1. INTRODUCTION
Wheat (Triticum aestivum L.) is one of the world’s principal food crops. Worldwide, approximately 808 million tons of wheat is produced on 219 million hectares [1]. Approximately half of India’s population depends on wheat for food and other needs; the average land under wheat cultivation was 30.38 million hectares from 2017 to 2022 [2]. However, rapid human population growth indicates that food demand will be more than quadruple by 2050 compared to its current level [3]. Heavy doses of chemical fertilizers, especially nitrogenous and phosphatic fertilizers, are applied to meet the nutrient demand of the crop. The non-judicious use of these chemical fertilizers is non-economical and increases environmental pollution. Potassium (K) and phosphorus (P) are among the most essential nutrients for growth and development. Even though most soils contain sufficient phosphorus and potassium minerals, most of which are bonded to other soil minerals, making them unavailable to plants in a free soluble state.
Under these circumstances, the use of biofertilizers can be a valuable approach to increase their availability. It has been demonstrated that the application of microbial inoculants along with chemical fertilizers is a successful strategy to increase crop yields [4,5]. Due to their beneficial interactions with plants, microbial communities such as rhizobacteria, which live in the rhizosphere and have traits that encourage plant growth, are more prevalent and are participating in this endeavor. The rhizosphere, which is a hotspot for the colonization of beneficial microorganisms, is home to the majority of plant growth-promoting rhizobacteria (PGPRs), which have been reported to enhance plant growth by direct mechanisms such as mineral solubilization, nitrogen fixation, and production of phytohormones such as auxin, gibberellin, and cytokines [4]. Furthermore, PGPR can produce ammonia, hydrogen cyanide (HCN), and bioactive metabolites such as siderophores and biosurfactants [6]. Due to their indirect effects on the synthesis of stress-alleviating enzymes, such as 1-aminocyclopropane-1-carboxylate (ACC) deaminase [7], antibiotic production, and induced systematic resistance, PGPR can modify plant stress responses under abiotic and biotic stress conditions.
P and K solubilization are also among the most commonly reported mechanisms of action linked to increasing the quantity of accessible P and K in the soil that might be readily absorbed by plants under P- and K-limited conditions in the soil [8]. Rhizosphere contains a wide range of P- and K-solubilizing bacterial populations, such as Azotobacter, Acinetobacter, Serratia, Bacillus, Enterobacter, Paenibacillus, Pantoea, Pseudomonas Methylobacterium, Ochrobactrum, Rhizobium, and Acetobacter. Although their population is low, this necessitates the use of outside inoculation to get desirable effects.
Low P and K availability in the soils of many wheat-growing regions also hinders the performance of these crops. An effective way to mitigate the effects of mineral deficits could be to inoculate plants with efficient P- and K-solubilizing PGPR. Moreover, semiarid regions have limitations such as water scarcity, high temperatures, less organic content, and low fertility. The commercially available P and K solubilizers are not effective in such conditions. Therefore, indigenous P- and K-solubilizing microbes isolated from stressful regions can be more appropriate candidates for use as biofertilizers in semiarid conditions. Thus, we aimed to study the effect of newly isolated P- and K-solubilizing bacterial isolates on wheat growth with various combinations of chemical fertilizers under semiarid conditions, emphasizing PGPR as a biologically sustainable fertilizer to enhance plant’s growth under P and K deficiency.
3. RESULTS
3.1. Isolation, Screening, and Quantitative Determination of P- and K-Solubilizing Bacteria
In the present study a total of 87 isolates were initially isolated, which were then screened thrice on Pikovskaya’s and Aleksandrov’s agar medium to confirm their efficacy and stability. Notably, 20 of the 87 isolates were categorized as PSB and KSB based on their P and K solubilization efficacy, while only 8 isolates based on their P- and K-solubilizing index and pH reduction were tested further for different attributes. There were significant differences (p < 0.05) in P solubilization between all strains and the control in the presence of TCP, which varied from 240.48 ± 28.62 to 786.362 ± 21.86 mg/L, and in the presence of RP, soluble P concentrations also varied significantly (p < 0.05) ranging from 26.98 ± 7.65 to 335.996 ± 14.65 mg/L. Similarly, significant differences (p < 0.05) were also recorded between all strains and control in terms of soluble K concentration in the presence of PAS (8.6 to 14.2 mg/L), mica (7.6 to 13.9 mg/L), and K-feldspar (4.3 to 13.3 mg/L) [Figure 1]. Maximum P solubilization in the presence of TCP and RP (768.362 ± 21.86 mg/L and 335.996 ±14.65 mg/L, respectively) was shown by strain SSRP30. However, maximum K was solubilized by strain SSRP9 (14.2 ± 0.069 mg/L) followed by SSRP30 (14.1 ± 0.20 mg/L) in the presence of PAS, whereas in the presence of mica and K-feldspar, maximum solubilization was shown by strain SSRP30 (13.9 ± 0.15 mg/L and 13.3 ± 0.17 mg/L, respectively).
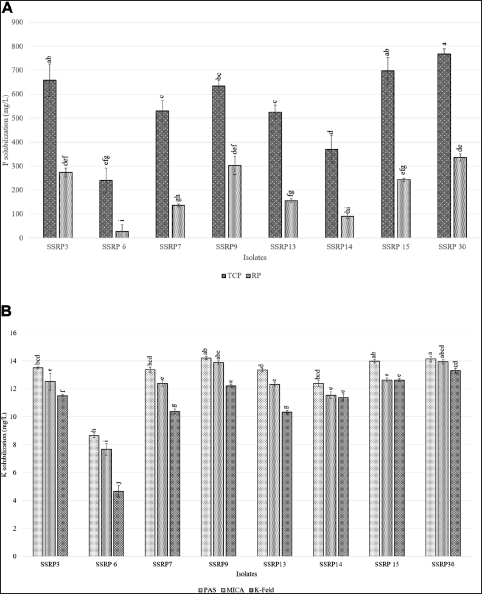 | Figure 1: The graph represents quantified values of (A) phosphate solubilization with two phosphate sources, i.e., TCP and rock phosphate and (B) potassium solubilization with three sources, i.e., PAS, MICA, and K-feldspar. The error bar corresponds to SE (n = 3), bars with different letters indicate a statistically significant difference (Tukey's test: P < 0.05) among different isolates.
