1. INTRODUCTION
Citrus trees are perennials that are commonly subjected to soil and atmospheric droughts. Citrus groves typically require irrigation in arid and semi-arid areas. Periods of water deficit harm yield and vegetative growth and minimize fruit size and occasionally quality, resulting in considerable economic losses [1-3]. In addition, stomatal conductance, transpiration, and CO2 assimilation are decreased by water stress [4-6]. The root system signals the leaves to induce stomatal closure [7]. Different drought-resistant strategies have been developed by plants, such as decreased leaf mass or enhanced root development [8,9]. Osmotic adjustment helps plants support growth and photosynthesis by maintaining the leaf turgor required to open the stomata [4]. Drought-affected plants experience significant damage owing to the secondary effects of oxidative stress. This stress is a consequence of the downregulation of the photosynthetic process, which in turn causes alterations in the electron transport chain during photosynthetic reactions. Consequently, reactive oxygen species (ROS) are generated when the photosynthetic process is insufficiently active [10]. Excessive ROS production may damage nucleic acids, proteins, and lipids. These substances can then be oxidized and undergo detrimental effects such as enzyme inhibition, chlorophyll degradation, membrane disruption, loss of organelle function, decreased metabolic efficiency, and carbon fixation [11]. Plants prevent cell dehydration by either promoting water inflow through the accumulation of active solutes that decrease osmotic potential or limiting water efflux through the hardness of cell walls [12]. Plants produce several essential solutes, such as sugars, polyols, polyamines, proline, glycine, and betaine [13]. Proline is the most studied amino acid among the previously listed osmolytes [14]. Soluble sugars are important osmoprotectants, which means that they are essential for cellular osmotic adjustment and shield cellular structures from external stress. Rangpur lime is one of the most drought-tolerant rootstocks, while Poncirus trifoliata (trifoliate orange) is considered susceptible to drought [15]. The present experiment aims to study the behavior of six new hybrid rootstocks under three different water stress conditions in a greenhouse based on morphological, physiological, and biochemical indicators.
3. RESULTS
3.1 Effect of Water Stress on the Stem Height, Diameter, and Number of Leaves
Water stress caused a reduction in all the growth parameters monitored, with the reduction being most noticeable at the 50% FC level. The variation in the percentage reduction in stem height [Figure 1], stem diameter [Figure 2], and the number of leaves [Table 2]. For stem height, at 75% FC, rootstock H2 had the highest percentage reduction (66%), and rootstock H6 had the lowest (24%). At 50% FC, the H6 rootstock had the lowest percentage reduction (48%), and the H2 hybrid had the highest reduction (76%). For stem diameter, at 75% FC, the H5 rootstock had the lowest percentage reduction (12%), and the H2 hybrid had the highest reduction (57%). At 50% FC, the highest percentage of reduction was recorded for H2 (74%), and the lowest for H5 (40%). In terms of leaf number, for the controls, hybrid H4 had the highest number of 25 leaves, and hybrids H1 and H5 had the lowest number (17). At 75% FC, hybrids H1 and H5 had the highest number of 14 leaves, and hybrid H1 had the lowest. At 50% FC, hybrid H2 had the highest number of 8 leaves, and hybrid H3 had the lower of 1 leaf [Figure 3].
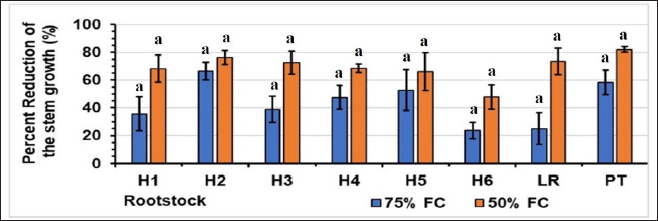 | Figure 1: Effect of water stress on the stem growth Stress levels with the same letter do not differ significantly for the same rootstock; at p ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
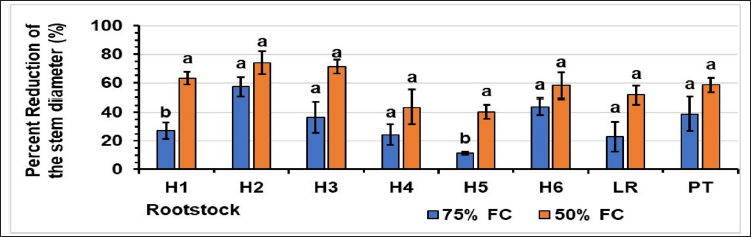 | Figure 2: Effect of water stress on the stem diameter Stress levels with the same letter do not differ significantly for the same rootstock; at p ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
Table 2: Effect of water stress on number of leaves.
