1. INTRODUCTION
Monoclonal antibodies (mAbs) are immunoglobulins that exhibit a high level of specificity, targeting a single antigen or epitope. mAbs are usually obtained from a clonal proliferation of cancerous human plasma cells that produce antibodies. In 1975, George Köhler and Ceasar Milstein developed hybridoma technology, which spurred new optimism. To create human-derived hybridomas, Kohler and Milstein used human–mouse hybrid cells, which have since become a cornerstone in the large-scale manufacturing of therapeutic antibodies [1,2]. Early therapeutic mAbs, derived from mice, were immunogenic in humans and had limited efficacy in stimulating immune responses in patients, hence restricting their clinical usefulness. In order to overcome these restrictions, methodologies for producing antibodies more human-like were devised throughout the later part of the 1980s [3,4]. However, the advent of novel technologies including recombinant DNA technology, phage display, and transgenic mice has led to the development of numerous kinds of mAbs, primarily chimeric, humanized, and fully human antibodies. These showed lower immunogenicity and greater efficacy as drug products.
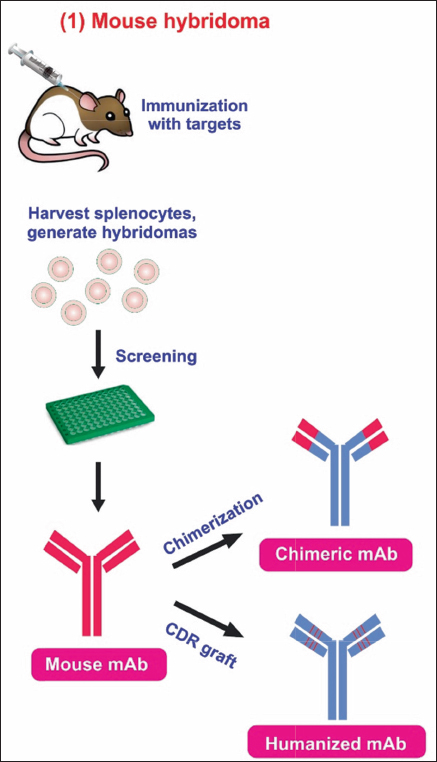
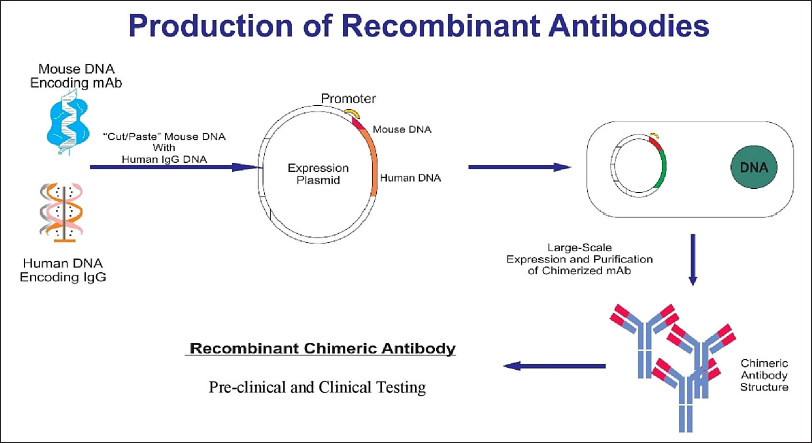
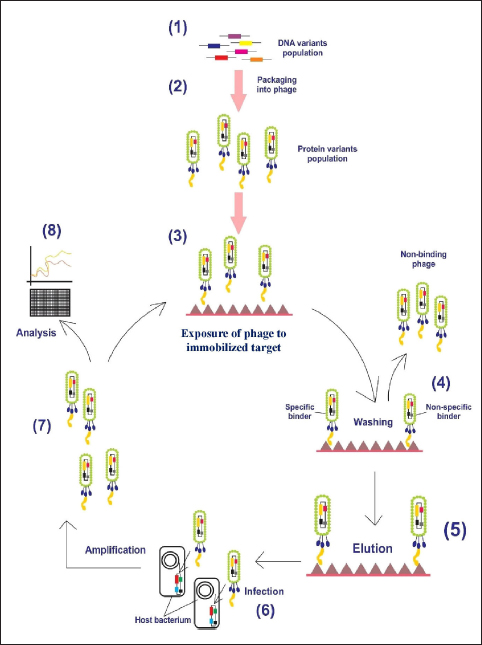
mAbs are versatile biomacromolecules that have high specificity for binding to various types of protein and non-protein substrates [5-7]. These mAbs can be produced using numerous approaches to enhance their functioning and utility [Figure 1] [8]. To date, there are currently over 130 therapeutically approved mAbs, with numerous others undergoing preclinical and clinical development [9]. Hybridoma technology is a widely employed technique for the production of mAbs. During this procedure, B lymphocytes that produce antibodies are separated from mice that have been immunized with a specific antigen. These B lymphocytes are then combined with immortal myeloma cell lines to create hybrid cells known as hybridoma cell lines. The hybridoma cells are cultivated in a controlled environment to generate mAbs that target a particular antigen.
