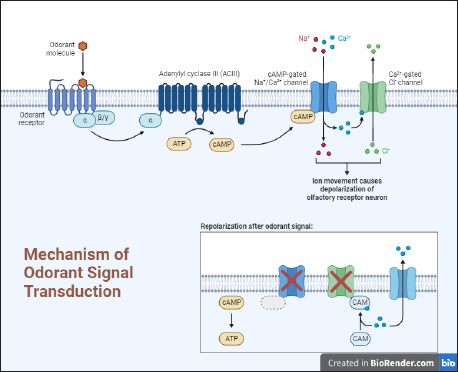
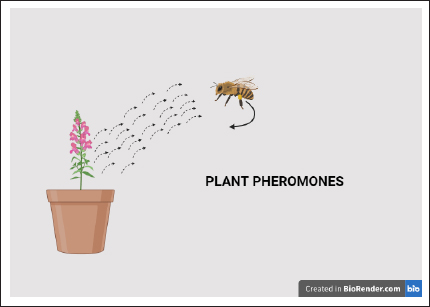
1. INTRODUCTION TO PHEROMONES
Chemical cues, called pheromones, are exchanged by individuals of the same species. By attracting one or more adult individuals to the pheromone-producing individual, several of the known insect pheromones function as long-range attractants. In order to attract members of the opposing sex for the express goal of mating, individuals of one sex produce sex pheromones. In general, one sex releases aggregation pheromones that attract conspecific male and female individuals to areas where mating takes place and where females choose sites for oviposition on suitable substrates for larval feeding. Pheromones thus function in unique biological situations and are very species-specific. Pheromones are distinct from other, less specialized insect attractants, such as the aromas of food or host plants, which may be all-purpose attractants for several species. Improvements in the collection, isolation, and chemical identification of extremely minute amounts of pheromones generated by live insects have sparked a boom in research on insect pheromones during the last 30 years. For the majority of the main stored-product insect pests, i.e., a variety of insects that infest and damage stored food, books, documents, fabrics, leather, carpets, and any other dried or preserved items, at the current time, attractant pheromones have been discovered and synthesized. Large populations of certain pest species may be attracted to traps using artificial pheromones. Pheromones have been used to manipulate and suppress populations in the study. However, pheromones are not commonly utilized to control insect pest populations at the moment. Synthetic pheromones are often utilized in traps as monitoring and detection techniques in pest control programs. Additionally, traps baited with non-pheromone food attractants, as well as unbaited traps that collect insects passively, are employed to monitor insect populations in settings where stored goods are present. In addition to discussing the use of pheromones and associated trapping and enticing techniques as Integrated Pest Management (IPM) tools, this chapter will also consider the possibility of creating pheromone-based strategies for pest population control. In 1966, the black carpet beetle, Attagenus unicolor (Brahm) (megatoma), chemically produced the first stored-product insect pheromone. Since then, pheromones from more than 40 species of insects that feed on stored goods have been found. Numerous review publications have thoroughly examined the biology, chemistry, and use of insect pheromones found in stored products [1].
Pheromones are classified according to their chemical makeup, purpose, and the environment in which they are employed by a species. The classification is given in the table below [Table 1].
Table 1: Types of pheromones and functions.
| Types of pheromones | Functions | References |
|---|
| Releasers | These cause a prompt, precise response. They are short-acting and are often linked to sexual attraction. | [65] |
| Signalers | These release information about the animal. | [7] |
| Modulators | These are found in sweat and impact emotions. They influence emotions and moods. | [66] |
| Primers | These impact endocrine or neuroendocrine responses connected to development and reproduction. | [24] |
2. TYPES OF PHEROMONES
2.1. Sex Pheromones
Sex pheromones are chemical signals that one member of a species emits, and another member of the same species detects. They provide a number of functions, including mating attraction, territorial marking, and communication. The ability of creatures to recognize potential mates of the other gender is the primary purpose of sex pheromones. It goes without saying that sex-specific chemical signals indicating gender are often created, whether they are male- or female-specific or a mix of the two. Many of the most well-known insect sex pheromones are female-specific, long-range male attractants, as a result of the finding of bombykol. Studies of several Hemiptera (true bugs) have mostly identified male-specific sex pheromones, which may represent historical prejudice rather than taxonomic class bias [Figure 1] [2]. Pheromones come in two different varieties: releasers and primers. While primer pheromones generate physiological changes in an animal that eventually lead to a behavioral response, releaser pheromones elicit rapid behavioral reactions in insects upon receiving them.
There are three main categories of chemically recognized releaser pheromones: those that promote recruitment, alarm behavior, and sexual attraction. Recruiting pheromones are mostly used to indicate routes to food supplies. Bees and other flying insects apply the compounds at irregular intervals, while terrestrial insects leave continuous odor trails. For warning pheromones, insects exhibit far reduced sensitivity and chemo specificity. Alarm selectivity is more influenced by volatility than by distinctive structural characteristics. The full sexual toolkit is released by sex pheromones. So, an inanimate item with a sex pheromone on it may attract and seek to mate with a male insect. It seems that the majority of insects are very choosy and sensitive to their species’ sex pheromone.