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 17 ± 0.67a | 11 ± 1.21b | 7 ± 1.54c |
| H2 | 22 ± 3.31a | 14 ± 5.06ab | 8 ± 2.92b |
| H3 | 19 ± 3.31a | 13 ± 1.00a | 1 ± 0.43b |
| H4 | 25 ± 2.52a | 13 ± 2.23b | 2 ± 2.08c |
| H5 | 17 ± 2.12a | 14 ± 1.00b | 7 ± 1.02c |
| H6 | 19 ± 3.18a | 12 ± 2.11ab | 3 ± 1.29b |
| LR | 36 ± 7.79a | 17 ± 2.88ab | 9 ± 2.05b |
| PT | 6 ± 2.00a | 2 ± 0.79ab | –3 ± 1.35b |
Stress levels with the same letter do not differ significantly for the same rootstock; at p ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), mean values ± standard error (n = 6).
 | Figure 3: Effect of water stress on the appearance of the hybrid citrus rootstocks tested: (A) 100% of the field capacity, (B) 75% of the field capacity, (C) 50% of the field capacity.
[Click here to view] |
3.2 Effect of Water Stress on the Stomatal Conductance of Leaves
According to the results in Figure 4, stomatal conductance varies according to the rootstock and irrigation dose. The stomatal conductance of control rootstocks was high, whereas that of rootstocks irrigated with 50% FC was low. For the controls, hybrid H2 had the highest value of 19.70 mmol/m2/s, and hybrid H5 had the lowest value of 15.75. For rootstocks irrigated with 75% FC, the highest stomatal conductance (14.53) was observed for hybrid H2, and the lowest value of 9.78 was recorded for hybrid H3. For the 50% FC level, rootstock H3 recorded the lowest stomatal conductance of 4.13 mmol/m2/s, while genotype H5 had the highest value of 6.85 mmol/m2/s.
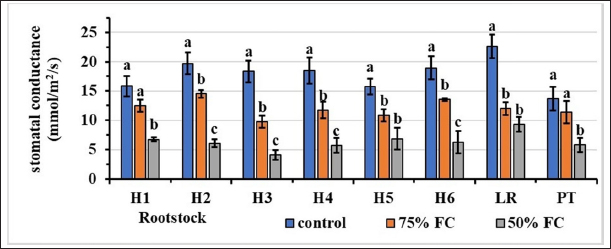 | Figure 4: Effect of water stress on stomatal conductance (mmol/m2/s) Stress levels with the same letter do not differ significantly for the same rootstock; at p ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
3.3 Effect of Water Stress on the FM of Leaves, Stems, and Roots
The water stress reduced the FM of different parts of the seedlings for all rootstocks: leaves [Table 3], stems [Table 4], and roots [Table 5]. For leaves, the mass of FM varied between 12.89 g and 2.56 g. For controls, the H2 rootstock had the highest mass, and the H1 rootstock had the lowest mass. For stems and roots, rootstock H3 had the highest mass (9.37 g), and hybrid H5 had the lowest mass (1.84 g). For roots, the root mass of hybrid H5 at 75% FC is higher than that of control, 10.71–10.40 g, respectively, and hybrid H6 has the same mass for control and 75% FC.
Table 3: Effect of water stress on the fresh matter of leaves (g).
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 7.18 ± 0.73a | 5.34 ± 0.12b | 4.00 ± 0.21b |
| H2 | 12.89 ± 2.60a | 7.31 ± 0.57ab | 4.27 ± 0.90b |
| H3 | 12.75 ± 1.73a | 7.02 ± 0.50b | 3.43 ± 0.45b |
| H4 | 9.90 ± 0.44a | 7.04 ± 0.20b | 2.98 ± 0.96c |
| H5 | 9.38 ± 0.87a | 6.19 ± 0.67b | 3.39 ± 0.54c |
| H6 | 9.24 ± 0.87a | 6.13 ± 0.75b | 2.56 ± 0.38c |
| LR | 12.68 ± 0.95a | 7.09 ± 0.33b | 3.40 ± 0.80c |
| PT | 1.89 ± 0.17a | 1.65 ± 0.13a | 1.11 ± 0.40a |
Table 4: Effect of water stress on the fresh matter of stems (g).