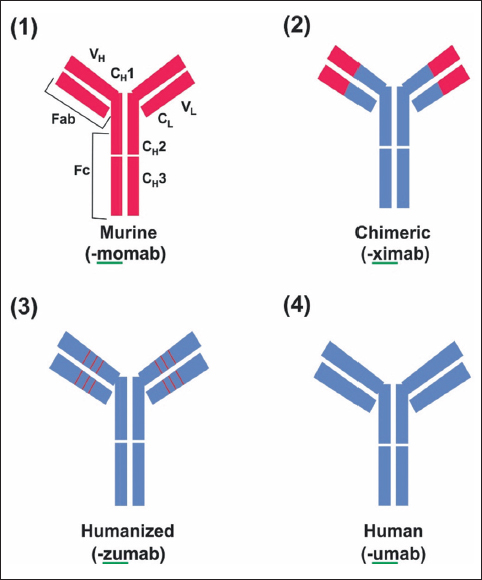 | Figure 1: Structure of mAb and modification of murine mAb in therapy. (1) The murine monoclonal antibody. (2) Chimeric monoclonal antibody has variable regions of murine origin and the rest is of human origin. (3) Humanized monoclonal antibody has a hypervariable region of murine origin and the rest is of human origin. (4) Fully human monoclonal antibody. [Click here to view] |
mAbs in the market developed by hybridoma technology are given in Table 1. The human mAbs product developed by phage display, transgenic mice, and recombinant technology is given in Table 2. The top best-selling mAb drugs in the year 2022 are given in Table 3.
Table 1: US FDA-approved monoclonal antibody in the market developed by hybridoma technology.
| S. No. | Antibody | Brand name | Year of approval | Target | Format | Indication | Company | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Muromonab-CD3 | Orthoclone OKT3 | 1986 | CD3 | Murine IgG2 | Kidney transplant rejection | Centocor Ortho Biotech Products L.P. | [10] |
| 2 | Abciximab | Reopro | 1994 | GPIIb/IIIa | Chimeric IgG1 Fab | Prevention of blood clots in angioplasty | Centocor Inc. | [10] |
| 3 | Rituximab | MabThera, Rituxan | 1997 | CD20 | Chimeric IgG1 | Non-Hodgkin lymphoma | Biogen Inc. | [10] |
| 4 | Palivizumab | Synagis | 1998 | RSV | Humanized IgG1 | Prevention of respiratory syncytial virus infection | MedImmune | [10] |
| 5 | Infliximab | Remicade | 1998 | TNFα | Chimeric IgG1 | Crohn’s disease | Janssen Biotech Inc. | [10] |
| 6 | Trastuzumab | Herceptin | 1998 | HER2 | Humanized IgG1 | Breast cancer | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 7 | Alemtuzumab | Campath, Lemtrada | 2001 | CD52 | Humanized IgG1 | Chronic myeloid leukemia | Berlex Inc. | [10] |
| 8 | Ibritumomab tiuxetan | Zevalin | 2002 | CD20 | Murine IgG1 | Non-Hodgkin lymphoma | Biogen Inc. | [10] |
| 9 | Omalizumab | Xolair | 2003 | IgE | Humanized IgG1 | Asthma | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 10 | Cetuximab | Erbitux | 2004 | EGFR | Chimeric IgG1 | Colorectal cancer | Bristol-Myers Squibb | [10] |
| 11 | Bevacizumab | Avastin | 2004 | VEGF-A | Humanized IgG1 | Colorectal cancer | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 12 | Natalizumab | Tysabri | 2004 | ITGA4 | Humanized IgG4 | Multiple sclerosis | Biogen Inc. | [10] |
| 13 | Ranibizumab | Lucentis | 2006 | VEGF-A | Humanized IgG1 Fab | Macular degeneration | Roche, F. Hoffmann-La Roche Ltd. | [10] |
| 14 | Eculizumab | Soliris | 2007 | C5 | Humanized IgG2/4 | Paroxysmal nocturnal hemoglobinuria | Alexion Pharmaceuticals Inc. | [10] |
| 15 | Certolizumab pegol | Cimzia | 2008 | TNFα | Humanized Fab, pegylated | Crohn’s disease | Celltech, UCB. | [10] |