The queen bee uses a sophisticated pheromone system to regulate worker behavior. The fatty acid 9-ketodecenoic acid, which is generated by the queen and transferred to the workers, is one well-known part of this system. This substance hinders the workers’ ability to raise queens and stops the development of ovaries in the workers. Additionally, virgin queen bees employ the same substance as a sex attractant. [3]. Insect pheromones, which are volatile in nature, make up the bulk of pheromones that have been discovered so far. It has been exceedingly uncommon to identify nonvolatile pheromones, particularly in vertebrates. The sexual behavior of male and female garter snakes is mediated by pheromones. The male sex identification pheromone includes squalene, but the female sex attractiveness pheromone of the Canadian red-sided garter snake, Thamnophis sirtalis parietalis, has a unique sequence of nonvolatile saturated and monounsaturated long-chain methyl ketones. Field testing has shown that these isolated, recognized, and partly synthesized chemicals are physiologically active [4].Even among closely related species, the Lepidoptera use a wide variety of pheromone structures. The puzzle is how signal divergence happens. It is unlikely that transitions to pheromones with various structures occur via adaptive modifications in modest increments because of the significant normalizing selection pressure on blend composition and response preferences. Here, we show evidence in favor of the theory that activation of a desaturase gene transcript found in the pheromone gland led to a significant change in the pheromone of an Ostrinia species. We also show that there are uncommon men who react to the novel pheromone mixture. They would make it possible to follow male reactions asymmetrically to the new mix, leading to the emergence of an Ostrinia species with structurally distinct sex pheromone components [5].
In regards to the presence of pheromonal communication, data on humans has caused the most debate. The likelihood of pheromonal communication in humans is evaluated in this review with a discussion of chemical substances produced by the axilla that may serve as pheromones, the likelihood that the vomeronasal organ (VNO), a putative pheromone receptor organ in many other mammals, is functional in humans, and the potential functions of pheromones in humans. The human axilla resembles other primates’ smell glands due to interactions between the axillary secretions and cutaneous microorganisms. Axillary secretions in humans have a chemistry that is similar to other mammalian pheromone systems, and they also have similar effects on conspecifics. The receptor is another unidentified element of whatever chemical molecules have pheromonal effects on people. There is abundant evidence to support the idea that the olfactory epithelium may react to pheromones, despite the fact that the VNO has been linked to the receipt of pheromones in many vertebrates. The VNO is only one avenue by which this information might reach the central nervous system. Additionally, a chemical need not be a pheromone just because it causes the VNO’s receptors to light up. The fact that crucial elements usually present in the functional VNO of other nonprimate animals are absent in humans is a significant caveat, indicating that the human VNO may not work as other mammals’ VNOs have been reported to. Pheromones are categorized as releasers, modulators, signalers, and primers from a larger viewpoint. The first three can definitely be found in humans, according to strong evidence. Examples include influences on the menstrual cycle (primer effects), the mother’s ability to recognize her infant by scent (signaler), and the possibility that people may emanate distinct odors depending on their emotions (suggestive of modulator effects). In mature humans, releaser effects are not supported by solid data. It is noteworthy that no bioassay-guided research project has succeeded in isolating genuine human pheromones, a step that would clarify the precise roles of human chemical signals [6].
2.2. Alarm Pheromones
Different creatures release alarm pheromones as a chemical warning in the presence of danger or threat. Conspecifics are forewarned by these signals, prompting an immediate and well-coordinated defensive reaction. Alarm pheromones have been well explored in several species and are important in a variety of ecological and evolutionary circumstances. Alarm pheromones cause a variety of behaviors to be triggered in reaction to a perceived threat. They may trigger defensive responses in social insects, such as the mobilization of the colony to fight or repel predators. Alarm pheromones in fish may cause predator-avoidance actions like running away or looking for cover. These reactions are often well-coordinated and crucial to the group’s survival [7].Alarm pheromones aren’t only used by prey species. Alarm cues may also be used to a predator’s advantage. For instance, certain predatory insects, such as ladybirds, are known to find their prey by making use of the alarm signals of aphids. Due to these complex ecological interactions, it is now possible for different species to respond differently to the same chemical stimuli. Academic research on alarm pheromones is one of many fields of study. Understanding these chemical signals may help with pest control and agriculture in real-world situations. Researchers and farmers may create ways to interfere with the communication of nuisance insects by understanding their alarm pheromones, possibly minimizing crop damage.
In other situations, specific pheromones may also be traded. The release of a biological substance by a member of any species to warn conspecifics of potential danger is one example of such an occurrence. Alarm pheromones (APs) are the biochemical signals that are emitted during this activity, which often occurs when an organism is in danger or under attack from a predator or its environment [8]. Many animals employ alarm signals to warn conspecifics, although their methods vary. Some animals utilize auditory warning calls, others visual or chemosensory cues, or a mix of both [9].
2.3. Aggregation Pheromones
Various species use aggregation pheromones to recruit conspecifics and establish groups. Insects, animals, and certain plants use these chemicals to coordinate social behavior and interactions. Insect aggregation pheromones are well studied. Bark beetles use aggregation pheromones to settle on host trees to mate and deposit eggs. VOCs in these pheromones are sensed by conspecifics from afar. Bark beetle aggregation pheromones are complex chemical mixtures that attract and govern behavior. These pheromones frequently include numerous components that attract and coordinate insects, according to studies.