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 3.93 ± 0.78a | 2.67 ± 0.23ab | 2.07 ± 0.08b |
| H2 | 9.28 ± 1.56a | 5.29 ± 0.26b | 3.35 ± 0.38b |
| H3 | 9.37 ± 0.43a | 4.12 ± 0.31b | 3.01 ± 0.45b |
| H4 | 5.99 ± 0.20a | 4.30 ± 0.08b | 2.95 ± 0.40c |
| H5 | 3.96 ± 0.88a | 2.20 ± 0.29ab | 1.84 ± 0.18b |
| H6 | 5.38 ± 0.01a | 3.52 ± 0.12ab | 2.11 ± 0.34c |
| LR | 6.57 ± 0.35a | 4.39 ± 0.11b | 3.04 ± 0.57b |
| PT | 4.11 ± 0.54a | 3.28 ± 0.33a | 2.79 ± 0.15a |
Table 5: Effect of water stress on fresh matter of roots (g).
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 7.93 ± 2.25a | 6.12 ± 1.04a | 5.30 ± 0.7a |
| H2 | 13.21 ± 3.25a | 9.77 ± 2.16a | 6.52 ± 0.82a |
| H3 | 14.27 ± 3.97a | 9.45 ± 2.16a | 7.43 ± 0.26a |
| H4 | 10.4 ± 1.67a | 10.71 ± 3.00a | 5.63 ± 1.72a |
| H5 | 7.72 ± 2.15a | 5.74 ± 1.54a | 3.98 ± 0.70a |
| H6 | 7.70 ± 1.73a | 7.71 ± 1.34a | 4.48 ± 0.65a |
| LR | 8.5 ± 0.90ab | 12.67 ± 3.52a | 4.88 ± 0.59b |
| PT | 6.12 ± 1.37a | 6.23 ± 1.65a | 5.64 ± 1.80a |
3.4 Effect of Water Stress on the Dry Matter of Leaves, Stems, and Roots
The water stress reduced the dry matter of the different parts of the seedlings: leaves [Table 6], stems [Table 7], and roots [Table 8]. The variation in dry matter was relatively same as that of FM, except for roots, where the root mass of H2, H4, and H6 hybrids at 75% FC was greater than that of the controls.
Table 6: Effect of water stress on the dry matter of leaves (g).
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 2.34 ± 0.24a | 1.76 ± 0.10b | 1.43 ± 0.06c |
| H2 | 4.38 ± 0.42a | 2.49 ± 0.15b | 1.47 ± 0.16c |
| H3 | 4.07 ± 0.37a | 2.51 ± 0.25b | 1.29 ± 0.13c |
| H4 | 3.21 ± 0.15a | 2.46 ± 0.13a | 1.12 ± 0.35b |
| H5 | 3.00 ± 0.39a | 2.04 ± 0.24ab | 1.28 ± 0.22b |
| H6 | 2.95 ± 0.11a | 2.15 ± 0.22b | 0.96 ± 0.17c |
| LR | 4.02 ± 0.45a | 2.89 ± 0.23a | 1.27 ± 0.33b |
| PT | 0.68 ± 0.07a | 0.65 ± 0.06a | 0.46 ± 0.13a |
Table 7: Effect of water stress on the dry matter of stems (g)
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 1.31 ± 0.17a | 1.01 ± 0.04b | 0.98 ± 0.04b |
| H2 | 3.84 ± 0.49a | 2.30 ± 0.08b | 1.64 ± 0.21b |
| H3 | 3.43 ± 0.29a | 1.94 ± 0.12b | 1.45 ± 0.19b |
| H4 | 2.43 ± 0.11a | 1.77 ± 0.36b | 1.42 ± 0.16c |
| H5 | 1.60 ± 0.31a | 0.91 ± 0.14a | 0.98 ± 0.09a |
| H6 | 2.02 ± 0.04a | 1.40 ± 0.21b | 1.01 ± 0.22b |
| LR | 2.94 ± 0.28a | 2.42 ± 0.32b | 1.54 ± 0.37b |
| PT | 1.65 ± 0.21a | 1.36 ± 0.07a | 1.11 ± 0.20a |
Table 8: Effect of water stress on the dry matter of roots (g).