| 16 | Tocilizumab | RoActemra, Actemra | 2010 | IL-6R | Humanized IgG1 | Rheumatoid arthritis | Chugai Pharmaceutical Co. Ltd. | [10] |
| 17 | Brentuximab vedotin | Adcetris | 2011 | CD30 | Chimeric IgG1 | Hodgkin lymphoma | Seattle Genetics Inc. | [10] |
| 18 | Pertuzumab | Perjeta | 2012 | HER2 | Humanized IgG1 | Breast Cancer | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 19 | Trastuzumab emtansine | Kadcyla | 2012 | HER2 | Humanized IgG1 | Breast cancer | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 20 | Obinutuzumab | Gazyva, Gazyvaro | 2013 | CD20 | Humanized IgG1 | Chronic lymphocytic leukemia | Biogen Inc. | [10] |
| 21 | Siltuximab | Sylvant | 2014 | IL-6 | Chimeric IgG1 | Castleman disease | Centocor Inc. | [10] |
| 22 | Vedolizumab | Entyvio | 2014 | α4β7 integrin | Humanized IgG1 | Ulcerative colitis, Crohn disease | Genentech Inc. | [10] |
| 23 | Blinatumomab | Blincyto | 2014 | CD19, CD3 | Murine bispecific tandem scFv | Acute lymphoblastic leukemia | Amgen | [10] |
| 24 | Pembrolizumab | Keytruda | 2014 | PD-1 | Humanized IgG4 | Melanoma | Merck & Co. Inc. | [10] |
| 25 | Idarucizumab | Praxbind | 2015 | Dabigatran | Humanized Fab | Reversal of dabigatran-induced anticoagulation | BIP | [10] |
| 26 | Dinutuximab | Unituxin | 2015 | GD2 | Chimeric IgG1 | Neuroblastoma | United Therapeutics Corp. | [10] |
| 27 | Mepolizumab | Nucala | 2015 | IL-5 | Humanized IgG1 | Severe eosinophilic asthma | Centocor Inc. | [10] |
| 28 | Elotuzumab | Empliciti | 2015 | SLAMF7 | Humanized IgG1 | Multiple myeloma | Bristol-Myers Squibb | [10] |
| 29 | Ixekizumab | Taltz | 2016 | IL-17α | Humanized IgG4 | Psoriasis | Eli Lilly | [10] |
| 30 | Reslizumab | Cinqaero, Cinqair | 2016 | IL-5 | Humanized IgG4 | Asthma | Celltech, UCB | [10] |
| 31 | Atezolizumab | Tecentriq | 2016 | PD-L1 | Humanized IgG1 | Bladder cancer | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 32 | Obiltoxaximab | Anthim | 2016 | Bacillus anthrasis PA | Chimeric IgG1 | Prevention of Inhalational anthrax | Elusys Therapeutics Inc. | [10] |
| 33 | Inotuzumab ozogamicin | Besponsa | 2017 | CD22 | Humanized IgG4 | Acute lymphoblastic leukemia | Wyeth Pharmaceuticals | [10] |
| 34 | Ocrelizumab | Ocrevus | 2017 | CD20 | Humanized IgG1 | Multiple sclerosis | Biogen Inc. | [10] |
| 35 | Emicizumab | Hemlibra | 2017 | Factor IXa, X | Humanized IgG4, bispecific | Hemophilia A | Chugai Pharmaceutical Co., Ltd. | [10] |
| 36 | Benralizumab | Fasenra | 2017 | IL-5Rα | Humanized IgG1 | Asthma | MedImmune | [10] |
| 37 | Gemtuzumab ozogamicin | Mylotarg | 2017 | CD33 | Humanized IgG4; ADC | Acute myeloid leukemia | Pfizer | [10] |
| 38 | Mogamulizumab | Poteligeo | 2018 | CCR4 | Humanized IgG1 | Mycosis fungoides or Sézary syndrome | Kyowa Hakko Kirin | [10] |