[Click here to view] |
3.2. PGP Traits
The PGP traits of the isolates are presented in Table 1. Based on the screening of PGP traits, SSRP30, SSRP9, and SSRP15 showed the highest, i.e., 58.8 ± 01.23, 54.4 ± 3.96, and 50.3 ± 1.36 µg/mL IAA production, respectively, as compared to other test strains [Table 1]. Similarly, highest ACC deaminase was observed in isolate SSRP15 (577.52 ± 0.78 nmol/mg/h) followed by SSRP9 (575.8 ± 3.65 nmol/mg/h) and SSRP3 (497.8 ± 1.40 nmol/mg/h). Furthermore, siderophore production was recorded as the highest for SSRP3 followed by SSRP6 as compared to other isolates; however, all the strains were positive for ammonia and HCN production except SSRP13.
Table 1: Plant growth-promoting attributes of bacterial isolates screened in vitro.
| Isolates | IAA (µg/mL) | ACCD nmol/mg/h | Ammonia | HCN | Siderophores |
|---|
| SSRP3 | 40.77 ± 3.9bc | 497.89 ± 1.40b | +++ | +++ | ++++ |
| SSRP6 | 23.11 ± 2.8ef | 34.60 ± 1.46e | ++ | +++ | ++++ |
| SSRP7 | 35.63 ± 2.2cd | 358.73 ± 2.88c | +++ | + | - |
| SSRP9 | 54.44 ± 2.5a | 575.80 ± 3.65a | ++ | + | - |
| SSRP13 | 16.17 ± 1.6f | 209.99 ± 1.89d | ++ | - | - |
| SSRP14 | 26.34 ± 2.6de | 50.11 ± 2.07e | +++ | - | - |
| SSRP15 | 50.35 ± 2.5ab | 577.52 ± 0.78a | ++ | + | - |
| SSRP30 | 58.8 ± 2.4a | 347.27 ± 1.75c | ++ | ++ | + |
3.3. Molecular Characterization
Notably, 16S rRNA PCR amplicon of ~1500 bp was sequenced and a Basic Local Alignment of concatenated 16S rRNA gene sequence revealed homology with the corresponding organism, and it is presented in Table 2. To assess the taxonomic position of the identified strains, sequences of 14 corresponding bacterial-type strains retrieved from the NCBI were compared. The dendrogram [Figure 2] was generated with MEGA X using the neighbor-joining algorithm with a 50% bootstrap majority-rule consensus tree (1000 replications). Streptomyces griseus was used as an outgroup. With >90% similarity and 0.020 nucleotide substitutions per site, four major groups were clustered.
Table 2: Percent homology with corresponding bacteria and accession numbers of isolates submitted.
| Isolate Name | Percent Similarity | Organisms | Accession No. |
|---|
| SSRP3 | 98.87% | Pseudomonas hunanensis | OR352453 |
| SSRP6 | 98.54% | Pseudomonas aeruginosa | OR352454 |
| SSRP7 | 99.11% | Lelliottia jeotgali | OR352455 |
| SSRP9 | 98.97% | Kosakonia oryzendophytica | OR352456 |
| SSRP13 | 99.44% | Acinetobacter baumannii | OR352457 |
| SSRP14 | 98.75% | Acinetobacter pittii | OR352458 |
| SSRP15 | 99.41% | Enterobacter cloacae | PP140923 |
| SSRP30 | 98.59% | Siccibacter colletis | OR150488 |
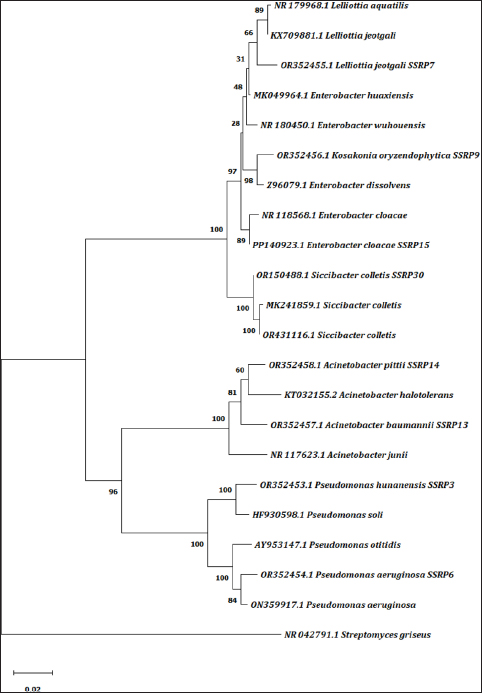 | Figure 2: Phylogenetic tree showing the relationships of native bacterial isolates based on partial 16S rRNA sequences constructed via the neighbor-joining method with the MEGA-XI software. The evolutionary distance was calculated using the maximum composite likelihood method with a bootstrap of 1000 replications. The number on the scale indicates the distance level with relative units.
[Click here to view] |
3.4. Effect of S. colletis Inoculation in Pot Trials
After 5 days of sowing (DAS) in pots, the bacterized wheat seeds displayed a 100% germination rate compared to those in the T1 treatment and the treatment involving non-bacterized seeds, i.e., 98%. The rhizobacterium S. colletis combined with RDF and 50% RDF significantly boosted plant growth. Among the physiological parameters, proline content, RWC, and MTS were found to be nonsignificant between the treatments (P > 0.05). However, there were significant differences in total chlorophyll and sugar contents between the treatments (P < 0.05), with T4 and T6 having the highest levels [Table 3]. The lowest contents of sugar and chlorophyll were observed in the plants from treatment T7, followed by those from treatment T1.
Table 3: Physiological growth parameters of wheat plants under pot and field conditions.
