
1. INTRODUCTION
Cocoa (Theobroma cacao L.) is a tropical species native to the Amazon Basin of great economic importance to produce cocoa beans, the basis of the global chocolate industry [1,2]. Annual cocoa production represents more than US$10 billion in income for numerous farming families around the world, producing approximately 4.5 million tons of cocoa per year [3]. Peru ranks ninth among the world’s largest cocoa producers and third in the Americas, with approximately 160,000 tons of cocoa per year in the past 2 years [4]. Peru is among the world’s leading producers and suppliers of fine aroma and organic cocoa [5]. National production is led by the San Martin region with 60,000 tons of cocoa, accounting for 38% of the country’s total production [6].
The demand for chocolate increases annually, but it is estimated that there will be a shortage of raw materials due to the spread of diseases that can lead to losses from 20% to the entire production of cocoa [7]. In this regard, traditional propagation methodologies have been developed, such as grafting and rooting of cocoa plant cuttings with the best agronomic characteristics such as resistance to diseases, higher production, and seed quality. These methodologies have certain disadvantages associated with the low rate of propagation and the undesirable bushy growth pattern [8]. Alternatives such as tissue culture, through somatic embryogenesis, allow the clonal propagation of superior cocoa genotypes with the same genetic backgrounds. This biotechnological tool was employed in a wide variety of cocoa genotypes [9,10].
Somatic embryogenesis relies on the totipotency of plant somatic cells to express cellular reprogramming [11], which has the potential to largely produce numerous individuals from a single explant and this is applied in different species and genotypes [12]. This morphogenetic pathway is developed by different biochemical and molecular events that encompass the stages of induction, expression, maturation, and germination [13,14]. Likewise, through somatic embryo induction, high propagation rates of material of genotypes of interest can be obtained while maintaining genetic stability, allowing its application for mass propagation of species with agronomic potential, conservation of genetic resources [15], application in genetic transformation [16,17] and gene editing [18].
Primary somatic embryos (PSE) formed from initial explants can be used for the formation of secondary somatic embryos (SSE) [19,20]. When opposed to primary somatic embryogenesis, secondary somatic embryogenesis has the advantage of the formation of a higher number of somatic embryos and repeatability [14,21]. In addition, numerous cycles of secondary embryogenesis can maintain the embryonic stage for long periods [22].
The osmotic potential of the culture medium can affect the development and percentage of somatic embryo formation [23]. As a solution, high molecular mass osmotic agents such as polyethylene glycol (PEG) and polyols (D-Mannitol and D-Sorbitol), which could reduce the osmotic potential in the culture medium, have been used. Thus, the application of these osmotic agents has increased the rate of somatic embryo multiplication in different species [24-26].
Therefore, the objective of the research was to determine the effect of three osmoregulatory (PEG, D-Mannitol, and D-Sorbitol) on the production of cocoa SSE of a highly productive elite genotype.
2. MATERIAL AND METHODS
2.1. Vegetal Material
Immature flower buds were obtained from the I52 genotype, which is known for producing the highest quality chocolate among the cocoa collection at the Instituto de Investigación para el Desarrollo Sustentable de Ceja de Selva (INDES-CES) in Amazonas, Peru. Flower buds 6–8 mm long, immature in appearance, were obtained from healthy plants with vigorous growth. The flower buds were transported to the Laboratorio de Fisiología y Biotecnología Vegetal at the INDES-CES of the Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas (UNTRM-A).
2.2. Indirect Primary Somatic Embryogenesis
The immature flower buds were soaked in a 1% calcium hypochlorite solution for 25 min and then rinsed 3 times with sterile distilled water in a laminar flow chamber. Cuts of 1/3 length from the base of the flower were made to extract the staminodes. Staminodes were transferred to Petri dishes containing culture media and sealed with parafilm to avoid contamination, plates were incubated in total darkness at a temperature of 25 ± 2°C. The composition of the culture media was based on DKW medium [27], which included DKW 10× macronutrient stock solutions A and B, DKW 100× micronutrient solution, DKW 1000× vitamin solution and DKW 1000× amino acid solution. In addition, the culture media were supplemented with salts from woody plant medium [28], Gamborg’s vitamin solution [29], and growth regulators such as thidiazuron, 6-benzylaminopurine (BAP) and 2,4-dichlorophenoxyacetic acid. The culture medium used for the staminodes culture initially consisted of primary callus growth medium followed by secondary callus growth medium (SCG), the Embryo development 4 (ED4) medium, and subcultured 6 times in the Embryo development 3 (ED3) medium, with intervals of 14 days between each medium [14]. Incubation was carried out in the absence of light at 25 ± 2°C. At the end of the process, the somatic embryos reached maturity. The preparation of all culture media used is indicated in Table 1.