| 39 | Galcanezumab | Emgality | 2018 | CGRP | Humanized IgG4 | Migraine prevention | Eli Lilly | [10] |
| 40 | Tildrakizumab | Ilumya | 2018 | IL-23 p19 | Humanized IgG1 | Plaque psoriasis | Merck & Co. Inc. | [10] |
| 41 | Fremanezumab | Ajovy | 2018 | CGRP | Humanized IgG2 | Migraine prevention | Teva Pharmaceutical Industries, Ltd. | [10] |
| 42 | Ibalizumab | Trogarzo | 2018 | CD4 | Humanized IgG4 | HIV infection | Taimed Biologics Inc. | [10] |
| 43 | Ravulizumab | Ultomiris | 2018 | C5 | humanized IgG2/4 | Paroxysmal nocturnal hemoglobinuria | Alexion Pharmaceuticals Inc. | [10] |
| 44 | Caplacizumab | Cablivi | 2019 | von Willebrand factor | Humanized Nanobody | Acquired thrombotic thrombocytopenic purpura | Ablynx | [10] |
| 45 | Romosozumab | Evenity | 2019 | Sclerostin | Humanized IgG2 | Osteoporosis in | Amgen | [10] |
| 46 | Risankizumab | Skyrizi | 2019 | IL-23 p19 | Humanized IgG1 | Plaque psoriasis | Boehringer Ingelheim Pharmaceuticals | [10] |
| 47 | Polatuzumab vedotin | Polivy | 2019 | CD79β | Humanized IgG1 ADC | Diffuse large B-cell lymphoma | Roche, F. Hoffmann-La Roche, Ltd. | [10] |
| 48 | Brolucizumab | Beovu | 2019 | VEGF-A | Humanized scFv | Macular degeneration | Novartis Pharmaceuticals Corp. | [10] |
| 49 | Crizanlizumab | Adakveo | 2019 | P-selectin | Humanized IgG2 | Sickle cell disease | Novartis Pharmaceuticals Corp. | [10] |
| 50 | Eptinezumab-jjmr | Vyepti | 2020 | CGRP block | Humanized | Migraine | Lundbeck | [11] |
| 51 | Isatuximab-irfc | Sarclisa | 2020 | Anti-CD38 | Chimeric | Multiple myeloma | SANOFI AVENTIS | [11] |
| 52 | Sacituzumab govitecan-hziy | Trodelvy | 2020 | Anti-Trop-2; SN-38; | Humanized | mTNBC | IMMUNOMEDICS | [11] |
| 53 | Daratumumab and hyaluronidase-fihj | Darzalex Faspro | 2020 | Anti-CD38, | Human | Multiple myeloma | JANSSEN | [11] |
| 54 | Inebilizumab-cdon | Uplizna | 2020 | Anti-CD19 | Humanized | NMOSD (AQP4+) | VIELA | [11] |
| 55 | Pertuzumab, trastuzumab, and hyaluronidase-zzxf | Phesgo | 2020 | Anti-HER2; | Humanized | Breast cancer (HER2+) | GENENTECH | [11] |
| 56 | Belantamab mafodotin-blm | Blenrep | 2020 | Anti-BCMA | Humanized | Multiple Myeloma | GLAXOSMITHKLINE | [11] |
| 57 | Satralizumab-mwge | Enspryng | 2020 | Anti-IL6 receptor | Humanized | NMOSD (AQP4+) | GENENTECH | [11] |
| 58 | ATOLTIVIMAB +2 | Inmazeb | 2020 | Zaire ebolavirus glycoprotein | Humanized IgG1κ | Zaire ebolavirus infection. | REGENERON | [11] |
| 59 | NAXITAMAB-GQGK | Danyelza | 2020 | Anti-glycolipid GD2 | Humanized IgG1 | Neuroblastoma | Y-MABS THERAPEUTICS | [11] |
| 60 | MARGETUXIMAB-CMKB | Margenza | 2020 | Anti-HER2 | chimeric IgG | Breast cancer | MACROGENICS | [11] |