| Pot trial | Field trial |
|---|
| Treatment | Chl a mg/g tissue | Chl b mg/g tissue | Total chl mg/g tissue | Proline µmol/g tissue | Sugar content mg/g tissue | RWC | MTS | Chl a mg/g tissue | Chl b mg/g tissue | Total chl mg/g tissue | Proline µmol/g tissue | Sugar content mg/g tissue | RWC | MTS |
|---|
| T1 | 1.53 ± 0.36c | 0.60 ± 0.05 | 2.23 ± 0.41c | 3.94 ± 0.06 | 6.41 ± 0.37d | 79.13 ± 4.69 | 13.04 ± 1.89 | 1.59 ± 0.26d | 0.60 ± 0.04 | 2.19 ± 0.30d | 3.87 ± 0.07c | 7.35 ± 0.36c | 79.55 ± 3.53 | 12.70 ± 1.86c |
| T2 | 2.62 ± 0.17ab | 0.63 ± 0.04 | 3.51 ± 0.24ab | 4.33 ± 0.24 | 8.49 ± 0.55bc | 84.78 ± 2.16 | 15.38 ± 0.92 | 2.20 ± 0.22c | 0.68 ± 0.11 | 3.14 ± 0.09b | 4.31 ± 0.17b | 8.38 ± 0.73b | 81.97 ± 0.87 | 14.19 ± 1.89abc |
| T3 | 2.61 ± 0.25ab | 0.63 ± 0.40 | 3.30 ± 0.36ab | 4.64 ± 0.34 | 9.04 ± 0.80ab | 82.11 ± 6.67 | 14.21 ± 3.29 | 2.70 ± 0.23b | 0.58 ± 0.29 | 3.28 ± 0.28b | 4.82 ± 0.04a | 9.35 ± 0.76a | 82.74 ± 4.98 | 16.50 ± 2.49a |
| T4 | 3.18 ± 0.28a | 0.67 ± 0.38 | 3.89 ± 0.26a | 4.75 ± 0.55 | 9.46 ± 0.28a | 82.41 ± 7.23 | 16.01 ± 2.35 | 3.19 ± 0.25a | 0.83 ± 0.10 | 3.87 ± 0.20a | 4.96 ± 0.17a | 9.67 ± 0.12a | 82.79 ± 5.62 | 15.13 ± 0.96ab |
| T5 | 1.97 ± 0.09bc | 0.63 ± 0.12 | 2.71 ± 0.16bc | 3.94 ± 0.35 | 7.80 ± 0.30c | 79.78 ± 0.37 | 13.44 ± 1.11 | 2.03 ± 0.07c | 0.66 ± 0.05 | 2.69 ± 0.12c | 4.30 ± 0.21b | 8.31 ± 0.41b | 79.74 ± 0.27 | 13.18 ± 1.31bc |
| T6 | 3.04 ± 0.24a | 0.65 ± 0.13 | 3.86 ± 0.16a | 4.80 ± 0.29 | 8.88 ± 0.67ab | 82.10 ± 4.21 | 13.81 ± 3.16 | 3.15 ± 0.25a | 0.73 ± 0.13 | 3.88 ± 0.12a | 4.83 ± 0.23a | 9.05 ± 0.32a | 81.83 ± 3.24 | 14.32 ± 1.99abc |
| T7 | 1.61 ± 0.43c | 0.62 ± 0.03 | 2.04 ± 0.46c | 4.17 ± 0.52 | 6.63 ± 0.35d | 79.13 ± 4.69 | 13.14 ± 1.69 | - | - | - | - | - | - | - |
| T8 | 2.60 ± 0.19ab | 0.64 ± 0.15 | 3.58 ± 0.15a | 4.44 ± 0.38 | 8.79 ± 0.56ab | 85.48 ± 1.28 | 15.50 ± 0.81 | - | - | - | - | - | - | - |
The biometric parameters of the wheat crops exhibited similar patterns. Treatments T3, T4, and T6 resulted in highly comparable plant biomasses in terms of wet and dry weights (P < 0.05). The NPK concentration in the straw varied across treatments and was significantly higher in T4, followed by T6 and T3 [Table 4], in contrast to the biometric measurements. However, when T7 and T8 were compared with the other treatments for growth characteristics, it was found that T8 tended to be nonsignificant to T4 and T6, respectively, and significantly higher than other treatments.
Table 4: Biometric growth parameters of wheat plants under pot conditions (a) and field conditions (b).
| (a) |
|---|
| Pot Trial |
|---|
| Treatment | Shoot Length cm | Root Length cm | Wet mass g/Plant | Dry mass g/Plant | P % in Straw | N % in straw | K % in straw |
|---|
| T1 | 63.96 ± 0.54d | 26.67 ± 1.27d | 11.34 ± 0.60c | 4.51 ± 0.03c | 0.12 ± 0.00bc | 0.51 ± 0.01c | 0.83 ± 0.03d |
| T2 | 69.23 ± 1.10abc | 31.75 ± 1.27bc | 12.44 ± 0.85abc | 5.46 ± 0.19ab | 0.14 ± 0.01cd | 0.64 ± 0.02b | 0.98 ± 0.10bc |
| T3 | 70.15 ± 1.76ab | 32.17 ± 1.94b | 12.41 ± 0.33abc | 5.89 ± 0.37a | 0.14 ± 0.00cd | 0.56 ± 0.04c | 0.93 ± 0.03c |
| T4 | 71.30 ± 1.82a | 35.56 ± 1.27a | 13.16 ± 0.29a | 6.06 ± 0.26a | 0.16 ± 0.01a | 0.73 ± 0.01a | 1.17 ± 0.03a |
| T5 | 66.46 ± 3.67bcd | 29.63 ± 1.94c | 11.80 ± 0.30bc | 5.07 ± 0.05bc | 0.14 ± 0.00bc | 0.66 ± 0.04b | 1.03 ± 0.05b |
| T6 | 70.07 ± 2.64ab | 32.60 ± 1.94b | 12.70 ± 0.23ab | 5.73 ± 0.25a | 0.14 ± 0.00b | 0.67 ± 0.04b | 1.04 ± 0.07b |
| T7 | 64.41 ± 1.23d | 30.90 ± 0.73bc | 11.62 ± 0.22bc | 4.66 ± 0.04c | 0.13 ± 0.01d | 0.56 ± 0.04c | 0.91 ± 0.02cd |
| T8 | 65.62 ± 3.20cd | 32.17 ± 0.73b | 12.66 ± 0.29ab | 5.60 ± 0.31ab | 0.14 ± 0.01bcd | 0.67 ± 0.04b | 1.00 ± 0.08bc |
| (b) |
|---|
| Field trial |
|---|
| Treatment | Plant Height cm | Ear Height cm | Spikelet/Plant | Ear Weight g/Ear | Grain_yld q/ha | Gtraw_yld q/ha | Biological_yld q/ha | Harvest Index | P% Straw | N% Straw | K% straw | P% grain | N% grain | K% grain |
|---|
| T1 | 90.40 ± 4.04b | 10.42 ± 1.04 | 43.80 ± 12.79 | 1.99 ± 0.37 | 41.89 ± 1.22c | 56.80 ± 1.70c | 98.89 ± 2.86c | 0.41 ± 0.02ab | 0.12 ± 0.00d | 0.51 ± 0.00e | 0.83 ± 0.02c | 0.29 ± 0.01c | 1.02 ± 0.04d | 0.47 ± 0.01d |
| T2 | 94.60 ± 3.58a | 10.76 ± 0.45 | 49.80 ± 5.72 | 2.42 ± 0.51 | 45.15 ± 2.11b | 65.11 ± 2.6b | 110.81 ± 4.68b | 0.41 ± 0.00ab | 0.14 ± 0.00bc | 0.62 ± 0.02c | 0.91 ± 0.00b | 0.33 ± 0.01b | 1.10 ± 0.04c | 0.51 ± 0.01c |