| Rootstock | Control | 75% FC | 50% FC |
|---|
| H1 | 2.86 ± 0.39a | 2.01 ± 0.24a | 2.27 ± 0.20a |
| H2 | 4.14 ± 0.49a | 4.41 ± 0.76a | 3.27 ± 0.60a |
| H3 | 5.15 ± 0.71a | 4.37 ± 0.87a | 3.69 ± 0.18a |
| H4 | 3.63 ± 0.60a | 4.57 ± 1.21a | 2.81 ± 0.76a |
| H5 | 2.52 ± 0.33a | 2.04 ± 0.58a | 1.89 ± 0.39a |
| H6 | 2.46 ± 0.09b | 3.42 ± 0.32a | 2.12 ± 0.32b |
| LR | 2.92 ± 0.20b | 6.81 ± 1.54a | 2.43 ± 0.34b |
| PT | 2.09 ± 0.13a | 2.25 ± 0.60a | 2.51 ± 0.85a |
3.5 Effect of Water Stress on the Chlorophyll Content of Leaves
Figure 5 shows that the irrigation dose reduced the chlorophyll content of rootstocks. For the controls, chlorophyll content was high, with the H3 rootstock having the highest value of 3.53 mg/g FM and the H4 hybrid having the lowest at 2.78. At 75% FC, rootstock H3 had the highest chlorophyll content of 3.09, rootstock H4 had 3.53 mg/g FM, and the lowest content of 2.01. In contrast, at 50% FC, the highest chlorophyll content of 2.67 mg/g FM was recorded for hybrid H3, and hybrid H5 had the lowest content of 1.47 mg/g FM.
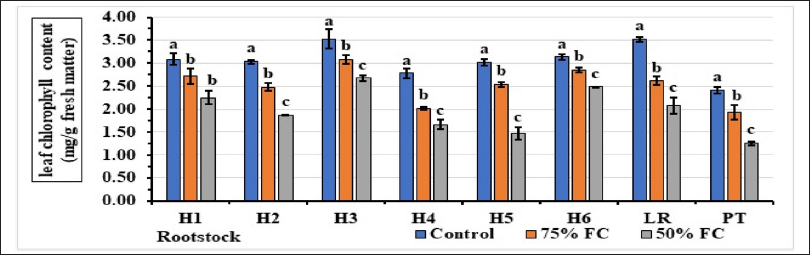 | Figure 5: Effect of water stress on leaf chlorophyll content. Stress levels with the same letter do not differ significantly for the same rootstock; at p ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
3.6 Effect of Water Stress on Leaf Proline Content
The results in Figure 6 show that water stress triggers proline accumulation in the leaves. For the controls, the highest proline content (49.2) was observed for hybrid H4, and the lowest was 34.9 for hybrid H1. At 75% FC, rootstock H4 had the highest content (85.8), and rootstock H1 had the lowest content (54.6). At the 50% level, hybrid H4 had the highest proline content of 122.1 µg/g DM, while hybrid H6 had the lowest at 85.3 µg/g.
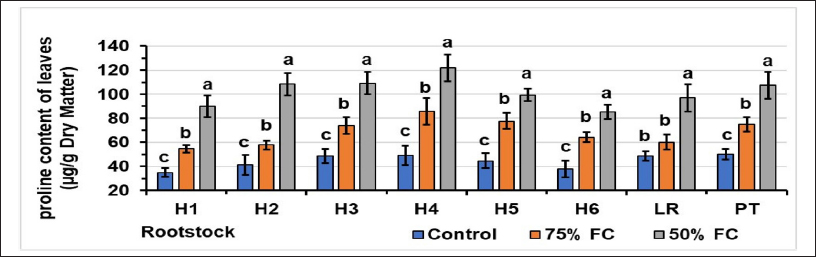 | Figure 6: Effect of water stress on proline content of leaves. Stress levels with the same letter do not differ significantly for the same rootstock; at P ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
3.7 Effect of Water Stress on the Soluble Sugar Content of Leaves
As shown in Figure 7, water stress increased the leaf sugar content of the rootstocks tested. For the controls, the sugar content was high, with the H3 rootstock at 2.36 mg/g DM and the lowest content of 1.53 observed for the H4 hybrid. At 75% FC, rootstock H3 had the highest sugar content (3.36), and rootstock H5 had the lowest (2.12). At 50% FC, the highest sugar content of 4.24 mg/g DM was recorded for hybrid H3, whereas hybrids H4 and H5 had the lowest content (2.60 mg/g).