| 61 | RITUXIMAB-ARRX | Riabni | 2020 | Anti-CD20 | chimeric IgG | Non Hodgkin lymphoma | AMGEN | [11] |
| 62 | Dostarlimab-gxly | Jemperli | 2021 | PD-1 | Humanized | Endometrial cancer | GSK | [12] |
| 63 | Loncastuximab tesirine-lpyl | Zynlonta | 2021 | CD19 | Humanized IgG1 | B cell lymphoma | ADC | [12] |
| 64 | Dostarlimab-gxly | Jemperli | 2021 | PD-1 | Humanized | Solid tumor | GSK | [12] |
| 65 | Ranibizumab-nuna | Byooviz | 2021 | VEGF | Humanized IgG1 | Macular degeneration | SAMSUNG | [12] |
| 66 | Ranibizumab | Susvimo | 2021 | VEGF | Humanized IgG1 | Macular degeneration | GENEN TECH | [12] |
| 67 | Faricimab-svoa | Vabysmo | 2022 | VEGF; Ang-2 | Humanized IgG1 | nAMD; DME | GENENTECH | [13] |
| 68 | Sutimlimab-jome | Enjaymo | 2022 | CP | Humanized | Cold agglutinin disease | BIOVERATIV | [13] |
| 69 | Bevacizumab-maly | Alymsys | 2022 | VEGF | Humanized IgG1 | mCRC | AMNEAL | [13] |
| 70 | Risankizumabrzaa | Skyrizi | 2022 | IL-23 | Humanized IgG1 | Plaque psoriasis | ABBVIE | [13] |
| 71 | Ranibizumab-eqr | Cimerli | 2022 | VEGF | Humanized IgG1 | nAMD; RVO; DME; | COHERUS | [13] |
| 72 | Spesolimab-sbzo | Spevigo | 2022 | IL-36R | Humanized IgG1 | Pustular psoriasis | BOEHRINGER | [13] |
| 73 | Bevacizumab-adc | Vegzelma | 2022 | VEGF | Humanized IgG1 | mCRC; NSCLC; | CELLTRION | [13] |
| 74 | Mirvetuximab soravtansine-gynx | Elahere | 2022 | FRα-DM4 | Humanized IgG1 | Ovarian cancer | IMMUNOGEN | [13] |
| 75 | Teplizumab-mzwv | Tzield | 2022 | CD3 | Humanized IgG1 | type 1 diabetes | PROVENTION | [13] |
| 76 | Osunetuzumab-axgb | Lunsumio | 2022 | CD20; CD3 | Humanized IgG1 | Follicular lymphoma | GENENTECH | [13] |
| 77 | Ublituximab-xiiy | Briumv | 2022 | CD20 | Chimeric IgG1 | Multiple sclerosis | TG | [13] |
| 78 | Teclistamab-cqyv | Tecvayli | 2022 | BCMA; CD3 | Humanized bispecific IgG4 | Multiple myeloma | JANSSEN | [13] |
| 79 | Elranatamab | Elrexfio | 2023 | BCMA, CD3 | Humanized IgG2 | Multiple myeloma | Pfizer | [10] |
| 80 | Rozanolixizumab | Rystiggo | 2023 | FcRn; | Humanized IgG4 | Generalized myasthenia gravis | UCB | [10] |
| 81 | Talquetamab | Talvey | 2023 | G protein-coupled receptor 5D, CD3; | Humanized IgG4 bispecific | Multiple myeloma | Janssen | [10] |
| 82 | Epcoritamab | Epkinly | 2023 | CD20, CD3 | Bispecific humanized IgG1 | Diffuse large B cell lymphoma | Abbvie | [10] |
| 83 | Glofitamab | Columvi | 2023 | CD20, CD3e | Bispecific | Diffuse large B-cell lymphoma | Genentech | [10] |
| 84 | Lecanemab | Leqembi | 2023 | Amyloid beta protofibrils; | Humanized IgG1 | Alzheimer’s disease | Biogen | [10] |
| 85 | Retifanlimab | Zynyz | 2023 | PD-1 | Humanized IgG4 | Merkel cell carcinoma | Incyte | [10] |
Table 2: US FDA-approved human monoclonal antibodies in the market.