| T3 | 101.00 ± 4.06a | 11.32 ± 0.61 | 52.80 ± 5.93 | 2.65 ± 0.29 | 54.49 ± 0.73a | 75.79 ± 0.66a | 130.08 ± 0.71a | 0.42 ± 0.00a | 0.14 ± 0.00c | 0.56 ± 0.03d | 0.91 ± 0.02b | 0.34 ± 0.00b | 1.12 ± 0.02bc | 0.52 ± 0.01bc |
| T4 | 95.40 ± 4.34a | 11.64 ± 0.41 | 53.20 ± 5.81 | 2.60 ± 0.66 | 55.36 ± 0.86a | 77.19 ± 1.12a | 132.33 ± 1.34a | 0.42 ± 0.01a | 0.16 ± 0.00a | 0.73 ± 0.01a | 0.96 ± 0.01a | 0.37 ± 0.01a | 1.20 ± 0.01a | 0.54 ± 0.00a |
| T5 | 96.80 ± 4.92a | 10.56 ± 0.89 | 48.40 ± 5.55 | 2.10 ± 0.37 | 43.49 ± 1.63bc | 66.29 ± 3.61b | 110.21 ± 4.88b | 0.40 ± 0.01b | 0.14 ± 0.00b | 0.66 ± 0.03b | 0.95 ± 0.02a | 0.33 ± 0.01b | 1.12 ± 0.02bc | 0.51 ± 0.00c |
| T6 | 100.20 ± 4.5a | 10.60 ± 0.76 | 49.00 ± 8.15 | 2.47 ± 0.75 | 54.85 ± 0.76a | 75.73 ± 0.30a | 130.40 ± 1.06a | 0.42 ± 0.00a | 0.14 ± 0.00b | 0.61 ± 0.01c | 0.94 ± 0.01aa | 0.36 ± 0.00a | 1.15 ± 0.03ab | 0.53 ± 0.00ab |
Positive effects were observed with observable variations in plant height among the treatment groups. Compared with those in other treatments, the shoot heights in T1 and T7 were the lowest. On the other hand, plants from treatment T4 were the tallest, while the other plants were closely related but differed significantly. Parallel trends were observed in the length of the roots of the plants among the treatments. Similarly, bacterial inoculation significantly affected the macronutrient content, i.e., NPK, in the present study. Substantial differences were recorded across the various treatment groups; in particular, treatments T4, T5, T6, and T8 exhibited the significantly highest levels of these nutrients within wheat straw, with T4 having the highest and T1 having the lowest NPK levels.
The treatments studied under controlled pot conditions demonstrated a significantly positive Pearson correlation coefficient (r), as depicted in Figure 3, when assessed in relation to the physiological and biometric parameters examined in this study. However, with the exception of the relative water content and membrane thermal stability, which demonstrated a slight inclination toward neutrality, the observed associations were consistently positive. This was also apparent in the clustered heatmap [Figure 4], which was based on similarities in correlation coefficients and displayed against growth parameters (horizontally) and treatment (vertically). Closely comparable treatments, such as T6 and T4, were grouped together. Similarly, growth characteristics with greater correlations were clustered together. As a consequence of S. colletis strains with RDFs or 50% RDFs, the locations of various growth metrics and macronutrient (NPK) concentrations in straw were represented by the four zones of the PCA biplot [Figure 5] according to the principal component analysis. Two main components (PC1, 85.55%; PC2, 9.21%) comprised the PCA, which explained 94.76% of the variance under potting conditions [Figure 5]. T4 had a significant effect on the N, P, and K contents of the wheat straw, as indicated by the upper right biplot. On the other hand, T8, T6, and T2 had an impact on the biometric parameters if the crop was represented by the lower right corner of the biplot, where PC1 had positive loading and PC2 had negative loading.
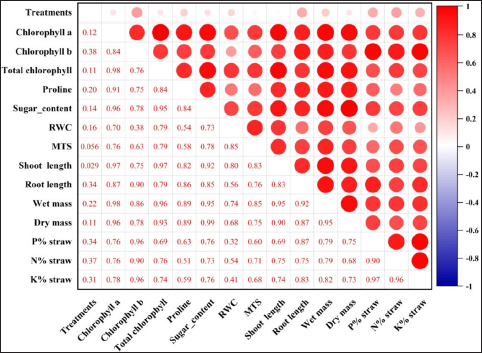 | Figure 3: Correlation coefficient matrix illustrating the impact of Siccibacter colletis on wheat straw nutritional content, physiological characteristics, and biometric measurements in pots with different fertilizer levels. In the matrix, the correlation coefficient (r) values are indicated by the dark red circles and are significantly positive at p < 0.01.
[Click here to view] |
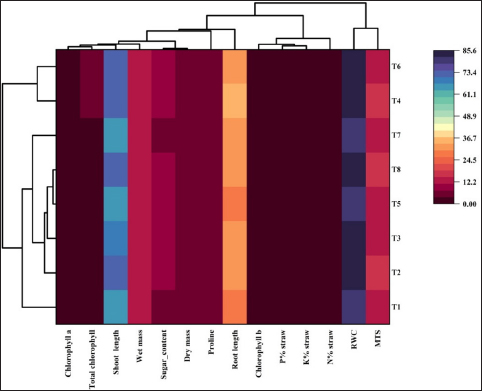 | Figure 4: Heatmap demonstrating that Siccibacter colletis interacts with various growth parameters and the nutritional content of wheat straw at different fertilizer levels in pots. *The color codes (lowest to highest: violet to yellow and blue) directly correlate to the values of the correlation coefficient (r).
[Click here to view] |
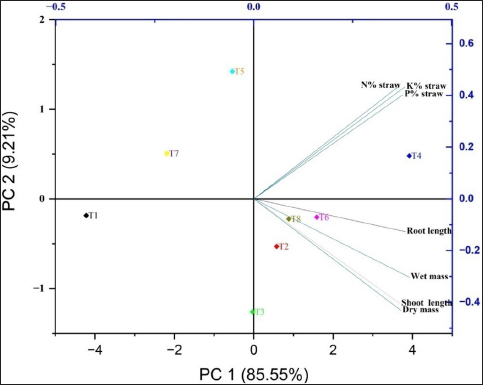 | Figure 5: The 2D biplot depicts the variables of wheat crop characteristics that are grouped according to their PC scores (PC1 and PC2) that are obtained from biometric data, nutrient content in straw, and treatment levels under pot conditions.