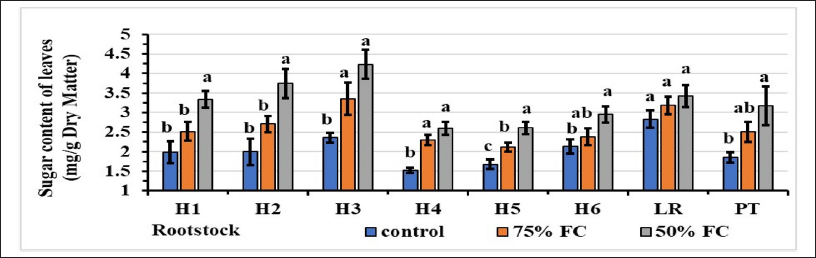 | Figure 7: Effect of water stress on soluble sugar content of leaves. Stress levels with the same letter do not differ significantly for the same rootstock; at P ≤ 0.05 (one-way ANOVA, separated by Duncan’s test), vertical bars represent the standard error (n = 6).
[Click here to view] |
4. DISCUSSION
Water stress is one of the main environmental factors that affect the development, growth, and productivity of plants. Citrus is often exposed to water shortages, particularly in arid and semiarid areas. The choice of rootstock is among the most important decisions a grower makes. Citrus rootstock influences the morphological, biochemical, and physiological characteristics of the grafted scion. A rootstock with drought tolerance is of great importance to remedy this climatic hazard and reduce production losses. In the present study, we evaluated the behavior of new hybrid citrus rootstocks under water-stress conditions in a greenhouse. The effects of 50% FC and 75% FC on stem height, stem diameter, number of leaves, stomatal conductance, fresh and dry biomass, total chlorophyll, soluble sugars, and proline were studied. The results showed that stem height, number of leaves, and stem diameter were affected by water stress, with a significant reduction in these parameters at the 50% FC level. The hybrid H6 has the lowest stem growth reduction; in contrast, the hybrid H2 has the highest stem growth reduction. In terms of leaves, hybrid H2 had the highest number of leaves, whereas hybrid H3 had the lowest. Indeed, the application of water stress to seedlings caused a reduction in stem diameter. Similar findings have been reported by Wu et al. [26] and Rodriguez-Gamir et al. [27]. Water stress changes the water relationship within the plants and decreases turgor, gas exchange, and growth [28]. Shafqat et al. [29] revealed that the ability of plants to maintain growth under limited water indicates their tolerance ability. Beniken et al. [18] reported that water stress caused leaf drop and a reduction in stem height. In the case of severe 50% FC water stress, variability between rootstocks was observed in growth, which could be explained by a reduction in photosynthetic and metabolic reactions that are limited to biosynthetic pathways. Similar results have been reported by Wu et al. [26] and Rodriguez-Gamir et al. [27]. The fresh and dry biomass of various plant parts was affected by stress. The lowest fresh matter of leaves was observed in the hybrid H6, compared to the hybrid H1, which showed the greatest tolerance. The reduction of fresh and dry matter shown in all hybrids and more pronounced in 50% FC may be the result of physiological and biochemical disturbance, which may be accentuated with the reduction of leaf area and leaf drop [30,31]. Craine [30] and Munns et al. [32] stated that practically all rootstocks examined showed inhibitory impacts of water stress on biomass. Our study also demonstrated the effect of water stress on stomatal conductance, since stomatal closure lowers photosynthesis and water loss. Our results are in agreement with those reported by Rodriguez-Gamir et al. [27], Hutton and Loveys [33], and De Campos et al. [34], who reported that this decrease in stomatal conductance is due to stomatal closure and a consequent reduction in transpiration, which also leads to a reduction in photosynthetic responses. Boyer and Kramer [5] reported that when water stress is experienced by plants, reducing stomatal conductance is the first short-term response to prevent water loss through transpiration. In the medium term, increasing root development maximizes water intake. García-Sánchez et al. [4] and Beniken et al. [18] reported that the tolerance capacity