure water (deionized) were purchased. All these chemicals were HPLC grade.
2.3.6.2. Extraction of FA
The FA was extracted from Fusarium strains according to the method described by Husna et al. [20]. In brief, the fungal mycelial mat was discarded from Czapek-Dox medium and the filtrate pH was corrected to 3.5–4.0 with 2N HCL. Then, the filtrate was extracted with equal volumes of ethyl acetate and shaken well in a separatory funnel. The separatory funnel was kept undisturbed for 30 minutes to divide into two layers. The upper layer containing ethyl acetate was taken in a conical flask. After rotary evaporation (50°C and 10 rpm) of ethyl acetate, the suspended residue was dissolved in 5 ml of ethanol and kept at 4°C for UPLC analysis.
 | Figure 1. Overlay of GA3 standard peaks for different concentrations (5, 50, 100, 250, 500 μg/g). [Click here to view] |
 | Figure 2. UPLC-photodiode array (PDA) chromatogram of GA3 produced by F. fujikuroi (BD047R) detected at 3.7 minutes. [Click here to view] |
2.3.6.3. Preparation of FA standard
FA in powder form was used as a standard for making stock solutions. Standard stock solution (1,000 μg/ml) was prepared in 5 ml methanol and kept at 8°C for further use. Five working standard solutions; 5, 10, 20, 50, and 100 μg/g were prepared in methanol and filtered by 0.2 μm polytetrafluorethylene filter. Finally, the filtrates were preserved at 4°C before use in UPLC.
2.3.6.4. UPLC analysis for FA
In UPLC analysis, quantification of FA was performed according to the method described by Husna et al. [20]. In brief, a C18 column (reversed phase) was used for chromatographic separations. Methanol (20%) and 2% H3PO4 (> 85%) were utilized as mobile phases. An isocratic ratio of mobile phases Methanol: H3PO4 (70%:30%) was utilized [4]. Then, the mobile phase solution was filtrated. The samples were run at 0.1 ml/minute for 10 minutes. The injected rate was 10 μl/sample. FA was identified by comparing its retention time and UV spectrum to the standard FA sample. For FA quantification, the retention durations and peak heights of the samples were compared to those of FA standards using a calibration curve.
2.4. Rice Seedling Test for Shoot Length Assessment
After sterilization, rice seeds (BRRI 29 variety) were soaked for 24 hours in spore suspension (1 × 106 conidia/ml). Twenty-five seeds inoculated with Fusarium strains were placed on filter paper (3 layers, sterile, water-moisture) in Petri dishes, and then the Petri dishes were kept for incubation at 25°C–26°C under a 12 hours light and 12 hours dark, cycle [22]. The seeds treated with sterile distilled water served as control and the test was independently replicated thrice. The elongation and stunting symptoms were compared to the seedlings in control and assessed by measuring the seedling heights at 15 days after incubation.
2.5. Statistical Analysis
The GA3 and FA concentrations were compared among the tested isolates. The standard curve was employed to calculate the concentration of each strain GA3 and FA using Microsoft Excel 10. The correlation was worked out among GA3, FA, and shoot length.
3. RESULTS
3.1. Isolation of Fusarium Strains
From the collected bakanae diseased rice plants, in total 121 strains of Fusarium were successfully isolated and identified through morphological and molecular methods described by Husna et al. [17]. Among them, 18 strains were selected based on virulence assay belonging to 5 Fusarium species (Table 1). These 18 Fusarium strains (14 strains from F fujikuroi, 1 strain from F. proliferatum, 1 strain from F. verticillioides, 1 strain from Fusarium commune, and 1 strain from F. sulawesiense) were further analyzed to produce GA3 and FA.
3.2. GA3 Production
The synthesis of GA3 by 18 Fusarium strains was assessed by comparing their retention time with GA3 standard. A calibration curve using a GA3 standard was created in order to accurately measure the concentration of GA3 in different strains. The retention time of GA3 ranged from 3.6 to 3.9 minutes for both standards and samples (Figs. 1 and 2).