Table 1: Composition of culture media for cocoa somatic embryogenesis.
| Component | Culture media for cocoa somatic embryogenesis | ||||
|---|---|---|---|---|---|
| PCG | SCG | ED4 | ED3 | EDL | |
| DKW macronutrient solution A (mL/L) | 100 | - | 100 | 100 | 100 |
| DKW macronutrient solution B (mL/L) | 100 | - | 100 | 100 | 100 |
| DKW micronutrient solution (mL/L) | 10 | - | 10 | 10 | 10 |
| DKW vitamin solution (mL/L) | 1 | - | 1 | 1 | 1 |
| DKW amino acid solution (mL/L) | - | - | - | - | 1 |
| WPM salt (g/L) | - | 2.3 | - | - | - |
| Gamborg’s vitamin solution (mL/L) | - | 1 | - | - | - |
| Glucose (g/L) | 20 | 20 | - | - | 20 |
| Sucrose (g/L) | - | - | 40 | 30 | - |
| Glutamine (mg/L) | 250 | - | - | - | - |
| Myoinositol (mg/L) | 200 | - | - | - | - |
| 2,4-D (mg/L) | 2 | 2 | - | - | - |
| TDZ (μg/L) | 5 | - | - | - | - |
| BAP (mg/L) | - | 0.05 | - | - | - |
| Potassium nitrate (g/L) | - | - | - | - | 0.3 |
| Phytagel (g/L) | 2 | 2.2 | 2 | 2 | 1.8 |
WPM: Woody plant medium, TDZ: Thidiazuron, BAP: 6-benzylaminopurine, 2,4-D: 2,4-dichlorophenoxyacetic, PCG: Primary callus growth, ED4: Embryo development 4, SCG: Secondary callus growth.
2.3. Indirect Secondary Somatic Embryogenesis
Primary embryo epicotyls were collected in the torpedo and cotyledonal stages of 10–40 mg and transverse cuts of 2 mm were made before seeding in a Petri dish containing SCG medium. The explants were transferred to Petri dishes with ED4 medium containing the osmoregulators PEG 6000, D-Mannitol or D-Sorbitol at concentrations of 0%, 1%, 3% and 5%. The explants were then subcultured in ED3 medium containing the same concentrations of the respective osmoregulators. Subculture in this latter medium was performed 6 times (at 14-day intervals) until the somatic embryos reached maturity. The entire process of indirect secondary somatic embryogenesis was carried out with incubation conditions in total darkness, at 25 ± 2°C. Finally, the embryos formed were transferred to Embryo germination and conversion (EDL) medium [Table 1].
2.4. Statistical Analysis
The experiment was established by four independent replicates composed of three experimental units (each experimental unit represented a Petri dish with 25 embryo segments with induced secondary embryogenesis) per treatment. To determine the formation of secondary embryos in each treatment, the frequency of callus and embryo formation was recorded in six subcultures in the ED3 medium. The effect of the osmoregulators was evaluated on six occasions, the first five evaluations were carried out every 7 days after sowing (DAS) and the sixth evaluation was carried out at 63 das. All generated data were subjected to an analysis of variance (ANOVA) using the generalized linear model (GLM) and means comparison by treatment using the Tukey test (P ≤ 0.05), and were analyzed with the R software (version 4.1.1).
3. RESULTS
3.1. The Effect of Osmotic Regulation
The highest number of SSE in cocoa was obtained with PEG 6000 at 1%. 39.8% of SSE was recorded in the first evaluation, as well as 53.5% at 14 DAS and 57.7% at 21 DAS, dropping to less than 20% from 35 DAS [Figure 1a]. The formation of heart embryos from globular embryos reached the highest frequency at 63 DAS with 19% [Figure 1b]. The evolution of the embryos from the heart state to the torpedo state reached 22.8% at 63 DAS [Figure 1c]. Embryos in the cotyledonary stage reached 165.4% at 63 DAS [Figure 1d].
 | Figure 1: Frequency of formation of secondary somatic embryos with polyethylene glycol 6000 in six different periods of time. Somatic embryos in stage: Globular (a), heart (b), torpedo (c) and cotyledonal (d). [Click here to view] |
The addition of D-Mannitol at a concentration of 5% was notable with the generation of 11.8% globular embryos on the 7th day. However, this type of embryos disappeared at 14 DAS and, in general, this was the treatment with the lowest frequency of embryos [Figure 2a]. The highest frequency of formation of heart stage embryos was obtained in the absence of D-Mannitol with a maximum frequency of 6.8% at 49 DAS [Figure 2b].