Aggregation pheromones are perceived via several sensory modalities. Olfaction is the main way insects perceive chemical stimuli. Insect antennae and appendages have specialized sensors to detect pheromone trails. In rats, the vomeronasal organ in the nasal cavity detects aggregation pheromones. Neural responses from these pheromone receptors affect behavior, such as mating or foraging [10]. Not just animals use aggregation pheromones. Certain plants produce volatile chemicals to attract herbivores that eat their tissues. When attacked by herbivorous insects, maize plants release volatile chemical substances. Herbivore enemy parasitic wasps use these chemicals as aggregation pheromones. Chemical signals attract wasps, which hunt on herbivorous insects and defend maize. The ecological and evolutionary effects of aggregation pheromones must be understood. Aggregation pheromones impact species distribution and density in ecosystems. Social insects like ants emit aggregation pheromones to create forage and defense colonies. This aggregation may cause ecological processes to cascade.
The evolution and use of aggregation pheromones may affect species’ fitness and survival. Complex social systems evolve as those who can react to aggregation pheromones profit from group life and collaboration. Plants may evolve aggregation pheromones to guard against herbivores, which can affect herbivore evolution and drive chemical cue and response mechanism development [11]. Aggregation is much harder. It shows the aggregation and even permanence of people at a given position at the endpoint level. Although aggregation may be the result of attraction and arrestment or, in certain situations, inadvertent arrival and arrestment, it does not immediately suggest a navigational system. Thus, odor-induced aggregation may be characterized by an endpoint distribution. The phrase “aggregation pheromone” refers to a variety of behaviors with different orienting mechanisms, behavioral and ecological purposes, and evolutionary factors. One way to distinguish aggregation pheromones from typical sex-attractants is to limit them to circumstances when both sexes react, or when the responder’s sex is the emitter’s.
Some bark beetles (Scolytinae) use aggregation pheromones to attack the victim in bulk. One sex generates a pheromone that attracts the other for mating and attracts both sexes of beetles. The collective onslaught weakens the tree, increasing the beetle’s chances of survival. Finding a partner precedes host colonization and drives orientation selection [12]. Certain creatures, especially social insects like ants and termites, employ trail pheromones as chemical signals to communicate and coordinate movement within a group. These pheromones act as a trail for other animals to follow, facilitating effective resource exploitation, navigation, and foraging. These pheromones may arise from a single gland or, in some rare situations, from a mixture of two glands. They can be made up of a single compound or, in one extraordinary example, a blend of as many as 14 compounds. They could be unique to a particular species or might be shared by many species. They are found by workers at trace levels on a trail and are present in glandular secretions in nanogram to picogram concentrations [13]. According to definitions, trail-laying is a field activity in which an insect marks a path with smell or odor traces so that other insects in the same group may follow it [14].
Scout ants locate food to start trails. When it returns to the nest after feeding, it leaves chemicals. It attracts additional workers in unknown ways within or outside the nest. It may include antennal touch, regurgitation, jerking, or another odor that gets workers to leave the nest. Recruitment is now recognized as part of the chemical process of trail-following. Recruitment is communication that brings nestmates to work. The hired laborers follow the food trail. Returning food-laden workers strengthens the path. When the food runs out, and the workers return empty, they stop applying trail stuff, and the odor disappears. Thus, the route survives only as long as needed [15]. Social insect societies utilize a variety of tactics for adaptation and optimization, but one prominent one is the application of trail pheromones. Foraging and navigation efficiency are vital to the colony’s survival and reproduction. Many ant and termite species have thrived due to improved trail pheromones and higher sensitivity. Some species use social insect trail pheromones, which is intriguing. Parasitic ants or kleptoparasites may follow host ants to food sources without foraging. These ecosystems have intricated evolutionary interactions and adaptations among species.
2.4. Territorial Pheromones
Territorial pheromones are chemical signals used by a variety of species to create and defend territories. They are an essential component of chemical communication in the animal world. These pheromones have been the focus of in-depth ethological investigation because they play a crucial function in a variety of ecological and evolutionary circumstances [16]. The complex chemical molecules that make up territorial pheromones provide unique information about the identity, sex, and health of the territorial owner. Specialized glands, which are found in varied forms throughout animals, may manufacture these substances. For example, it is known that the sebaceous or urine glands of animals are providers of territorial pheromones. Urine markings are often used by rats to define territorial boundaries. These distinctive smell signals in the markings provide information about the identity of the bearer and its propensity to protect the area [17]. Territorial pheromones have a crucial role in ecology. They control who has access to resources, lessen the likelihood of direct disputes, and support ecological equilibrium. For instance, territorial marking with urine and scent markings helps large cats, who are territorial carnivores, designate their hunting territory and minimize interactions with possible rivals. This reduces potentially fatal confrontations and frees up energy for essential tasks like hunting.
Intraspecific territoriality may affect population dynamics, dispersion patterns, and resource use. Territorial pheromones are often the mediators of intraspecific territoriality. Chemical signals have a crucial role in controlling foraging and resource exploitation in social insects like ants because territories are not only about space but also about resources. Understanding the spatial organization of populations and their interactions in natural environments requires an understanding of territoriality and territorial pheromones [18]. There are several practical uses for knowing territorial pheromones. Researchers have looked at the possible use of artificial territorial pheromones in agriculture to control insect populations or prevent animals from entering certain regions [67-69]. Understanding territorial behavior in animals may help conserve biodiversity by designing protected areas and habitat corridors.