In UPLC analysis, out of 18 strains, 15 strains of Fusarium species were capable of producing GA3 at different concentrations in Czapek-Dox media. In contrast, GA3 was not synthesized in the Czapek-Dox media inoculated with sterile distilled water that served as a control.
The production of GA3 levels ranges from 14.43 to 327.87 μg/g. The highest concentration (327.87 μg/g) of GA3 was produced by the strain BD066R of F. fujikuroi, while the lowest concentration (14.43 μg/g) was produced by the strain BD006R of F. proliferatum. The Fusarium strains of F. fujikuroi species produced higher concentrations of GA3 compared to other species. All the strains of F. fujikuroi produced GA3 except BD117R. Other than F. fujikuroi species, F. proliferatum and F. verticillioides produced GA3 in low concentration while F. commune and F. sulawesiense could not produce GA3 (Table 2).
 | Table 2. GA3 and FA production by Fusarium strains. [Click here to view] |
 | Figure 3. Overlay of FA standard peaks for different concentrations (10, 20 and 50 μg/g) with peaks of methanol and ethanol. Blue color peak indicates methanol and yellow color peak indicates ethanol. [Click here to view] |
3.3. FA Production
The synthesis of FA by 18 Fusarium strains was assessed by comparing their retention time with FA standard (Table 2). A calibration curve using an FA standard was created in order to accurately measure the concentration of FA in different strains. The retention time of FA ranged from 3.6 to 3.9 minutes for both standards and samples (Figs. 3 and 4).
In UPLC analysis, out of 18 strains, 15 strains of Fusarium species were capable of producing FA at different concentrations in Czapek-Dox media. In contrast, FA was not synthesis in the Czapek-Dox media inoculated with sterile distilled water that was served as a control.
The production of FA levels ranged from 47.26 to 254.64 μg/g. The highest concentration (254.64 μg/g) of FA was produced by the strain BD019R of F. commune while the lowest concentration (47.26 μg/g) of FA was produced by the strain BD023R of F. fujikuroi. The Fusarium strains in F. fujikuroi species produced lower concentrations of FA compared to other Fusarium species. All the strains of F. fujikuroi produced FA except BD087R and BD094R. Other than F. fujikuroi species, F. proliferatum, F. verticillioides, and F. commune produced FA while F. sulawesiense could not produce FA (Table 2).
Most of the tested Fusarium strains showed the capability to produce GA3 and FA in different concentrations. The Fusarium strains of F. fujikuroi, in general, produced GA3 at a high level while producing FA comparatively at a lower level. In contrast, F. proliferatum and F. verticillioides strains produced GA3 at a low level but FA at a high level. In F. fujikuroi species, the BD117R strain could not produce GA3 while the BD087R and BD094R strains could not produce FA. The F. commune strain produced only FA, not GA3 and F. sulawesiense could not produce GA3 and FA.