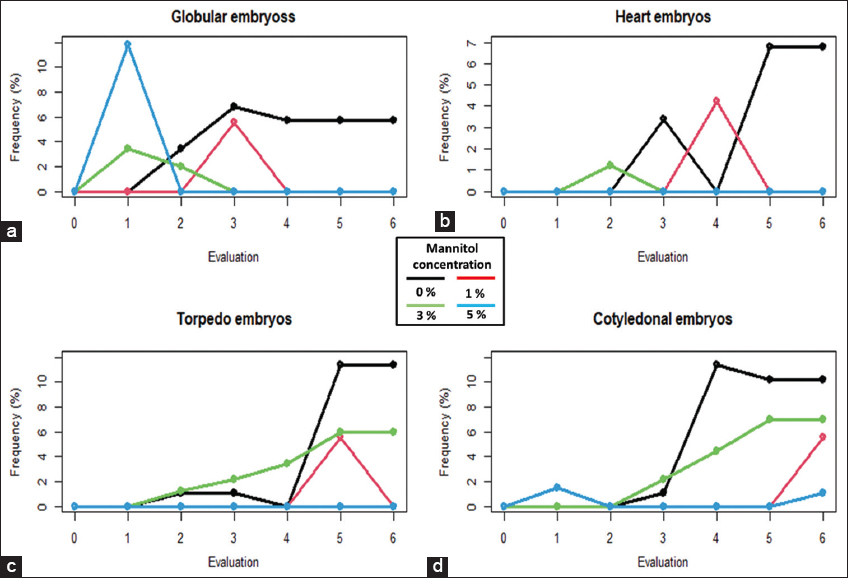 | Figure 2: Frequency of the formation of secondary somatic embryos with D-mannitol in six different periods of time. Somatic embryos in stage: Globular (a), heart (b), torpedo (c) and cotyledonal (d). [Click here to view] |
The absence of D-Mannitol in the culture medium allowed higher frequencies of embryo formation with 11.4% in the torpedo stage at 49 DAS [Figure 2c] and 11.4% in the cotyledonary stage at 35 DAS [Figure 2d]. From the last cotyledonary stage, it is possible to generate complete plants.
The 1% sorbitol allowed the largest number of embryos compared to the other concentrations and the absence of this osmoregulator. Embryos in the globular, heart, torpedo and cotyledonary stages reached frequencies of 17.2% in the first 7 days [Figure 3a], 14.7% at 21 DAS [Figure 3b], 21% at 49 DAS [Figure 3c], and 57.9% frequency of embryo formation at 63 DAS [Figure 3d], respectively.
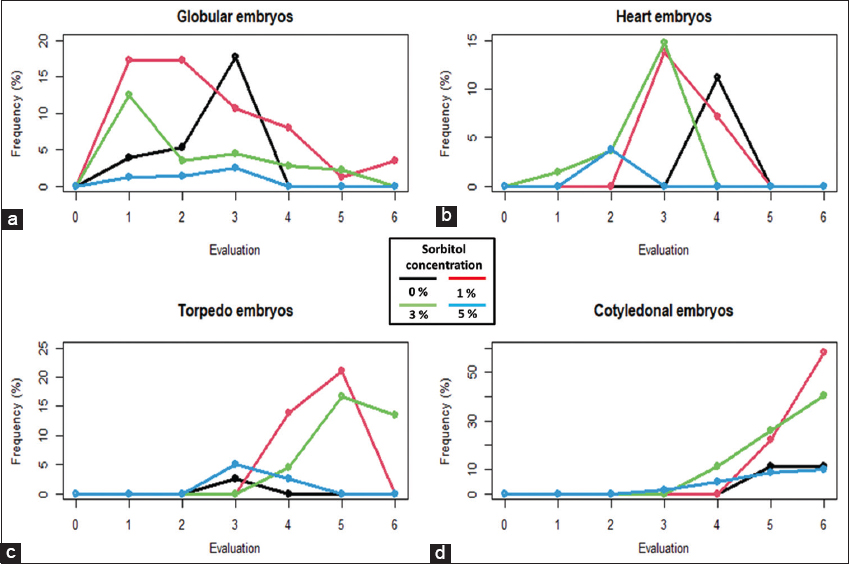 | Figure 3: Frequency of the formation of secondary somatic embryos with sorbitol at six different periods of time. Somatic embryos in stage; globular (a), heart (b), torpedo (c) and cotyledonal (d). [Click here to view] |
3.2. Number of Somatic Embryos by Development Stage
Callus formation in the SCG medium occurred in the first 14 days, obtaining a good percentage of embryos in the torpedo and cotyledonal stage, which gave rise to globular embryos from the 7th day after their introduction in the ED3 medium. The degree of development of globular somatic embryos varied between treatments. The total number of resulting somatic embryos was affected by a significant interaction of each osmoregulator and its concentration. Data showed that in the presence of PEG 6000 at a concentration of 1%, a greater development of globular and cotyledonal somatic embryos was produced, with means of 30.33 and 55.2, respectively [Table 2].