5. PHEROMONES IN HUMANS
5.1. Human Pheromones and Their Existence
Axillary steroids, vaginal aliphatic acids, and stimulators of the vomeronasal organ are the three kinds of potential human pheromones that have received the majority of attention in the search for evidence of these substances. Examples from each of these groups have been patented for use in commerce and, in some instances, actively promoted, although there is insufficient proof to back up any specific claim that a drug behaves as a human pheromone. Although the enormous axillary scent glands in humans are ideally suited for pheromone generation, they may also be employed for non-pheromonal odor communication, such as the exchange of immune system-related information. The presence of human pheromone systems may be suggested by hypothesized menstrual synchronization among social groups of women and hypothesized acceleration of the menstrual cycle brought on by the scents of males, although the evidence in both situations is still ambiguous [34].
Pheromones may be released from the skin into the environment via two different methods. According to one theory, putative pheromones are very volatile. If there are any odorous molecules on the skin, they will volatilize and be inhaled by someone nearby. The idea that relatively non-volatile pheromones found on or connected to the skin surface may naturally be delivered by the skin itself is a more intriguing prospect [35].
5.2. Pheromones and Sexual Attraction
Increasing data suggests that pheromones have a significant impact on human sexual attraction. For instance, research revealed that women exposed to the perspiration of males in optimal physical shape assessed them as more appealing compared to women exposed to the perspiration of men who were not in optimal physical shape. A further study revealed that males who were exposed to the fragrance of an artificial pheromone known as androstenone had an increased inclination to approach and engage in flirtatious behavior with females. The detection of pheromones occurs via the vomeronasal organ, situated in the palatal region behind the incisors. The vomeronasal organ transmits messages to the brain, which then interprets these signals and subsequently affects behavior. Pheromones are believed to have various effects on sexual desire. Initially, they may effectively heighten individuals’ awareness of one another’s existence. Additionally, they have the potential to stimulate the secretion of hormones that enhance sexual pleasure. Furthermore, they have the potential to shape individuals’ evaluations of one another’s physical appeal.
5.3. Below are Many Study Publications That Have Examined the Influence of Pheromones on Human Sexual Attraction
This study examines the research supporting the influence of pheromones on human sexual attraction. The authors’ conclusion asserts the presence of compelling data demonstrating the capacity of pheromones to have an impact on human behavior, including the realm of sexual desire [36]. This study presents a thorough and extensive examination of the existing research on pheromones and their impact on human behavior. The author’s conclusion is that the data on the influence of pheromones on human sexual attraction remains inconclusive; however, there is an increasing body of evidence indicating their involvement [37].
According to this meta-analysis of 28 studies, pheromones do indeed have a modest but noteworthy influence on human sexual desire. Women had a more pronounced impact compared to men. According to this comprehensive study and synthesis of 44 investigations, it was shown that pheromones may have a modest but noteworthy impact on human sexual desire. The impact was more pronounced in males than in females. This study presents a thorough examination of the existing body of research on pheromones and their role in human mating. The author’s conclusion is that the data on the influence of pheromones on human sexual attraction remains inconclusive. However, there is an increasing body of evidence indicating their involvement [38].
In general, the study of pheromones and sexual attraction is now in its preliminary phase, although there is increasing evidence to indicate that pheromones do indeed have an impact on human sexual attraction. Further investigation is required to have a deeper comprehension of the mechanisms by which pheromones operate and their potential for augmenting sexual allure. Aside from the aforementioned study publications, many further studies have examined the influence of pheromones on human sexual desire. Several of these studies have had favorable outcomes, while others have yielded unfavorable outcomes. The collective assessment remains inconclusive; nevertheless, the available research strongly indicates that pheromones do really have an influence on human sexual desire.
5.4. Pheromones and Social Behavior
This research examined the impact of artificially produced human male pheromones on the sociosexual behavior of males. Thirty-eight heterosexual males, aged 26 to 42, participated in a 2-week initial period followed by a 6-week trial. The experiment was placebo-controlled and double-masked, evaluating a pheromone specifically created to enhance the romantic aspects of their lives. Each participant maintained a daily log documenting their engagement in six sociosexual behaviors: stroking, affection, kissing, formal dates, informal dates, sleeping next to a romantic partner, sexual intercourse, and self-stimulation leading to ejaculation (masturbation). These logs were faxed on a weekly basis. A much higher number of individuals using pheromones saw an increase in sexual intercourse and sleeping with a romantic partner compared to those using a placebo. There was a higher inclination for those using pheromones compared to those using a placebo to have an increase in petting, affection, kissing, and informal dating, but not in self-stimulation leading to ejaculation or in formal dates. A considerably greater percentage of those using pheromones, compared to those using a placebo, exhibited a rise in at least two, three or more of the five sociosexual activities with a female partner. Consequently, there was a notable rise in male sociosexual activities influenced by a woman’s sexual desire and collaboration but no increase in male masturbation, which only involves the guy. The preliminary studies need replication but indicate that human male pheromones influence the sexual appeal of males to women [39].
5.5. Pheromones in Plants
Plant pheromones include a wide range of chemicals, and their precise function is only sometimes fully known. Nevertheless, ongoing research in this field is progressing swiftly, revealing the significant involvement of plant pheromones in several plant functions, such as:
Pollination: Plant pheromones have the ability to allure pollinators towards flowers, thereby facilitating effective pollination and reproduction. Defense against herbivores and pests: Plant pheromones serve as a means to alert neighboring plants of potential threats, deter herbivores and pests, or entice predators that prey on herbivores and pests.
Interplant competition: Plant pheromones may be used to hinder the growth or prevent the germination of rival plants. Interactions with other creatures: Plant pheromones facilitate communication with other organisms, including beneficial bacteria and fungi, to foster mutually advantageous relationships.