| S. No. | Antibody | Brand name | Year of approval | Target | Format | Technology | Indication | Company | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Adalimumab | Humira | 2002 | TNFα | Human IgG1 | Phage display | Rheumatoid arthritis | AbbVie Inc. | [10] |
| 2 | Panitumumab | Vectibix | 2006 | EGFR | Human IgG2 | Transgenic mice | Colorectal cancer | Amgen | [10] |
| 3 | Ustekinumab | Stelara | 2009 | IL-12/23 | Human IgG1 | Transgenic mice | Psoriasis | Centocor Ortho Biotech Inc. | [10] |
| 4 | Canakinumab | Ilaris | 2009 | IL-1β | Human IgG1 | Transgenic mice | Muckle-Wells syndrome | Novartis Pharmaceuticals Corp. | [10] |
| 5 | Golimumab | Simponi | 2009 | TNFα | Human IgG1 | Transgenic mice | Rheumatoid and psoriatic arthritis, ankylosing spondylitis | Centocor Ortho Biotech Inc. | [10] |
| 6 | Ofatumumab | Arzerra | 2009 | CD20 | Human IgG1 | Transgenic mice | Chronic lymphocytic leukemia | Genmab A/S | [10] |
| 7 | Denosumab | Xgeva, Prolia | 2010 | RANKL | Human IgG2 | Transgenic mice | Bone loss | Amgen | [10] |
| 8 | Belimumab | Benlysta | 2011 | BLyS | Human IgG1 | Phage display | Systemic lupus erythematosus | GlaxoSmithKline. | [10] |
| 9 | Ipilimumab | Yervoy | 2011 | CTLA-4 | Human IgG1 | Transgenic | Metastatic melanoma | Bristol-Myers Squibb | [10] |
| 10 | Raxibacumab | Abthrax | 2012 | B. anthrasis PA | Human IgG1 | Transgenic mice | Anthrax infection | GlaxoSmithKline | [10] |
| 11 | Ramucirumab | Cyramza | 2014 | VEGFR2 | Human IgG1 | Phage display | Gastric cancer | Eli Lilly | [10] |
| 12 | Nivolumab | Opdivo | 2014 | PD-1 | Human IgG4 | Transgenic mice | Melanoma, non-small cell lung cancer | Bristol-Myers Squibb | [10] |
| 13 | Necitumumab | Portrazza | 2015 | EGFR | Human IgG1 | Phage display | Non-small cell lung cancer | Eli Lilly | [10] |
| 14 | Secukinumab | Cosentyx | 2015 | IL-17α | Human IgG1 | Transgenic mice | Psoriasis | Novartis Pharmaceuticals Corp. | [10] |
| 15 | Alirocumab | Praluent | 2015 | PCSK9 | Human IgG1 | Transgenic mice | High cholesterol | Regeneron Pharmaceuticals Inc. | [10] |
| 16 | Evolocumab | Repatha | 2015 | PCSK9 | Human IgG2 | Transgenic mice | High cholesterol | Amgen | [10] |
| 17 | Daratumumab | Darzalex | 2015 | CD38 | Human IgG1 | Transgenic mice | Multiple myeloma | Genmab A/S | [10] |
| 18 | Olaratumab | Lartruvo | 2016 | PDGFRα | Human IgG1 | Transgenic mice | Soft tissue sarcoma | Eli Lilly | [10] |
| 19 | Bezlotoxumab | Zinplava | 2016 | Clostridium difficile enterotoxin B | Human IgG1 | Transgenic mice | Prevention of Clostridium difficile infection recurrence | Merck & Co. Inc. | [10] |
| 20 | Brodalumab | Siliq, Lumicef | 2017 | IL-17R | Human IgG2 | Transgenic mice | Plaque psoriasis | MedImmune | [10] |