of these rootstocks is linked to their high osmotic adjustment capacity via osmolyte accumulation, which enables them to maintain their vital photosynthetic activities under water-stress conditions. Results show that the chlorophyll content decreased with the increase in stress. At 50% FC, the highest chlorophyll content was recorded for the hybrid H3 and the lowest for H5. Hussain et al. [35] reported that water stress decreases the chlorophyll content of citrus rootstock. Citrus rootstock with high chlorophyll content in water stress conditions is considered tolerant [36]. Water stress affects the normal functioning of photosynthetic machinery; degradation and photooxidation of chlorophyll caused by transpiration imbalance disturb the plant’s capacity to harvest light and reduce total photosynthetic output, leading to a reduction in carbohydrate transport and growth [37]. Several authors, Cohen and Naor [38], Weibel et al. [39], and Gijón et al. [40] have reported that the drop in chlorophyll content is the result of reduced stomatal opening. This stomatal closure limits water loss through evapotranspiration while simultaneously reducing the inflow of atmospheric CO2 required for photosynthesis. Proline content increased with the increase in stress levels. Among the hybrids tested, the highest content was recorded in the hybrid H4, whereas the lowest content was observed in the hybrid H6. Hussain et al. [35] observed an increase in proline content in citrus rootstock when subjected to water stress. Proline, as an osmoprotectant compound, prevents macromolecules from dehydration, adjusts osmotic pressure, protects proteins from denaturation, detoxifies reactive oxygen species, and protects membranes against lipid peroxidation [28,41]. Shekafandeh et al. [42] observed that tolerant genotypes accumulate more proline than sensible genotypes. Other authors, Khedr et al. [43], Demiral and Türkan [44], and Ma et al. [45], have reported that proline strengthens the antioxidant system and combats stress damage. It may also play a role in regulating the cytoplasmic pH or constitute a nitrogen reserve used by the plant after the stress period. Sugar content increased with the increase in stress levels. Among the hybrids tested, the highest content was recorded in the hybrid H3, whereas the lowest content was observed in the hybrids H4 and H5. Plants develop other tolerance mechanisms consisting of the accumulation of solutes such as sugars, which reduce osmotic potential at the cellular level by osmotic adjustment in the sense of keeping water in the cell. Similar results have been reported in citrus fruits by Molinari et al. [46] and Beniken et al. [18]. These researchers observed that the total soluble sugar content of rootstock leaves increases with the severity of water stress. This increase could be a parameter of adaptation to water-deficit conditions, as it helps maintain high cellular integrity at the tissue level.
REFERENCES
1. Levy Y, Shalhevet J, Bielorai H. Effect of irrigation regime and water salinity on grapefruit quality. J Am Soc Hortic Sci. 1979;104(3):356-9. [CrossRef]
2. Levy Y, Bielorai H, Shalhevet J. Long-term effects of different irrigation regimes on grapefruit tree development and yield. J Am Soc Hortic Sci. 1978;103(5):680-3. [CrossRef]
3. Pérez-Pérez JG, Romero P, Navarro JM, Botía P. Response of sweet orange cv “Lane late” to deficit irrigation in two rootstocks. I: Water relations, leaf gas exchange and vegetative growth. Irrig Sci. 2008;26(5):415-25. [CrossRef]
4. García-Sánchez F, Syvertsen JP, Gimeno V, Botía P, Perez-Perez JG. Responses to flooding and drought stress by two citrus rootstock seedlings with different water-use efficiency. Physiol Plant. 2007;130(4):532-42. [CrossRef]