The symbiotic relationship between a bee and a blooming plant, which is made possible by the release of plant pheromones, is shown in full in the image. This process is started by a blossoming plant with pink blooms, which release plant pheromones, which are shown in the illustration as dotted lines. Pheromones are chemical signals that travel through the atmosphere and allow plants and possible pollinators to communicate with one another. A bee is lured to the plant when it detects these pheromones because of the chemical signals. A key component of plant-pollinator dynamics is this interaction, in which the plant uses these chemical cues to draw in bees and other pollinators. These pollinators are then essential to the plant’s pollination process and subsequent reproduction.
5.6. Several Instances of Plant Pheromones Include
Methyl jasmonate is a pheromone emitted by plants when they are harmed by herbivores or pests. It serves as an attractant for predators that prey on herbivores and pests while simultaneously functioning as a warning signal to neighboring plants [40]. The compound is (Z)-3-hexene. The compound is named (Z)-3-hexen-1-ol. -1-ol is a pheromone secreted by some plants’ roots to attract advantageous microbes and fungi. These bacteria facilitate the absorption of nutrients by plants from the soil and provide protection against illnesses [41]. (E)-2-hexenal is a pheromone emitted by some plants to signal the presence of rival plants. This may elicit the secretion of defensive chemicals from the rival plants [42].
5.7. Utilization of Plant Pheromones
The use of plant pheromones has significant promise for many applications in the fields of agriculture and horticulture. Plant pheromones may serve several purposes, such as enhancing pollination in crops; pheromone lures may be used to attract pollinators, specifically targeting fruit trees and vegetable crops. Implementing this technique may enhance agricultural productivity and the quality of crops.
Prevent infestations and infections: Pheromone traps may be used to ensnare pests, while pheromone repellents can be utilized to discourage pests and illnesses from assaulting crops. Enhance plant growth and development: Plant pheromones have the potential to enhance plant growth and development, as well as enhance plants’ ability to withstand stress. Volatile Organic Compounds (VOCs): The process by which plants emit chemical compounds that have the ability to attract pollinators, deter herbivores, and facilitate communication between plants [43].
Plants emit chemical substances via many mechanisms, such as: Plants exude chemicals into the soil via their roots. These substances possess the ability to allure advantageous microorganisms, deter pests, or impede the sprouting of rival plants. Plants emit chemical substances into the atmosphere via their foliage. These chemicals possess the ability to allure pollinators, deter herbivores, or transmit signals to neighboring plants. Flowers emit a diverse array of chemical substances in order to allure pollinators. The compounds include nectar, fragrances, and pigments. Plants synthesize a diverse array of chemical compounds to lure pollinators. The compounds include nectar, fragrances, and pigments. Nectar is a viscous solution containing sugars that is secreted by flowers. It serves as nourishment for pollinators, including bees, butterflies, and bats. Aromas: Flowers emit a diverse range of aromas in order to allure pollinators. These fragrances might possess a sweet, flowery, or even fruity aroma. Flowers use color to allure pollinators. Certain flowers exhibit vibrant hues to lure pollinators from a distance, but others possess more subdued tones that captivate bees at close range.
5.8. Deterrence of Plant-Eating Animals
Plants also synthesize a diverse array of chemical substances to deter herbivores. These chemicals possess toxicity, an unpleasant taste, or the ability to cause irritation in the digestive tract of herbivores. Phytotoxins: Certain plants synthesize phytotoxins that are toxic to herbivorous organisms. As an example, the foxglove plant synthesizes a chemical known as digitalin, which is toxic to cardiac muscle [44]. Unpalatable chemicals: Some plants generate substances that have an unpleasant taste for herbivores. As an example, the milkweed plant generates a chemical known as cardenolide, which has a distinctly bitter flavor. Aggravating substances: Certain plants generate chemicals that provoke irritation in the digestive tract of herbivores. As an example, the chili pepper plant generates a chemical known as capsaicin, which causes irritation to the mucous membranes of the mouth and stomach [45].Plants use chemical molecules to engage in communication with other plants. When a plant is subjected to herbivore assault, it has the ability to emit chemical compounds that serve as a warning to other plants about the imminent threat. Subsequently, other plants have the capability to synthesize defensive chemicals as a means of safeguarding themselves from herbivorous organisms. Plants use chemical molecules to compete with each other. For instance, several plants emit chemical substances that impede the development of other plants that are in competition with them. Allelopathy: exploring how some plants release chemicals to inhibit the growth of nearby competing plants. Plants have the ability to generate and emit allelochemicals, which may disrupt the establishment and development of both similar and different plant species. Allelopathy has a crucial role in facilitating interactions between different plant species in both natural and controlled settings. The primary emphasis of this study is to examine the phenomenon of allelopathy and the role of allelochemicals in grasslands and forests. Allelopathy is the main factor behind plant invasion, worsens the deterioration of grasslands, and plays a role in the natural regeneration of forests. Moreover, pastures and tree plantations often experience autotoxicity, which refers to intraspecific allelopathy. Phenolics, terpenoids, and nitrogen-containing chemicals found in herbaceous and woody species have a role in allelopathy in grasslands and forests.Terpenoids, which have a wide range of metabolites, are qualitative allelochemicals found in annual grasslands. On the other hand, phenolics, which have a limited number of specialized metabolites, are quantitative allelochemicals found in permanent forests. Crucially, allelochemicals have a role in underground ecological interactions and plant-soil feedback, which in turn have an impact on the biodiversity, productivity, and sustainability of grasslands and forests. Allelopathic plants have the ability to distinguish the identity of neighboring plants by means of signaling molecules, which allows them to regulate the production of allelochemicals. Hence, allelochemicals and signaling chemicals work together in a synergistic manner to control both interactions between different species and interactions within the same species in grasslands and forests. The study of allelopathy and allelochemicals in grasslands and forests has yielded valuable knowledge on the interactions between plants and their impact on biodiversity, productivity, and sustainability. This research has significantly enhanced our understanding of terrestrial ecosystems and global environmental changes. Allelochemicals include a wide range of plant secondary metabolites, mostly categorized into three groups: phenolics, terpenoids, and nitrogen-containing substances [Table 3]. Phenolics are found in many different parts of plants and consist of a varied collection of molecules that have an aromatic ring with at least one hydroxyl group and maybe other additional groups, such as simple phenolic acids, coumarins, flavonoids, and quinones. Within forest ecosystems, a significant quantity of lignin derived from decaying organic matter undergoes decomposition, resulting in the formation of diverse phenolic acids. The lignin-derived phenolic acids are the primary allelochemicals found in forest soil, which result in a decrease in the quantity and richness of forest species [46].