[Click here to view] |
3.5. Effect of S. colletis Inoculation in Field Trials
Like in the pot experiments, we studied the effects of bacterial inoculation in conjunction with RDF or a reduced dose of 50% RDF under field conditions. The results revealed a significant increase in several biometric growth indices, including dry biomass output, plant height, and yield-related attributes. Furthermore, the investigation indicated a considerable increase in the macronutrient content in grain and straw. When comparing the treatment groups to T1, i.e., the absolute control, a significant difference in physiological parameters, was detected. Among all the treatments, T6, which had 50% RDF and bacterial inoculum, and T4, which had RDF and bacterial inoculum, exhibited the significantly highest levels of total chlorophyll. In terms of proline and sugar concentrations, there were no significant differences found between treatments T3, T4, and T6; however, they were significantly higher compared to other treatments. There were no statistically significant differences in the relative water content measured in any treatment group (P > 0.05). In contrast to the results of pot studies, the membrane thermal stability data revealed significant variation across the treatment groups. The plants from T3 had the highest MTS, whereas the MTS values from T2, T4, and T6 were statistically identical. Except for the number of spikelets per plant, ear height, and weight, all the biometric parameters significantly differed among the treatment groups.
Furthermore, T4 exhibited the best yield qualities, such as straw, grain, and biological yields, followed by T6 and T3 [Table 4]. However, the harvest indices were significantly different among the groups but more or less similar throughout the treatment groups. The NPK content in the wheat straw and grain was significantly highest in T4. However, no significant difference was found in P content of the wheat straw in T6, T5, and T2. Similarly, the K contents in the wheat straw were found to be significantly higher in treatments T4, T6, followed by T5, and similar trends were generally recorded for the K content in the grains. Similarly, compared with that in the absolute control, the N content in the straw was significantly different across the treatments but was similar in the grains [Table 4].
Consistent with the findings from the controlled pot experiments and similar to those of the pot studies, Pearson’s correlation coefficient between the treatments and various physiological and biometric parameters revealed an overall significant positive correlation [Figure 6]. However, parallel to the results observed under pot conditions, the RWC and MTS tended to reach neutrality, as did the ear height and harvest index. Similarly, the harvest index showed a lower positive correlation with the nitrogen content (r = 0.086) in the wheat straw.
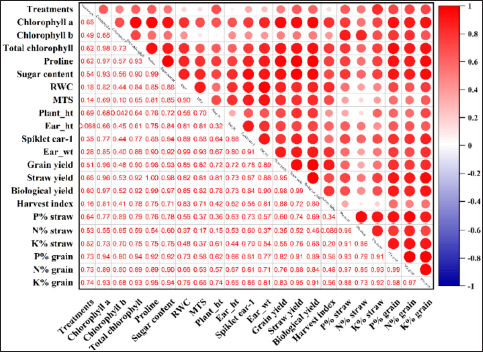 | Figure 6: Correlation coefficient matrix illustrating the impact of Siccibacter colletis wheat straw and grain nutritional content, physiological characteristics, and biometric measurements in the field under different fertilizer levels. In the matrix, the correlation coefficient (r) values are indicated by the dark red circles and are significantly positive at p < 0.01.
[Click here to view] |
A heatmap generated for physiological and biometric parameters (horizontally) against various treatments (vertically) depicted different groupings based on their similarities. In line with the results observed in the controlled pot experiments, treatments containing RDF, either alone or in combination with S. colletis, demonstrated significant positive interactions with respect to yield-related attributes under field conditions [Figure 7], which was reflected by the proximity of T6–T4 to both the inoculum control and the absolute control (i.e., T1 and T2) within the heatmap.
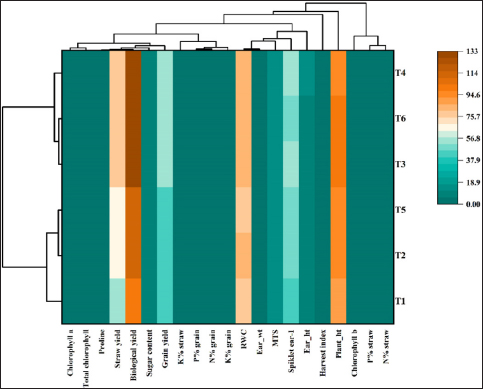 | Figure 7: Heatmap demonstrating the relationships between the physiological, biometric, and mineral contents of wheat straw and Siccibacter colletis plants grown in the field under various treatment conditions. **The color codes (lowest to highest: violet to yellow and blue) directly correlate to the values of the correlation coefficient (r).
[Click here to view] |
PCA revealed two principal components contributing 75.15% (PC1) and 14.01% (PC2) of the variance, collectively accounting for 89.16% of the total variance. It is evident from the biplot that treatment T6, with 50% RDF together with rhizobacterial inoculation, and treatment T3, involving RDF alone, predominantly influenced wheat yield attributes, as shown in the upper right quartile, having a large positive loading for both principal components [Figure 8]. The nutritional content of both the grain and straw was predominantly affected by treatment T4, as demonstrated by a positive loading on PC1 and a negative loading on PC2, placing it in the lower right quartile of the biplot, in contrast to the results from the pot trials.
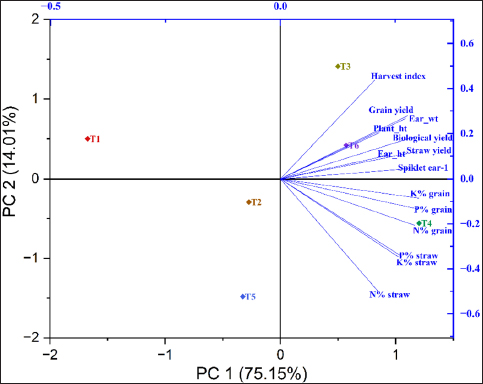 | Figure 8: The 2D biplot depicts the variables of wheat crop characteristics that are grouped on PC scores (PC1 and PC2) that are obtained from mineral content in grain and straw and biometric parameters along with treatment levels in the field.