Table 2: Effect of osmoregulators and their concentration on the number of secondary somatic embryos in cocoa by degree of development.
| Osmoregulators concentration (%) | Embryogenic callus | Number of somatic embryos per explant | |||||
|---|---|---|---|---|---|---|---|
| PEG 6000 | D-Mannitol | D-Sorbitol | Globular | Heart | Torpedo | Cotyledonal | |
| 0 | 0 | 0 | 2.00d | 0.55b | 0.23c | 0.45d | 3.63c |
| 1 | 0 | 0 | 22.65a | 30.33a | 5.70a | 8.40a | 55.2a |
| 3 | 0 | 0 | 4.68c | 3.98b | 0.35c | 4.13c | 10.80b |
| 5 | 0 | 0 | 5.00c | 5.78b | 2.05a | 3.58c | 11.03b |
| 0 | 1 | 0 | 1.18d | 0.92b | 0.70c | 0.92d | 0.92b |
| 0 | 3 | 0 | 4.18c | 0.90b | 0.20c | 3.13c | 3.40c |
| 0 | 5 | 0 | 0.35d | 1.95b | 0.00d | 0.00e | 0.45 d |
| 0 | 0 | 1 | 4.83c | 9.57b | 3.48b | 5.80b | 13.38b |
| 0 | 0 | 3 | 9.33b | 4.23b | 3.28b | 5.75b | 12.90b |
| 0 | 0 | 5 | 3.50c | 0.82b | 0.63c | 1.25d | 4.15c |
Mean values in the column with diferent letters are signifcantly diferent (P≤0.05).
Treatment without the addition of osmoregulators showed a low formation of SSE [Figure 4a] compared to the other treatments. The addition of 3% D-Mannitol to the culture medium allowed the number of SSE in the torpedo stage to increase to 3.13 [Figure 4b]. The osmoregulator D-Sorbitol at 1% increased the number of somatic embryos in all stages, but the highest number was observed in the globular and cotyledonal stages [Figure 4c and d]. The osmoregulator that allowed the greatest formation of somatic embryos was PEG 6000 at 1%, with somatic embryos in a higher frequency of globular and cotyledonal stages [Figure 4e and g].
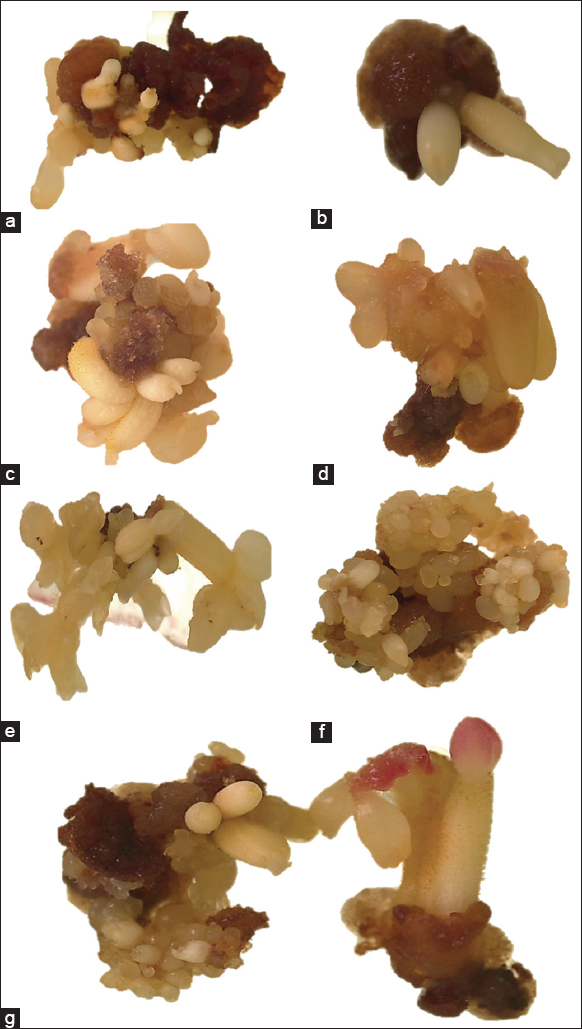 | Figure 4: Somatic embryos developed in a medium free of osmoregulators (a). Embryos in the heart and torpedo stage developed in a medium with 3% D-Mannitol (b). Development of embryos in the torpedo state with 1% D-Sorbitol (c). Embryos in cotyledonary stage with 1% D-Sorbitol (d). Somatic embryos predominantly in the cotyledonary state with polyethylene glycol (PEG) 6000 at 1% in the medium (e). Pro-duction of embryos with PEG 6000 in the medium, in which three stages are observed: globular, heart and torpedo (f). Embryos in mature cotyledonary stage developed in medium with 1% PEG 6000 (g). [Click here to view] |
4. DISCUSSION
Plant somatic cells have a high embryogenic capacity, but for this, it is necessary to induce cell reprogramming in vitro and obtain PSE. The develop embryogenic cells that continue to divide and allow the formation of SSE. From these embryogenic cells found in the outermost layers of developing embryos, new meristematic centers are formed [30]. The response to secondary embryogenesis can vary between species and even between cultivars, due to the presence of genotype dependence [10,21].