| 21 | Guselkumab | Tremfya | 2017 | IL-23 p19 | Human IgG1 | Phage display | Plaque psoriasis | MorphoSys | [10] |
| 22 | Dupilumab | Dupixent | 2017 | IL-4Rα | Human IgG4 | Transgenic mice | Atopic dermatitis | Regeneron Pharmaceuticals Inc. | [10] |
| 23 | Sarilumab | Kevzara | 2017 | IL-6R | Human IgG1 | Transgenic mice | Rheumatoid arthritis | Regeneron Pharmaceuticals Inc. | [10] |
| 24 | Avelumab | Bavencio | 2017 | PD-L1 | Human IgG1 | Phage display | Merkel cell carcinoma | Merck | [10] |
| 25 | Durvalumab | Imfinzi | 2017 | PD-L1 | Human IgG1 | Transgenic mice | Bladder cancer | MedImmune | [10] |
| 26 | Burosumab | Crysvita | 2018 | FGF23 | Human IgG1 | Transgenic mice | X-linked hypophosphatemia | Kyowa Hakko Kirin/Ultragenyx | [10] |
| 27 | Lanadelumab | Takhzyro | 2018 | Plasma kallikrein | Human IgG1 | Phage display | Hereditary angioedema attacks | Dyax Corp. | [10] |
| 28 | Erenumab | Aimovig | 2018 | CGRPR | Human IgG2 | Transgenic mice | Migraine prevention | Novartis | [10] |
| 29 | Cemiplimab | Libtayo | 2018 | PD-1 | Human mAb | Transgenic mice | Cutaneous squamous cell carcinoma | Regeneron Pharmaceuticals Inc. | [10] |
| 30 | Emapalumab | Gamifant | 2018 | IFNγ | Human IgG1 | Phage display | Primary hemophagocytic lymphohistiocytosis | NovImmmune | [10] |
| 31 | Moxetumomab pasudodox | Lumoxiti | 2018 | CD22 | Murine IgG1 dsFv | Phage display | Hairy cell leukemia | MedImmune | [10] |
| 32 | Teprotumumab-trb | Tepezz | 2020 | IGF-1R block | Human IgG1 | Transgenic mice | Thyroid eye disease | Horizon therapeutics | [11] |
| 33 | Evinacu-mab-dgnb | Evkeeza | 2021 | ANGPTL3 | Human | Recombinant | HoFH LDL-C | Regeneron | [12] |
| 34 | Amivantamab-vmjw | Rybrevan | 2021 | EGFR MET Receptor | Human IgG1 | Recombinant | NSCLC | Janssen | [12] |
| 35 | Aducanumab-avwa | Aduhelm | 2021 | Amyloid beta | Human IgG1 | Recombinant | Alzheimer’s disease | Biogen | [12] |
| 36 | Anifro-lumab-fnia | Saphnelo | 2021 | (I)IFNR | Human | Recombinant | SLE | Astrazeneca | [12] |
| 37 | Tisotumab vedotin-tftv | Tivdak | 2021 | Tissue Facto | Human | Recombinant | Cervical | Seagen | [12] |
| 38 | Adalimumab-aqvh | Yusimry | 2021 | TNF | Human IgG1 | Recombinant | RA | Coherus | [12] |
| 39 | Tezepelumab-ekko | Tezspire | 2021 | TSLP | Human | Recombinant | Severe asthma | Astra zeneca | [12] |
| 40 | Tralokinumab-ldrm | adbry | 2021 | IL-13 | Human IgG4 | Recombinant | Eczema | Leo | [12] |
| 41 | Nivolumab; relatlimabrmbw | Opdualag | 2022 | PD-1; LAG-3 | Human IgG4 | Recombinant | melanoma | Bms | [13] |
| 42 | Tremelimumabactl | Imjudo bla 761289 | 2022 | CTLA-4 | Human IgG2 | Recombinant | Liver cancer | Astra Zeneca | [13] |
| 43 | Tremelimumabactl | Imjudo bla 761270 | 2022 | CTLA-4 | Human IgG2 | Recombinant | Liver cancer | Astra Zeneca | [13] |