Table 3: Comparison of pheromone products and their claims.
| Product | Manufacturer | Claims | Reference |
| Athena Pheromone Oils | Athena Pheromone | Contains human pheromones to attract mates, boost confidence, and reduce stress. | [64] |
| Evo-Science Pheromone Sprays | Evo-Science | Contains human pheromones to attract mates, increase social status, and improve mood. | [54] |
| Pheromone XS | Pheromone XS | Contains human pheromones to attract mates, increase sexual attraction, and boost self-esteem. | [22] |
| True Pheromones | True Pheromones | Contains human pheromones to attract mates, increase social intelligence, and reduce anxiety. | [1] |
| Alpha Male Pheromones | Alpha Male Pheromones | Contains human pheromones to attract women, increase dominance, and boost confidence | [55] |
Terpenoids, such as monoterpenes, sesquiterpenes, diterpenes, triterpenes, and steroids, are a group of molecules that originate from the 5-carbon isoprene. Monoterpenes and their derivatives exhibit high volatility and may interact with nearby plants in their gaseous form. The release of volatile allelochemicals by donor plants primarily affects neighboring plants via two primary mechanisms: the formation of “terpene clouds” that directly influence certain target plants [16] or the leaching of these chemicals into the soil, indirectly affecting other target plants. The volatile compounds produced by A. frigida include a large amount of terpenoids, namely the monoterpene camphor, which plays a significant role as an allelochemical in influencing nearby species [47]. Nitrogenous substances mostly include alkaloids, nonprotein amino acids, benzoxazinoids, and cyanogenic glycosides. Nitrogen-containing allelochemicals are less familiar compared to phenolics and terpenoids. Nevertheless, a number of distinct nitrogen-containing metabolites have been recognized as allelochemicals, that possess noteworthy ecological ramifications for grasslands and forests. Hexadecahydro-1-azachrysen-8-yl ester [Figure 3], which has been discovered as a possible alkaloid allelochemical in Imperata cylindrica, has the ability to inhibit root development and diminish mycorrhizal colonization [48]. There are various nonprotein amino acids involving allelopathic interferences with co-occurring species in grasslands. The presence of meta-Tyrosine in fine fescue grasses (Festuca rubra) might hinder the growth of other plants by inhibiting root formation [49].
REFERENCES
1. Law JH, Regnier FE. Pheromones. Annu Rev Biochem. 1971;40:533-48.
2. Gomez-Diaz C, Benton R. The joy of sex pheromones. EMBO Rep. 2013;14(10):874-83. [CrossRef].
3. Regnier FE, Law JH. Insect pheromones. J Lipid Res. 1968;9(5):541-51. [CrossRef].
4. Mason RT, et al. Sex pheromones in snakes. Science. 1989;245(4925):290-3. [CrossRef].
5. Roelofs WL, et al. Evolution of moth sex pheromones via ancestral genes. Proc Natl Acad Sci. 2002;99(21):13621-6. [CrossRef].
6. Wysocki CJ, Preti G. Facts, fallacies, fears, and frustrations with human pheromones. Ann N Y Acad Sci. 2004;1019:1201-11.
7. Verheggen FJ, Haubruge E, Mescher MC. Alarm pheromones—chemical signaling in response to danger. Vitam Horm. 2010;83:215-39.
8. Kurrey K, Ghosh S, Sinha JK. Alarm pheromones. In: Encyclopedia of Sexual Psychology and Behavior. Cham: Springer International Publishing; 2023:1-4. [CrossRef].
9. Lopes LE, et al. The alarm pheromone and alarm response of the clonal raider ant. J Chem Ecol. 2023;49:1-10. [CrossRef].
10. Wertheim B, et al. Pheromone-mediated aggregation in nonsocial arthropods: an evolutionary ecological perspective. Annu Rev Entomol. 2005;50(1):321-46.
11. Symonds MRE, Wertheim B. The mode of evolution of aggregation pheromones in Drosophila species. J Evol Biol. 2005;18(5):1253-63. [CrossRef].
12. Cardé RT. Defining attraction and aggregation pheromones: teleological versus functional perspectives. J Chem Ecol. 2014;40:519-20. [CrossRef].