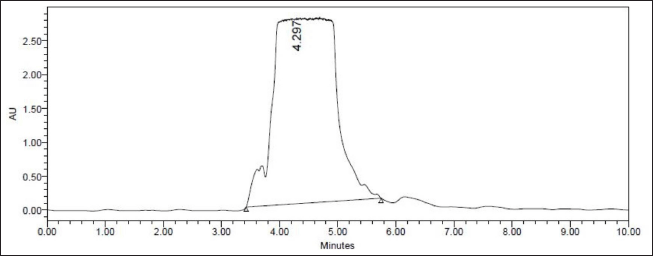 | Figure 4. UPLC-photodiode array (PDA) chromatogram of FA produced by F. fujikuroi (BD043R) detected at 4.2 minutes. [Click here to view] |
 | Figure 5. Rice seedling test for shoot length assessment; (a) The BD080R strain of F. fujikuroi inoculated seedlings showed elongation symptom; (b) The BD006R strain of F. proliferatum inoculated seedlings showed stunting symptom. [Click here to view] |
3.4. Role of GA3 and FA in Bakanae Symptoms Development
All 18 Fusarium strains were analyzed through rice seedling test by shoot length assessment to confirm the function of GA3 and FA in bakanae symptoms development. The characteristic disease symptoms viz. elongation, stunted, rotted, wilted and the leaf tip turned yellow were observed in the seedlings inoculated with Fusarium strains. In the seedling test, the highest shoot length was recorded in the seedlings inoculated with F. fujikuroi BD080R strain which produced a higher amount GA3 (319.98 μg/g) and a lower amount of FA (56.09 μg/g). In contrast, the lowest shoot length was found in the seedlings inoculated with F. proliferatum BD006R strain which produced a higher amount FA (247.33 μg/g) and a lower amount of GA3 (14.43 μg/g). Thus, the produced elongation and stunting symptom were resembled with the typical bakanae symptom (Fig. 5). Again, most of the strains of F. fujikuroi species showed stem elongation symptoms while F. proliferatum, F. verticillioides, and F. commune strains showed stunting symptoms. Overall, the F. fujikuroi strains produced a high concentration of GA3 but a relative low concentration of FA. However, the F. proliferatum and F. verticillioides strains produced a low concentration of GA3 but relatively high concentration of FA.
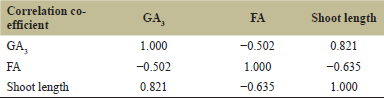 | Table 3. Correlation co-efficient between GA3 and FA produced by strains and shoot length of inoculated rice seedling by strains. [Click here to view] |
The strains BD023R, BD043R, BD047R, BD050R, BD056R, BD058R, BD066R, BD080R, BD087R, BD087R, BD094R, BD111R, BD113R, and BD118R (F. fujikuroi) inoculated rice seedlings showed a positive relationship between the shoot length and GA3 production. The shoot length was high due to the effect of GA3. The hypothesis that GA3 contributes to elongated seedlings was validated by the positive correlation between GA3 production and shoot length (0.821) (Table 3). The strain BD006R (F. proliferatum) and BD013R (F. verticillioides) inoculated rice seedlings produced reduced shoot length due to the effect of a low concentration of GA3 but relative high concentration of FA. Again, the strain BD117R (F. fujikuroi) and BD19R (F. commune) could not produce GA3 but produce only FA in high concentration, therefore these strains inoculated rice seedlings showed reduced shoot length i.e. stunting symptoms. The strain BD026R (F. sulawesiense) inoculated rice seedlings showed about normal shoot growth without the production of any GA3 and FA (Fig. 6).
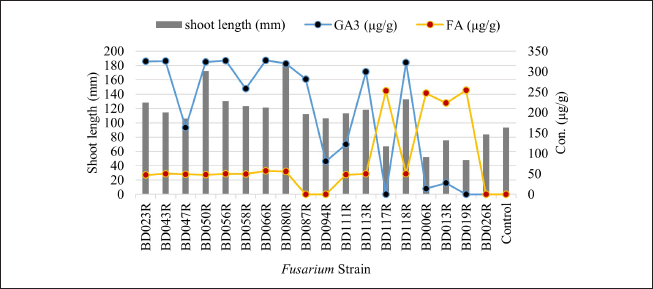 | Figure 6. Effect of GA3 and FA production on shoot length (mm) of rice seedling inoculated with Fusarium strains. [Click here to view] |
4. DISCUSSION
Fusarium species could produce variable amounts of GA3 and FA in bakanae diseased rice plants [23]. In this study, 18 Fusarium strains belonging to 5 Fusarium species isolated from Bangladesh were examined to produce GA3 and FA in vitro conditions. The GA3 and FA produced by the Fusarium strains were detected and quantified as these were associated directly with bakanae symptom development.