5. Boyer JS, Kramer PJ. Water Relations of Plants and Soils. 1st ed. Academic Press. San Diego;1995.
6. Syvertsen JP, Lloyd J, Kriedemann PE. Salinity and drought stress effects on foliar ion concentration, water relations, and photosynthetic characteristics of orchard citrus. Aust J Agric Res. 1988;39(4):619-27. [CrossRef]
7. Davies WJ, Zhang J. Root signals and the regulation of growth and development of plants in drying soil. Annu Rev Plant Physiol Plant Mol Biol. 1991;42(1):55-76. [CrossRef]
8. Lei Y, Yin C, Li C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol Plant. 2006;127(2):182-91. [CrossRef]
9. Zhang J, Davies WJ. Sequential response of whole plant water relations to prolonged soil drying and the involvement of Xylem Sap ABA in the regulation of stomatal behaviour of sunflower plants. New Phytol. 1989;113(2):167-74. [CrossRef]
10. Apel K, Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol. 2004;55(1):373-99. [CrossRef]
11. Scandalios JG. Oxidative stress: molecular perception and transduction of signals triggering antioxidant gene defenses. Braz J Med Biol Res. 2005;38(7):995-1014. [CrossRef]
12. Zhang J, Nguyen HT, Blum A. Genetic analysis of osmotic adjustment in crop plants. J Exp Bot. 1999;50(332):291-302. [CrossRef]
13. Parida AK, Das AB. Salt tolerance and salinity effects on plants: a review. Ecotoxicol Environ Saf. 2005;60(3):324-49. [CrossRef]
14. Szabados L, Savouré A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010;15(2):89-97. [CrossRef]
15. Davies FS, Albrigo LG. Citrus. 1st ed. Wallingford: Oxford University Press; 1994.
16. Hoagland DR, Arnon DI. The water-culture method for growing plants without soil. Circ Calif Agric Exp Stn. 1950;347:1-39.
17. Lourkisti R, Oustric J, Quilichini Y, Froelicher Y, Herbette S, Morillon R, et al. Improved response of triploid citrus varieties to water deficit is related to anatomical and cytological properties. Plant Physiol Biochem. 2021;162:762-75. [CrossRef]
18. Beniken L, Beqqali M, Dahan R, Benkirane R, Omari FE, Benazouz A, et al. Evaluation of resistance of ten citrus rootstocks resistant to tristeza regarding the water deficit. Fruits. 2011; 66(6):373-84. [CrossRef]
19. Pedroso FKJV, Prudente DA, Bueno ACR, Machado EC, Ribeiro RV. Drought tolerance in citrus trees is enhanced by rootstock-dependent changes in root growth and carbohydrate availability. Environ Exp Bot. 2014;101:26-35. [CrossRef]
20. Boris R, Eran R, Tanya G, Jhonathan EE, Shimon R. Phenotypic plasticity and water flux rates of Citrus root orders under salinity. J Exp Bot. 2012;63(7):2717-27. [CrossRef]
21. Bañuls J, Serna MD, Legaz F, Talon M, Primo-Millo E. Growth and gas exchange parameters of Citrus plants stressed with different salts. J Plant Physiol. 1997;150(1):194-9. [CrossRef]
22. Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water-stress studies. Plant Soil. 1973;39(1):205-7. [CrossRef]
23. DuBois M, Gilles KA, Hamilton JK, Rebers PA, Fred S. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350-56. [CrossRef]
24. Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949;24(1):1-15. [CrossRef]
25. Ahrens WH, Cox DJ, Budhwar G. Use of the arcsine and square root transformations for subjectively determined percentage data. Weed Sci. 1990;38(4-5):452-8. [CrossRef]
26. Wu Q-S, Xia R-X, Zou Y-N. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. Eur J Soil Biol. 2008;44(1):122-8. [CrossRef]
27. Rodríguez-Gamir J, Ancillo G, Aparicio F, Bordas M, Primo-Millo E, Forner-Giner MÁ. Water-deficit tolerance in citrus is mediated by the down regulation of PIP gene expression in the roots. Plant Soil. 2011;347(1):91-104. [CrossRef]
28. Alam A, Ullah H, Attia A, Datta A. Effects of salinity stress on growth, mineral nutrient accumulation and biochemical parameters of seedlings of three citrus rootstocks. Int J Fruit Sci. 2020;20(4):786-804. [CrossRef]