13. Morgan ED. Trail pheromones of ants. Physiol Entomol. 2009;34(1):1-17. [CrossRef].
14. SUDD J. Interaction between ants on a scent trail. Nature. 1959;183:1588. [CrossRef].
15. Wheeler DE, Buck N, Evans JD. Expression of insulin pathway genes during the period of caste determination in the honey bee, Apis mellifera. Insect Mol Biol. 2006;15(5):597-602. [CrossRef].
16. Bind RH, Minney SM, Rosenfeld S, Hallock RM. The role of pheromonal responses in rodent behavior: future directions for the development of laboratory protocols. J Am Assoc Lab Anim Sci. 2013;52(2):124-9.
17. Fan AM, Ting D. In Encyclopedia of Toxicology (Third Edition). 2014.
18. Boulay R, Galarza JA, Chéron B, Hefetz A, Lenoir A, van Oudenhove L, Cerdá X. Intraspecific competition affects population size and resource allocation in an ant dispersing by colony fission. Ecology. 2010;91(11):3312-21. [CrossRef].
19. Sandler BH, Nikonova L, Leal WS, Clardy J. Sexual attraction in the silkworm moth: structure of the pheromone-binding-protein–bombykol complex. Chem Biol. 2000;7(2):143-51.
20. Blomquist GJ, Dillwith JW, Pomonis JG. Sex pheromone of the housefly: Metabolism of (Z)-9-tricosene to (Z)-9, 10-epxytricosane and (Z)-14-tricosen-10-one. Insect Biochem. 1984;14(3):279-84.
21. Müller-Schwarze D, Houlihan PW. Pheromonal activity of single castoreum constituents in beaver, castor canadensis. J Chem Ecol. 1991;17:715-34.
22. Araneda RC, Firestein S. The scents of androstenone in humans. J Physiol. 2004;554(1):1.
23. Boland W. The chemistry of gamete attraction: chemical structures, biosynthesis, and (a) biotic degradation of algal pheromones. Proc Natl Acad Sci. 1995;92(1):37-43. [CrossRef].
24. Mitaka Y, Matsuyama S, Mizumoto N, et al. Chemical identification of an aggregation pheromone in the termite Reticulitermes speratus. Sci Rep. 2020;10:7424. [CrossRef].
25. Prestwich GD. Chemical studies of pheromone reception and catabolism. Pheromone Biochemistry. 1987:473-527. [CrossRef].
26. Leal WS. Pheromone reception. The Chemistry of Pheromones and Other Semiochemicals II. 2005:1-36. [CrossRef].
27. Vogt RG. The molecular basis of pheromone reception: its influence on behavior. Pheromone Biochemistry. 1987:385-431. [CrossRef].
28. Prestwich GD. Chemical studies of pheromone receptors in insects. Arch Insect Biochem Physiol. 1993;22(1-2):75-86. [CrossRef].
29. Mori K. The synthesis of insect pheromones. In: Total Synthesis of Natural Products. 1981;4:1-183.
30. Bruce TJ, Aradottir GI, Smart LE, Martin JL, Caulfield JC, Doherty A, et al. The first crop plant was genetically engineered to release an insect pheromone for defense. Sci Rep. 2015;5(1):11183.
31. Dulac C. Molecular biology of pheromone perception in mammals. Semin Cell Dev Biol. 1997;8(2):197-205.
32. Cafazzo S, Natoli E, Valsecchi P. Scent-marking behavior in a pack of free-ranging domestic dogs. Ethology. 2012;118(10):955-66.
33. Witzgall P. Pheromones-future techniques for insect control? IOBC WPRS Bulletin. 2001;24(2):115-23.
34. Warren STH. Human pheromones: have they been demonstrated? Horm Behav. 2003;54(2):89-97.
35. Berliner DL, Jennings-White C, Lavker RM. The human skin: fragrances and pheromones. J Steroid Biochem Mol Biol. 1991;39(4):671-9.
36. Grammer K, Fink B, Neave N. Human pheromones and sexual attraction. Eur J Obstet Gynecol Reprod Biol. 2005;118(2):135-42.
37. Martín-Sánchez A, McLean L, Beynon RJ, Hurst JL, Ayala G, Lanuza E, et al. From sexual attraction to maternal aggression: When pheromones change their behavioral significance. Horm Behav. 2015;68:65-76.
38. Wu Y, Wei R, Nan Y, Hu Y. Androstadienone modulates human aggression in a sex-dependent manner. Soc Cogn Affect Neurosci. 2023;18(1):nsad006.
39. Cutler WB, Friedmann E, McCoy NL. Pheromonal influences on sociosexual behavior in men. Arch Sex Behav. 1998;27:1-13.
40. Yu X, Zhang W, Zhang Y, Zhang X, Lang D, Zhang X. The roles of methyl jasmonate to stress in plants. Funct Plant Biol. 2018;46(3):197-212.
41. Aschmann SM, Nishino N, Arey J, Atkinson R. Kinetics of the reactions of OH radicals with 2-and 3-methylfuran, 2,3-and 2,5-dimethylfuran, and E-and Z-3-hexene-2,5-dione, and products of OH+ 2,5-dimethylfuran. Environ Sci Technol. 2011;45(5):1859-65.
42. Kunishima M, Yamauchi Y, Mizutani M, Kuse M, Takikawa H, Sugimoto Y. Identification of (Z)-3:(E)-2-hexenal isomerases essential to the production of the leaf aldehyde in plants. J Biol Chem. 2016;291(27):14023-33.