In this study, the UPLC analysis was performed to detect and quantify GA3 and FA. The obtained results from UPLC analysis depicted that all the strains of F. fujikuroi, F. proliferatum, and F. verticillioides were able to produce GA3 in high concentration. Interestingly, the strains of F. commune and F. sulawesiense could not produce GA3. Likewise in an earlier report, Fusarium species including F. fujikuroi, F. proliferatum, and F. verticillioides were found to produce GA3 in bakanae diseased rice plants [3,4]. Thus, GA3 can be used as a physiological marker to distinguish one Fusarium species from others.
In contrast, in this study, the Fusarium species like F. proliferatum and F. verticillioides produced FA in high concentration. In earlier, similar findings were observed by several authors [2,3, 24]. Interestingly, F. commune could not produce GA3 but produced FA in high concentration. F. commune was reported to be associated with bakanae disease but there was no report earlier on this pathogen for GA3 and FA production. In fact, GA3 and FA production are governed by the expression of gene clusters responsible for producing GA3 and FA [25]. The variable GA3 and FA profiles were observed in Fusarium species due to the large number of genetic differences between and within the species [26]. In addition, intraspecific variability among the Fusarium strains effect on the variability in the production of GA3 and FA [27]. According to Bhalla et al. [28], the GA3 production by the different strains of Fusarium was varied due to different metabolic pathways of gibberellin production. Besides, the physiological variability of the Fusarium strains was linked with the pathogenic variability. We can get information about the potentiality of a strain to become pathogenic in disease development by understanding the pathogenic and physiological variability of that strain.
In the rice seedling test, we found that the shoots were elongated when inoculated with F. fujikuroi strains that produced GA3 in higher concentrations. In contrast, the stunted shoots were found when inoculated with F. proliferatum, F. verticillioides, and F. commune that produced FA in higher concentration.
The GA3 and FA are responsible for the pathogenicity and the generation of pathogenic diversity in the pathogen. The disease symptoms index mostly relies on the specific type and concentration of GA3 and FA produced, as well as the interactions between the host and pathogen [29,30]. The synthesis of GA3 and FA varies across the strains and is directly linked to their potential to induce bakanae disease [13,23,28]. The symptoms of bakanae disease are related to an imbalance of the phytohormone GA3 [31]. According to Shakeel et al. [9] F. fujikuroi was well recognized for producing GA3. Thus, previous several studies established that the role of GA3 causing elongation symptoms and FA causing stunted seedlings inoculated by Fusarium strains isolated from bakanae disease [3,32]. Thus, our investigation confirmed earlier findings that GA3 and FA are involved in bakanae disease symptom development.
In our study, GA3 contributes to elongate symptom was validated by the positive correlation between GA3 production and shoot length whereas FA production contributes to producing reduced shoot length. According to Puyam et al. [3] the production of GA3 and FA were positively correlated with elongation and stunting type of symptoms, respectively. In addition, Quazi et al. [4] found a correlation between the quantity of FA and the development of bakanae symptoms Wu et al. [33] also found that FA concentration reduced plant height and root length. Additionally, it was reported that bakanae susceptible plants have a negative correlation between plant height and FA concentration [34]. Fusarium strains produced high quantities of FA that prevented GA3 transportation from early leaves to internodes of bakanae-infected plants, and thus, plants became stunted [4].
5. CONCLUSION
The production of GA3 and FA by Fusarium strains causing bakanae disease of rice in Bangladesh was reported in this study as the first attempt. The production of GA3 and FA varied among the 18 Fusarium strains. This study also exposed the co-occurrence of GA3 and FA among the tested strains of F. fujikuroi, F. proliferatum, and F. verticillioides. The development of bakanae disease symptoms was significantly influenced by the concentration levels of GA3 and FA. Thus, the information on the GA3 and FA-producing Fusarium strains for bakanae disease development will contribute in effective management practices for higher rice yields.