The induction of primary and SSE is a function of the supplementation of growth regulators and abiotic (osmotic) stress factors to which the ex-plants are exposed. These factors influence the initiation of signal expression leading to genetic reprogramming related to callus formation before somatic embryo induction [31]. The incubation of the explants under osmotic stress factors such as PEG 6000, D-Sorbitol, and D-Mannitol at low concentrations causes a greater accumulation of proline as an indicator of stress in the embryogenic cells. The accumulation of endogenous proline exerts a stimulating effect on somatic embryogenesis mainly through the cell wall glycoprotein that is related to morpho-regulatory functions [32,33].
Embryonic asynchrony is the main disadvantage of somatic embryogenesis when mass propagation of cocoa plants is intended [34,35]. To date, no research has been reported on the induction of secondary embryogenesis of cocoa primary embryo cells using osmoregulators. However, some studies evaluated the embryogenic response of cocoa explants, and showed that the expression time of secondary embryos is shorter than obtaining primary embryos and, apparently, there are no significant differences in terms of the response of globular explants and cotyledons [10,36].
The use of PEG has been reported in the induction of somatic embryos in Carica papaya L. Golden THB variety, exerting a positive effect by inducing 17.58 somatic embryos per callus using a concentration of 40 g/L of PEG, higher than those treatments without the addition of the osmoregulator. Likewise, the use of this osmoregulator positively influences the maturation of somatic embryos. At higher concentrations of the osmoregulator, there was a negative effect on the number of total somatic embryos, with differences of up to 31.85% in relation to the control [26]. This effect has been confirmed in the present study. At concentrations of 30 and 50 g/L of PEG 6000 in the medium, a reduction was observed in somatic embryos in the cotyledonary stage was observed. In a similar study in Phoenix dactylifera L. by increasing PEG concentrations in the culture medium, the growth rate of embryogenic callus and somatic embryos decreased, showing that the effect of this osmoregulator varies depending on embryonic development and the hormonal content of the culture medium [37].
The induction of somatic embryos has been optimized in Cajanus cajan L. by adding osmoregulators in the culture medium. Specifically, by exposing the embryogenic callus to dehydration stress imposed by PEG and osmotic stress created by mannitol, producing high-frequency somatic embryos, with 83 somatic embryos in the globular stage and 38 somatic embryos in the heart stage when using 4% PEG; while using 0.6 M mannitol, 87 somatic embryos in the globular stage and 28 somatic embryos in the heart stage [32].
The addition of the osmoregulator mannitol in the culture medium allowed the increase of the embryogenic rate in celery (Apium graveolens). By adding 3% and 4% mannitol, cell lysis is avoided and an increase in the number of somatic embryos obtained per explant is evidenced with the formation of 39.5% and 47.4% of somatic embryos, respectively. It is clear that the osmotic potential is an important factor for somatic embryogenesis, but not the only one, since the effect of mannitol could be manifested after a short exposure period (2–3 days), suggesting an embryogenic induction mechanism [38].