43. Fang Y, Zeng R, Lu S, Dai L, Wan X. The synergistic attractiveness effect of plant volatiles to sex pheromones in a moth. J Asia-Pac Entomol. 2018;21(1):380-7.
44. Duke SO, Dayan FE. Modes of action of phytotoxins from plants. In: Allelopathy: a physiological process with ecological implications. Dordrecht: Springer Netherlands; 2006:511-36.
45. Yan L, Riaz M, Jiang C. Exogenous application of proline alleviates B-deficiency-induced injury while aggravating aluminum toxicity in trifoliate orange seedlings. Scientia Horticulturae. 2020;268:109372.
46. Xu Y, Chen X, Ding L, Kong CH. Allelopathy and allelochemicals in grasslands and forests. Forests. 2023;14(3):562. [CrossRef].
47. Li XF, Wang J, Huang D, et al. Allelopathic potential of Artemisia frigida and successional changes of plant communities in the northern China steppe. Plant Soil. 2011;341:383-98. [CrossRef].
48. Hagan DL, Jose S, Lin CH. Allelopathic exudates of Cogongrass (Imperata cylindrica): implications for the performance of native pine savanna plant species in the Southeastern US. J Chem Ecol. 2013;39:312-22. [CrossRef].
49. Bertin C, Weston LA, Huang T, Jander G, Owens T, Meinwald J, Schroeder FC. Grassroots chemistry: meta-tyrosine, an herbicidal nonprotein amino acid. Proc Natl Acad Sci. 2007;104(43):16964-9. [CrossRef].
50. Nollet F. Applications of pheromones in crop protection. In: Biopesticides Handbook. CRC Press; 2024:171-90.
51. Silverstein RM. Pheromones: background and potential for use in insect pest control. Science. 1981;213(4514):1326-32.
52. Pickett JA, Wadhams LJ, Woodcock CM. Developing sustainable pest control from chemical ecology. Agric Ecosyst Environ. 1997;64(2):149-56. [CrossRef].
53. Barclay H. Models for pest control using sex pheromones and chemosterilants. Int J Trop Insect Sci. 1987;8(2):187-96. [CrossRef].
54. Winman A. Do perfume additives termed human pheromones warrant being termed pheromones? Physiol Behav. 2004;82(4):697-701.
55. Pause BM. Is the human skin a pheromone-producing organ? J Cosmet Dermatol. 2004;3(4):223-8.
56. Mori K. Pheromone Synthesis. In: Schulz S, editor. The chemistry of pheromones and other semiochemicals I. Topics in Current Chemistry. Vol 239; 2004. [CrossRef].
57. Souza JP, Bandeira PT, Bergmann J, Zarbin PH. Recent advances in the synthesis of insect pheromones: an overview from 2013 to 2022. Nat Prod Rep. 2023:40:866-89. [CrossRef].
58. Zarbin PH, Villar JA, Corrêa AG. Insect pheromone synthesis in Brazil: an overview. J Braz Chem Soc. 2007;18:1100-24. [CrossRef].
59. Ishmuratov GY, Kharisov RY, Gazetdinov RR, Tolstikov GA. Insect pheromones are synthesized by oxidative transformations of natural monoterpenoids. Chem Nat Compd. 2005;41(6):617-35.
60. De Figueiredo RM, Berner R, Julis J, Liu T, Türp D, Christmann M. Bidirectional, organocatalytic synthesis of lepidopteran sex pheromones. J Org Chem. 2007;72(2):640-2. [CrossRef].
61. Guy JH, Burns SE, Barker JM, Edwards SA. Reducing post-mixing aggression and skin lesions in weaned pigs by application of a synthetic maternal pheromone. Animal Welfare. 2009;18(3):249-55. [CrossRef].
62. Westermann FL, Bell VA, Suckling DM, Lester PJ. Synthetic pheromones as a management technique–dispensers reduce Linepithema humile activity in a commercial vineyard. Pest Manag Sci. 2016;72(4):719-24. [CrossRef]
63. Wyatt TD. Pheromones. Curr Biol. 2017;27(15):R739-43.
64. Clifton J. The erotics of the axillary pose. VENUE. 2022;1(1):22-48.
65. Precone V, Paolacci S, Beccari T, Ragione LD, Stuppia L, Baglivo M, et al. Pheromone receptors and their putative ligands: possible role in humans. Euro Rev Med Pharmacol Sci. 2020;24(4):2140-50.
66. Mostafa T, El Khoury G, Hassan A. Pheromones in sex and reproduction: do they have a role in humans? J Adv Res. 2012;3(1):1-9.
67. Kirsch P. Pheromones: their potential role in control of agricultural insect pests. Am J Alternat Agri. 1988;3(2-3):83-97. [CrossRef].
68. Mishra SS, Shroff S, Sahu JK, Naik PP, Baitharu I. Insect pheromones and its applications in management of pest population. In: Kumar D, Shahid M (eds) Natural Materials and Products from Insects: Chemistry and Applications. Cham: Springer; 2020. [CrossRef]
69. Rizvi SAH, George J, Reddy GVP, Zeng X, Guerrero A. Latest developments in insect sex pheromone research and its application in agricultural pest management. Insects. 2021;12(6):484. [CrossRef]
70. Kausar MA, Rabie AM, Azam F, Anwar S, Alenazi F, Alshammari KF, et al. The role of Aedes aegypti in inducing/aggravating IgE-mediated allergic airway disease: extensive computational studies for identification of allergenic proteins. J Biomol Struc Dyn. 2024;42(5):2738-45.