[Click here to view] |
4. DISCUSSION
PGPR, by virtue of its ability to provide or mobilize different nutrients and secrete other metabolites, sustains plant growth with the minimum amount of chemical fertilizers and alleviates many stresses such as salinity, high temperature, and pests. P and K being the major nutrients required by the plants are to be supplied externally in the form of chemical fertilizers frequently. However, the unfavorable soil pH renders P unavailable for plants due to the formation of insoluble complexes with aluminum or iron minerals. Similarly, K is also not present in available form in the soil for the plants. Furthermore, various abiotic stresses such as high temperature, drought, and salinity also reduce plant growth, especially in semiarid regions such as Mahendergarh, India. P- and K-solubilizing microorganisms have emerged as potential candidates for sustainable supply of these minerals to plants. Besides providing P and K, many microorganisms have the potential to alleviate or reduce the effects of various abiotic stresses in such areas. However, these microorganisms are adapted to certain soil conditions and fail to provide desirable effects under unfavorable soil conditions. So the researchers are working on different approaches to ensure the effectiveness of P- and K-solubilizing bacteria in sustaining crop yields in diverse agricultural environments. This involves several methodologies and approaches, such as analyzing the area-specific soil characteristics (soil texture, pH, and nutrient content), applying high-throughput methods for isolating and identifying area-specific indigenous microbes, molecular techniques for strain development, compatible testing with agrochemical and other beneficial microbes, and conducting several seasonal field trials to prepare area-specific bioinoculant formulations for sustaining crop yields in such areas. A large number of bacteria belonging to Enterobacteriacea, including Enterobacter cloacae, Enterobacter sp. CM94, Klebsiella michiganensis TS8, and Lelliottia jeotgali MR [30-32], are reported to be efficient PGPR and have been applied in the field for improving crop yields. The most efficient bacterial isolate in the present study was identified as S. colletis SSRP30 (acc. no. OR150488), based on the 16S rRNA gene sequence they are primarily present in the rhizosphere soil and as mutualistic endophytes [33,34],. Jackson et al. [33] published the first description of S. colletis as a unique species isolated from tea leaves and poppy seeds. Both Siccibacter and Siccibacter-derived genera have been reported for their various plant growth-promoting characteristics [35]. According to Chamkhi, et al. [34], S. colletis from alfalfa rhizosphere has an extensive capacity to colonize plant roots and shield plants from biotic and abiotic stresses. Similarly, Salazar-RamÍRez et al[36]. isolated S. colletis from the candelilla (Euphorbia antisyphilitica) rhizosphere. This is the first study in which S. colletis was reported from the wheat rhizosphere soil of Aravalli foothills. The foothill soils are rich in minerals, and hence the probability of finding mineral-weathering microorganisms is always high. The results of the in vitro experiment unequivocally demonstrated that the S. colletis SSRP30 strain, which was chosen for this study, possesses many PGP features, including IAA, ACC deaminase, siderophore production, HCN production, and biofilm formation, all of which directly or indirectly aid in plant growth and development.
The competitive saprophytic ability of the microorganisms varies and determines their fate when applied in the field. Isolates that showed promising results in the in vitro experiments failed to perform under field conditions because of several environmental and edaphic factors, which vary from soil to soil. Hence, it is imperative to evaluate the competitive saprophytic ability of isolates by means of pot and field experiments and analyze their effect on yield. The culture was able to provide significant yield benefits compared to the control, which indicated the superior competitive saprophytic ability of the isolate. Since under unfavorable soil pH conditions, the P and K form complexes with different minerals, making them unavailable for plants, the applied isolates were instrumental in the sustainable release of these macronutrients, which resulted in better biometric parameters and crop yield.
The effects of bacterial inoculation were investigated in both the field and pot. Wheat, being the staple crop in Northwest India, is being grown in stressed soil affected by salinity and water stress. The macro- and micronutrient availability in rhizospheric soil can be significantly increased by applying microbial inoculants [37,38]. In the soil ecosystem, microorganisms employ several mechanisms, including acidolysis, production of organic acids, which lower the pH of the surroundings to mineralize/solubilize mineral nutrients, and transform them into forms that may be used by plants [39,40]. Our findings are in line with this fact, S. colletis SSRP30 lowered the pH in Pikovskaya’s and Aleksandrov’s broth to release the bound mineral in the broth, thus improving the NPK uptake in wheat crops. Similar reports were also presented for various strains of Siccibacter sp. having the capacity to solubilize P and K. Furthermore, growth hormones such as IAA and ACC deaminase are crucial plant hormones that increase plant biomass and play a pivotal role in exerting tolerance toward various environmental conditions. Extracellular polysaccharides produced by bacterial inoculants can assist in nutrition intake by holding nutrients and energy substrates, which also provide resistance to water and temperature stress, complemented by ACC deaminase [34,35,41-45]. S. colletis SSRP30 enhanced the total biomass of wheat crops in both pot and field studies, reflected through improved nutrient uptake and phytohormone production and increased chlorophyll and sugar content in plants [46-48].
With respect to wheat plants supplemented with RDF, 50% RDF, and without chemical fertilizer under net house conditions, S. colletis SSRP30 significantly enhanced height, wet and dry mass, yield and other yield-associated traits, grain yield, straw yield, biological yield, and the harvest index in the wheat crop. Compared with those in plants supplied with RDF, 50% lower doses of RDF, and without chemical fertilizer, the values in RDFs inoculated with bacterial inoculum plants were significantly greater, followed by those in RDFs without inoculum, 50% lower doses of RDF, and no chemical fertilizer with inoculum. This may be due to the fact that plants do not use all the nutrients applied through chemical fertilizers. A significant amount gets bound to other components of soil, reducing its availability and requiring extra energy to extract and use the same [49]. When bioinoculants such as S. colletis SSRP30 are applied to the field, they aid the plants in nutrient acquisition through the mechanisms discussed above. The lowest values were recorded for the absolute control without the inoculant. Plant growth and development in pot and field experiments are significantly enhanced by the application of PGPR [46,50-53].
Among the different treatments, the highest NPK content was recorded in the grain and straw of the plants inoculated with the bacterial inoculum and RDF compared to that in the other treatments. These findings are consistent with earlier research showing that PGPR inoculation of plants increased the amount of NPKs (macronutrients) in wheat straw and grain [54-56].
Based on the findings presented in this study as well as the findings of earlier research, S. colletis can be a potential candidate for in vivo application as a bioinoculant with or without combinations of insoluble P and K. Thus, S. colletis SSRP30 can improve plant growth and increase yield by mobilizing unavailable P and K existing in the soil and increasing plant uptake in fields.