29. Shafqat W, Mazrou YSA, Nehela Y, Ikram S, Bibi S, Naqvi SA, et al. Effect of three water regimes on the physiological and anatomical structure of stem and leaves of different citrus rootstocks with distinct degrees of tolerance to drought stress. Hortic. 2021;7(12):554. [CrossRef]
30. Craine JM. Reconciling plant strategy theories of Grime and Tilman. J Ecol. 2005;93(6):1041-52. [CrossRef]
31. Romero-Aranda R, Moya JL, Tadeo FR, Legaz F, Primo-Millo E, Talon M. Physiological and anatomical disturbances induced by chloride salts in sensitive and tolerant citrus: beneficial and detrimental effects of cations. Plant, Cell & Environment. 1998;21(12):1243-53. [CrossRef]
32. Munns R, James RA, Läuchli A. Approaches to increasing the salt tolerance of wheat and other cereals. J Exp Bot. 2006;57(5):1025-43. [CrossRef]
33. Hutton RJ, Loveys BR. A partial root zone drying irrigation strategy for citrus—effects on water use efficiency and fruit characteristics. Agric Water Manag. 2011;98(10):1485-96. [CrossRef]
34. de Campos MKF, de Carvalho K, de Souza FS, Marur CJ, Pereira LFP, Filho JCB, et al. Drought tolerance and antioxidant enzymatic activity in transgenic “Swingle” citrumelo plants over-accumulating proline. Environ Exp Bot. 2011;72(2):242-50. [CrossRef]
35. Hussain S, Khalid MF, Saqib M, Ahmad S, Zafar W, Rao MJ, et al. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol Plant. 2018;40(8):135. [CrossRef]
36. Dbira S, Al Hassan M, Gramazio P, Ferchichi A, Vicente O, Prohens J, et al. Variable levels of tolerance to water stress (drought) and associated biochemical markers in tunisian barley landraces. Molecules. 2018;23(3):613. [CrossRef]
37. Anjum SA, Xie XY, Wang LC, Saleem MF, Man C, Lei W. Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agri Res. 2011;6(9):2026-32. [CrossRef]
38. Cohen S, Naor A. The effect of three rootstocks on water use, canopy conductance and hydraulic parameters of apple trees and predicting canopy from hydraulic conductance. Plant Cell Environ. 2002; 25(1):17-28. [CrossRef]
39. Weibel A, Johnson RS, DeJong TM. Comparative vegetative growth responses of two peach cultivars grown on size-controlling versus standard rootstocks. J Am Soc Hortic Sci. 2003;128(4):463-71. [CrossRef]
40. Gijón M del C, Gimenez C, Perez-López D, Guerrero J, Couceiro JF, Moriana A. Rootstock influences the response of pistachio (Pistacia vera L. cv. Kerman) to water stress and rehydration. Sci Hortic. 2010;125(4):666-71. [CrossRef]
41. Martínez-Cuenca MR, Primo-Capella A, Forner-Giner MA. Screening of ‘King’ mandarin (Citrus nobilis Lour) × Poncirus trifoliata ((L.) Raf.) hybrids as salt stress-tolerant citrus rootstocks. Hortic Environ Biotechnol. 2021;62(10):337-51. [CrossRef]
42. Shekafandeh A, Assar P, Taghipour L. Cell membrane stability and water relations of different scions grafted onto Mexican lime under water stress conditions and recovery. Plant Physiol Rep 2019;24(2):182-91. [CrossRef]
43. Khedr AHA, Abbas MA, Wahid AAA, Quick WP, Abogadallah GM. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J Exp Bot. 2003; 54(392):2553-62. [CrossRef]
44. Demiral T, Türkan I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ Exp Bot. 2005;53(3):247-57. [CrossRef]
45. Ma Q-Q, Wang W, Li Y-H, Li D-Q, Zou Q. Alleviation of photoinhibition in drought-stressed wheat (Triticum aestivum) by foliar-applied glycinebetaine. J Plant Physiol. 2006;163(2):165-75. [CrossRef]
46. Molinari HBC, Marur CJ, Filho JCB, Kobayashi AK, Pileggi M, Júnior RPL, et al. Osmotic adjustment in transgenic citrus rootstock Carrizo citrange (Citrus sinensis Osb. x Poncirus trifoliata L. Raf.) overproducing proline. Plant Sci. 2004; 167(6):1375-81. [CrossRef]