6. ACKNOWLEDGMENTS
This research work has been funded by USM Research Grant: 1001.PBIOLOGI.8011097. The Org. for Women in Science for the Developing World (OWSD) also funded.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
11. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation
12. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Wiemann P, Sieber CM, von Bargen KW, Studt L, Niehaus EM, Espino JJ, et al. Deciphering the cryptic genome: genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathog 2013;9(6):e1003475.
2. Zainudin NAIM, Razak AA, Baharuddin S. Secondary metabolite profiles and mating populations of Fusarium species in section Liseola associated with bakanae disease of rice. Malays J Microbiol 2008;4(1):6–13.
3. Puyam A, Pannu PPS, Kaur J, Sethi S. Variability in production of gibberellic acid and fusaric acid by Fusarium moniliforme and their relationship. J Plant Pathol 2017;99(1):103–8.
4. Quazi SAJ, Sariah M, Ahmad ZABM, Hawa J. Detection of fungal metabolites from Bakanae diseased plants and their relationship with Bakanae disease symptoms expression. Am J Biosci Bioeng 2016;4(6):77–89.
5. Volante A, Tondelli A, Aragona M, Valente MT, Biselli C, Desiderio F, et al. Identification of bakanae disease resistance loci in japonica rice though genome wide association study. Rice 2017;10(29):1–16.
6. Bacon CW, Porter JK, Norred WP, Leslie JF. Production of fusaric acid by Fusarium species. Appl Environ Microbiol 1996;62(11):4039–43.
7. Zainudin NAIM, Razak AA, Salleh B. Bakanae disease of rice in Malaysia and Indonesia: etiology of the causal agent based on morphological, physiological and pathogenicity characteristics. J Plant Prot Res 2008;48(4):475–85.
8. Machado CMM, Oishi BO, Pandey A, Soccol CR. Kinetics of Gibberella fujikuroi growth and gibberellic acid production by solid-state fermentation in a packed-bed column. Biotechnol Prog 2004;20:1449–53.
9. Shakeel Q, Mubeen M, Sohail MA, Ali S, Iftikhar Y, Bajwa RT, et al. An explanation of the mystifying bakanae disease narrative for tomorrow’s rice. Front Microbiol 2023;14:1153437.
10. Nelson PE, Desjardins AE, Plattner RD. Fumonisins, mycotoxins produced by Fusarium species: biology, chemistry, and significance. Annu Rev Phytopathol 1993;31:233–52.
11. Singh VK, Singh HB, Upadhyay RS. Role of fusaric acid in the development of ‘Fusarium wilt’ symptoms in tomato: physiological, biochemical and proteomic perspectives. Plant Physiol Biochem 2017;118:320–32.
12. Amatulli MT, Spadaro D, Gullino ML, Garibaldi A. Molecular identification of Fusarium spp. associated with bakanae disease of rice in Italy and assessment of their pathogenicity. Plant Pathol 2010;59(5):839–44.3059.2010.02319.x
13. Kaur J, Pannu PPS, Sucheta S. Morphological, biochemical and molecular characterization of Gibberella fujikuroi isolates causing bakanae disease of basmati rice. J Mycol Plant Pathol 2014;44(1):78–82.
14. Rojas C, Hedden P, Gaskin P, Tudzynski B. The P450–1 gene of Gibberella fujikuroi encodes a multifunctional enzyme in gibberellin biosynthesis. Proc Nat Acad Sci 2001;98(10):5838–44.
15. Mohd Hawa M, Nor NMIM, Azuddin NF, Latiffah Z. Mycotoxin production by Fusarium proliferatum and Fusarium fujikuroi causing stem rot of Hylocereus polyrhizus in Malaysia. Malays Appl Biol 2023;52(3):13–22.
16. Husna A, Miah MA, Nor NMIM. Rice bakanae disease: an emerging threat to rice production in Bangladesh. Asian J Med Biol Res 2020;6(4):608–10.