REFERENCES
1. Food and Agriculture Organization of the United Nations. Food and Agriculture Organization Corporate Statistical Database, 2024 [cited 2024 March 9]. Rome: Food and Agriculture Organization of the United Nations (FAO). Available from: www.fao.org/faostat/en/#data/QCL
2. Udhayan N, Naik AD, Hiremath GM. An economic analysis of wheat cultivation in north-Karnataka, India. Int J Plant Soil Sci. 2023;35(20):939-45. [CrossRef]
3. Pérez Vázquez A, Leyva Trinidad DA, Gómez Merino FC. Challenges and proposals to achieve food security by the year 2050. Rev Mex De Cienc Agric. 2018;9(1):175-89. [CrossRef]
4. Kumar P, Singh S, Pranaw K, Kumar S, Singh B, Poria V. Bioinoculants as mitigators of multiple stresses: a ray of hope for agriculture in the darkness of climate change. Heliyon. 2022;8(11):e11269. [CrossRef]
5. Mahadevamurthy M, Channappa TM, Sidappa M, Raghupathi MS, Nagaraj AK. Isolation of phosphate solubilizing fungi from rhizosphere soil and its effect on seed growth parameters of different crop plants. J Appl Biol Biotechnol. 2016;4(6):22-6. [CrossRef]
6. Patil S, Nikam M, Anokhina T, Kochetkov V, Chaudhari A. Multi-stress tolerant plant growth promoting Pseudomonas spp. MCC 3145 producing cytostatic and fungicidal pigment. Biocatal Agric Biotechnol. 2017;10:53-63. [CrossRef]
7. Orozco-Mosqueda MDC, Glick BR, Santoyo G. Acc deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiol Res. 2020;235:126439. [CrossRef]
8. Yadav AN. Phosphate-solubilizing microorganisms for agricultural sustainability. J Appl Biol Biotechnol. 2022;10(3):1-6. [CrossRef]
9. Pikovskaya R. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiologia. 1948;17:362-70.
10. Aleksandrov V. Organo-mineral fertilizers and silicate bacteria. Dokl Akad Nauk. 1958;7:43-8.
11. Premono ME, Moawad A, Vlek P. Effect of phosphate solubilizing Pseudomonas putida on the growth of maize and its survival in the rhizosphere. Indones J Agric Sci. 1996;11(1):13-23.
12. Holiday ER, Preedy JR. The precision of a direct-reading flame photometer for the determination of sodium and potassium in biological fluids. Biochem J. 1953;55(2):214-20. [CrossRef]
13. Mehta S, Nautiyal CS. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr Microbiol. 2001;43(1):51-6. [CrossRef]
14. Gordon S, Paleg LG. Quantitative measurement of indole acetic acid. Physiol Plant. 1957;10(1):37-48. [CrossRef]
15. Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160(1):47-56. [CrossRef]
16. Jensen H. Notes on the biology of Azotobacter. J Appl Microbiol. 1951;14(1):89-94. [CrossRef]
17. Cappuccino JG, Sherman N. Microbiology: a laboratory manual. 10th ed. San Francisco (CA): Pearson; 1999.
18. Saleh SS, Glick BR. Involvement of gacS and rpoS in enhancement of the plant growth-promoting capabilities of Enterobacter cloacae CAL2 and UW4. Can J Microbiol. 2001;47(8):698-705. [CrossRef]
19. Walkley A, Black IA. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934;37(1):29-38. [CrossRef]
20. Subbiah BV, Asija GL. A rapid procedure for the determination of available nitrogen in soils. J Curr Sci. 1956;25:259-60.
21. Olsen SR, Cole CV, Watanabe FS. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Washington (DC): US Government Printing Office; 1954.
22. Jackson ML. Soil chemical analysis. New Delhi: Prentice Hall lnc; 1973.
23. Barnes JD, Balaguer L, Manrique E, Elvira S, Davison AW. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ Exp Bot. 1992;32(32):85-100. [CrossRef]
24. DuBois M, Gilles KA, Hamilton JK, Rebers Pt, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350-6. [CrossRef]
25. Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water-stress studies. Plant Soil. 1973;39(1):205-7. [CrossRef]
26. Ibrahim AM, Quick JS. Genetic control of high temperature tolerance in wheat as measured by membrane thermal stability. Crop Sci. 2001;41(5):1405-7. [CrossRef]
27. Lindner RC. Rapid analytical methods for some of the more common inorganic constituents of plant tissues. Plant Physiol. 1944;19(1):76-89. [CrossRef]
28. Koenig RA, Johnson CR. Colorimetric determination of phosphorus in biological materials. Ind Eng Chem Anal Ed. 1942;14(2):155-6. [CrossRef]
29. Pickett EE, Koirtyohann SR. Emission flame photometry-a new look at an old method. J Anal Chem. 1969;41(14):28-42. [CrossRef]
30. Panigrahi S, Mohanty S, Rath CC. Characterization of endophytic bacteria Enterobacter cloacae MG00145 isolated from Ocimum sanctum with indole acetic acid (iaa) production and plant growth promoting capabilities against selected crops. S Afr J Bot. 2020;134:17-26. [CrossRef]
31. Sharma A, Chakdar H, Vaishnav A, Srivastava AK, Khan N, Bansal YK, et al. Multifarious plant growth-promoting rhizobacterium Enterobacter sp. CM94-mediated systemic tolerance and growth promotion of chickpea (Cicer arietinum L.) under salinity stress. Front Biosci (Landmark Ed). 2023;28(10):241. [CrossRef]
32. Chen L, Bai Y, Liu S, Liu H, Chen R, Xiao Y. Effects of plant growth-promoting rhizobacteria Klebsiella michiganensis TS8 and Lelliottia jeotgali MR2 on the growth and cadmium uptake of Arabidopsis thaliana under cadmium stress. Sheng Wu Gong Cheng Xue Bao. 2022;38(5):1915-28. [CrossRef]
33. Jackson EE, Masood N, Ibrahim K, Urvoy N, Hariri S, Forsythe SJ. Description of Siccibacter colletis sp. nov., a novel species isolated from plant material, and emended description of Siccibacter turicensis. Int J Syst Evol Microbiol. 2015;65(4):1335-41. [CrossRef]