| 44 | Dalimumab-aacf | Idacio | 2022 | TNF | Human IgG1 | Recombinant | RA JIA PsA | Fresenius | [13] |
| 45 | Pozelimab | Veopoz | 2023 | Complement 5 | Human IgG4 | Recombinant | CHAPLE disease | Regeneron | [10] |
| 46 | Nirsevimab | Beyfortus | 2023 | RSV | Human IgG1 | Recombinant | RSV infection | Sanofi | [10] |
Table 3: Top five best-selling monoclonal antibody drugs in 2022.
| S. No. | Antibody | Brand name | Company | Target | Technology/ formats | Indication | Revenue (USD) | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Adalimumab | Humira | AbbVie | TNFα | Phage display | Rheumatoid arthritis | 21.2 billion | [14] |
| 2 | Pembrolizumab | Keytruda | Merck & Co. | PD-1 | Humanized IgG4 | Melanoma | 20.9 billion | [14] |
| 3 | Ustekinumab | Stelara | Janssen | IL-12/23 | Human IgG1 | Plaque psoriasis, psoriatic arthritis | 9.7 billion | [14] |
| 4 | Dupilumab | Dupixent | Sanofi Genzyme, Regeneron | IL-4Rα | Human IgG4 | Atopic dermatitis, asthma, chronic rhinosinusitis with nasal polyps | 8.7 billion | [14] |
| 5 | Nivolumab | Opdivo | Bristol-Myers Squibb | PD-1 | Transgenic mice | Melanoma, non-small cell lung cancer | 8.2 billion | [14] |
mAb drugs have developed from clinical research to commercialization over the past few decades. In the past few years, the total number of antibody drugs approved for launch has proliferated, with 130 approved and available on the market [15]. mAb treatments were the fastest-growing category in the worldwide biopharmaceutical market in 2022, with four of the top ten best-selling biopharmaceutical products. Keytruda, which is a drug developed by Merck and introduced in 2014, garnered USD 20.9 billion in sales in 2022 [16]. In recent years, the global mAb market has grown, reaching USD 178.5 billion in 2021, which is an increase of 12% year over year. As of April 2022, there were 250 mAb therapies in Phase III clinical trials around the world [17]. Antibody medications are expected to be licensed and marketed in great numbers in the next few years, with a market value of USD 420–460 billion in 2028. The global mAb market has practically been split up by Roche, Johnson & Johnson, Merck, Novartis, AbbVie, and Amgen. The top five antibody drugs in the world by sales in 2022 were Humira, Keytruda, Stelara, Dupixent, and Opdivo, with a combined share of more than 50% of the global market [1]. Therefore, this review article discusses the rapid growth of therapeutic mAb products as well as the technological developments in producing these antibodies using various in vitro methods. Furthermore, the study offers a thorough synopsis of the approved mAbs currently accessible on the market, together with their distinct targets, structures, and permissible uses.