17. Husna A, Zakaria L, Mohamed Nor NM. Fusarium commune associated with wilt and root rot disease in rice. Plant Pathol 2021;70(1):123–32.
18. Ooi KHH. Pencirian dan pengawalan kimia Fusarium oxysporum, penyebab penyakit layu vascular pada rosel. Ph.D. thesis, Universiti Sains Malaysia, Malaysia, 2002.
19. Burmeister H, Grove MD, Peterson RE, Weisleder D, Plattner RD. Isolation and characterization of two new fusaric acid analogs from Fusarium moniliforme NRRL 13,163. Appl Environ Microbiol 1985;50(2):311–4.
20. Husna A, Miah MA, Zakaria L, Nor NMIM. Fusarium andiyazi, a pathogenic species associated with rice Bakanae disease in Malaysia. Curr Microbiol 2024;81(10):308.
21. Barendse GWM, Van de Werken PH, Takahashi N. High-performance liquid chromatography of gibberellins. J Chromatogr A 1980;198(4):449–55.
22. Qiu J, Lu Y, He D, Lee YW, Ji F, Xu J, et al. Fusarium fujikuroi species complex associated with rice, maize, and soybean from Jiangsu Province, China: phylogenetic, pathogenic, and toxigenic analysis. Plant Dis 2020;104:2193–201.
23. Lale G, Jogdand V, Gadre RV. Morphological mutants of Gibberella fujikuroi for enhanced production of gibberellic acid. J Appl Microbiol 2006;100(1):65–72.
24. Leslie JF, Plattner RD, Desjardins AE, Klittich CJ. Fumonisin B1 production by strains from different mating populations of Gibberella fujikuroi (Fusarium section Liseola). Phytopathology 1992;82(3):341–5.
25. Niehaus EM, Von Bargen KW, Espino JJ, Pfannmüller A, Humpf HU, Tudzynski B. Characterization of the fusaric acid gene cluster in Fusarium fujikuroi. Appl Microbiol Biotechnol 2014;98(4):1749–62.
26. Rabaaoui A, Asta CD, Righetti L, Susca A, Logrieco AF, Namsi A, et al. Phylogeny and mycotoxin profile of pathogenic Fusarium species isolated from sudden decline syndrome and leaf wilt symptoms on date palms (Phoenix dactylifera) in Tunisia. Toxins 2021;13(463):1–19.
27. Gálvez L, Urbaniak M, Waskiewicz A, Stepien L, Palmero D. Fusarium proliferatum—causal agent of garlic bulb rot in Spain: genetic variability and mycotoxin production. Food Microbiol 2017;67:41–8.
28. Bhalla K, Singh SB, Agarwal R. Quantitative determination of gibberellins by high performance liquid chomatography from various gibberellins producing Fusarium strains. Environ Monit Assess 2010;167(1–4):515–20.
29. Ou SH. Rice diseases. 2nd edition, Commonwealth Mycological Institute Kew, Surrey, UK, 1985.
30. Singh R, Sunder S. Foot rot and Bakanae of rice: an overview. Rev Plant Pathol 2012;5:565–604.
31. Quazi SAJ, Meon S, Jaafar H, Ahmad ZAB. The role of phytohormones in relation to bakanae disease development and symptoms expression. Physiol Mol Plant Pathol 2015;90:27–38.
32. Abo-Elnaga HIG, Ahmed NG. Pathogenicity, toxicity and gibberellic acid content of Fusarium moniliforme causing root rot and damping off of pepper. Plant Pathol 2007;6:318–23.
33. Wu H, Bao W, Liu D, Ling N, Ying R, Raza W, et al. Effect of fusaric acid on biomass and photosynthesis of watermelon seedlings leaves. Caryologia 2008;61(3):258–68.
34. Lee YH, Crill JP, Lapis DB. Role of gibberellic acid and fusaric acid in rice plant inoculated with Gibberella fujikuroi (Sawada) Ito and Kimura. Plant Pathol J 1989;5(2):126–30.