34. Chamkhi I, Zwanzig J, Ibnyasser A, Cheto S, Geistlinger J, Saidi R, et al. Siccibacter colletis as a member of the plant growth-promoting rhizobacteria consortium to improve faba-bean growth and alleviate phosphorus deficiency stress. Front Sustain Food Syst. 2023;7:1134809. [CrossRef]
35. Mamarasulov B, Davranov K, Jahan MS, Jabborova D, Nasif O, Ansari MJ, et al. Characterization, enzymatic and biochemical properties of endophytic bacterial strains of the medicinal plant Ajuga turkestanica (Rgl.) Brig (lamiaceae). J King Saud Univ Sci. 2022;34(6):102183. [CrossRef]
36. Salazar-RamÍRez M, Sáenz-Mata J, Preciado-Rangel P, Fortis-Hernández M, Rueda-Puente E, Yescas-Coronado P, et al. Plant growth-promoting rhizobacteria associated to candelilla rhizosphere (Euphorbia antisyphilitica) and its effects on Arabidopsis thaliana seedlings. Not Bot Horti Agrobot Cluj Napoca. 2021;49(2):12294. [CrossRef]
37. Belimov AA, Shaposhnikov AI, Syrova DS, Kichko AA, Guro PV, Yuzikhin OS, et al. The role of symbiotic microorganisms, nutrient uptake and rhizosphere bacterial community in response of pea (Pisum sativum L.) genotypes to elevated al concentrations in soil. Plants. 2020;9(12):1801. [CrossRef]
38. Dobrin A, Zugravu MM, Mot A, Musat M, Burnichi F, Ciceoi R. The influence of microbial inoculants on micro-and macronutrients on calcaric alluvial soil. Rom Agric Res. 2021;38:487-94. [CrossRef]
39. Elhaissoufi W, Ghoulam C, Barakat A, Zeroual Y, Bargaz A. Phosphate bacterial solubilization: a key rhizosphere driving force enabling higher P use efficiency and crop productivity. J Adv Res. 2022;38:13-28. [CrossRef]
40. Ullah S, Bano A, Ullah A, Shahid MA, Khan NJR. A comparative study of plant growth promoting rhizobacteria (PGPR) and sowing methods on nutrient availability in wheat and rhizosphere soil under salinity stress. Rhizosphere. 2022;23:100571. [CrossRef]
41. Sayahi N, Djemal R, Ben Merdes K, Saidii MN, Yengui M, Gdoura R, et al. Characterization of Siccibacter sp. Strain C2 a novel rhizobacterium that enhances tolerance of barley to salt stress. Curr Microbiol. 2022;79(8):239. [CrossRef]
42. Chen Z, Guo Z, Zhou L, Xu H, Liu C, Yan X. Advances in identifying the mechanisms by which microorganisms improve barley salt tolerance. Life. 2024;14(1):6. [CrossRef]
43. Martynenko E, Arkhipova T, Safronova V, Seldimirova O, Galin I, Akhtyamova Z, et al. Effects of phytohormone-producing rhizobacteria on casparian band formation, ion homeostasis and salt tolerance of durum wheat. Biomolecules. 2022;12(2):230. [CrossRef]
44. Backer R, Rokem JS, Ilangumaran G, Lamont J, Praslickova D, Ricci E, et al. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front Plant Sci. 2018;9:1473. [CrossRef]
45. Vejan P, Abdullah R, Khadiran T, Ismail S, Nasrulhaq BA. Role of plant growth promoting rhizobacteria in agricultural sustainability—a review. Molecules. 2016;21(5):573. [CrossRef].
46. Pereira SIA, Abreu D, Moreira H, Vega A, Castro PML. Plant growth-promoting rhizobacteria (PGPR) improve the growth and nutrient use efficiency in maize (Zea mays L.) under water deficit conditions. Heliyon. 2020;6(10):e05106. [CrossRef]
47. Urana R, Singh N, Sharma P. Effects of PGPR on growth and photosynthetic pigment of Trigonella foenum-graceum and Brassica juncea in PAH-contaminated soil. SN Appl Sci. 2019;1(7):761. [CrossRef]
48. Lalay G, Ullah S, Ahmed I. Physiological and biochemical responses of Brassica napus L. to drought-induced stress by the application of biochar and plant growth promoting rhizobacteria. Microsc Res Tech. 2022;85(4):1267-81. [CrossRef]
49. Abebe TG, Tamtam MR, Abebe AA, Abtemariam KA, Shigut TG, Dejen YA, et al. Growing use and impacts of chemical fertilizers and assessing alternative organic fertilizer sources in Ethiopia. Appl Env Soil Sci. 2022:4738416. [CrossRef]
50. Fan X, Zhang S, Mo X, Li Y, Fu Y, Liu Z. Effects of plant growth-promoting rhizobacteria and N source on plant growth and N and P uptake by tomato grown on calcareous soils. Pedosphere. 2017;27(6):1027-36. [CrossRef]
51. Dal Cortivo C, Barion G, Visioli G, Mattarozzi M, Mosca G, Vamerali T. Increased root growth and nitrogen accumulation in common wheat following PGPR inoculation: assessment of plant-microbe interactions by ESEM. Agric Ecosyst Environ. 2017;247:396-408. [CrossRef]
52. Li H, Qiu Y, Yao T, Ma Y, Zhang H, Yang X. Effects of PGPR microbial inoculants on the growth and soil properties of Avena sativa, Medicago sativa, and Cucumis sativus seedlings. Soil Tillage Res. 2020;199:104577. [CrossRef]
53. Poria V, Debiec-Andrzejewska K, Fiodor A, Lyzohub M, Ajijah N, Singh S, et al. Plant growth-promoting bacteria (PGPB) integrated phytotechnology: a sustainable approach for remediation of marginal lands. Front Plant Sci. 2022;13:999866. [CrossRef]
54. Rana A, Joshi M, Prasanna R, Shivay YS, Nain L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur J Soil Biol. 2012;50:118-26. [CrossRef]
55. Yadav RC, Sharma SK, Varma A, Singh UB, Kumar A, Bhupenchandra I, et al. Zinc-solubilizing Bacillus spp. In conjunction with chemical fertilizers enhance growth, yield, nutrient content, and zinc biofortification in wheat crop. Front Microbiol. 2023;14:1210938. [CrossRef]
56. Kumar A, Maurya BR, Raghuwanshi R. The microbial consortium of indigenous rhizobacteria improving plant health, yield and nutrient content in wheat (T. aestivum). J Plant Nutr. 2021;44(13):1942-56. [CrossRef]

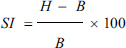 where H is the diameter of the halo zone around the colony and B is the diameter of the bacterial colony.
where H is the diameter of the halo zone around the colony and B is the diameter of the bacterial colony.