Negative effects have also been reported with the addition of mannitol in culture medium, as observed in studies carried out on Dioscorea spp., where it was determined that the use of different osmotic agents affects the in vitro survival of the species Dioscorea alata. In this sense, the use of mannitol in the culture medium of this species is not recommended, as it causes the death of all explants [39]. This fact was corroborated in our study, since the formation of embryos obtained was relatively low, with respect to the medium in the absence of this osmoregulator. Mannitol had a negative effect on Eucalyptus globulus cotyledons, inhibiting callus and somatic embryos by adding 36.44 g/L of mannitol to the culture medium [40]. This could indicate that the addition of mannitol in culture medium does not have a positive effect on the somatic embryogenesis of woody species. Likewise, mannitol can reduce the water absorption capacity of the culture medium, generating a reduction in water potential, much more than other osmoregulators such as sorbitol [41].
Sorbitol has the ability to generate less severe osmotic stress than other osmoregulators and is used as an energy source for explants [39]. For this reason, there are many investigations using sorbitol in somatic embryogenesis, except for cocoa, in which no investigations have been reported. In the case of avocado (Persea americana L.) good results were obtained using sorbitol, superior to those that can be obtained with PEG, with a maximum of 58.2% regeneration in medium with 2.5% sorbitol [42]. These results are very close to those obtained in our study, since an average frequency of 57.9% of cotyledonary embryos was reached at 63 days of induction of SSE.
The use of sorbitol has allowed to induce higher rates of SSE in Elaeis guineensis; by adding 0.2 M sorbitol, 22 somatic embryos per primary embryo have been obtained [24]. It has also been reported the increase in the formation of somatic embryos in Swietenia mahagoni with the addition of 20 and 40 g/L of sorbitol combined with 6 BAP to the culture medium, for which it was determined that sorbitol is one of the osmotic agents that influence the obtaining of somatic embryos and that the response to it can vary according to concentrations and genotypes [43].
5. CONCLUSION
In this work, the effect of the osmoregulators PEG 6000, D-Mannitol, and D-Sorbitol in the increase of SSE of cocoa was determined. The greatest response was achieved by adding PEG 6000 1% and D-Sorbitol 1% to the ED3 culture medium, obtaining a greater number of somatic embryos that completed the cotyledonal stage. The greatest formation of embryos was achieved with PEG 6000 1%, with 30.33 SSE in the globular stage and 55.2 SSE in the cotyledonal stage per explant.
6. ACKNOWLEDGMENTS
This study was financially supported by the Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica, (CONCYTEC, Peru) through its executing unit Programa Nacional de Estudios de Investigación Científica y Estudios Avanzados (PROCIENCIA) as part of the projects “Innovación biotecnológica para la producción masiva de embriones somáticos de cacao (T. cacao L.) fino de aroma en la Región Amazonas” (CONTRACT N° 033-2018-FONDECYT-BM-IADT-AV) and “Disminución de la absorción del cadmio en el cacao peruano mediante la edición genética de sus transportadores empleando la tecnología CRISPR-CAS9” (CONTRACT N° 26-2018-FONDECYT-BM-IADT-AV). In addition, financial support was provided by the UNTRM-A as part of the project “Creación de los servicios del centro de investigación, innovación y transferencia tecnológica de café de la Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas” (SNIP N°352439).
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to the conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All datasets were generated and analyzed in the present study.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Motamayor JC, Lachenaud P, Da Silva E Mota JW, Loor R, Kuhn DN, Brown JS, et al. Geographic and genetic population differentiation of the amazonian chocolate tree (Theobroma cacao L). PLoS One 2008;3:e3311. [CrossRef]
2. García L, Angulo Castro F, Hernández-Amasifuen AD, Corazon-Guivin MA, Alburquerque Vásquez J, Guerrero-Abad JC, et al. Global studies of cadmium in relation to Theobroma cacao:A bibliometric analysis from Scopus (1996-2020). Sci Agropecu 2021;12:611-22. [CrossRef]
3. International Cocoa Organization - Statistics. International Cocoa Organization. Available from:https://www.icco.org/statistics [Last accessed on 2023 Dec 02].
4. Food and Agriculture Organization. FAOSTAT. Available from:https://www.fao.org/faostat/en/#data/QCL [Last accessed on 2023 Dec 02].
5. Huamán O, Sánchez K, Armando C, Pari E, Acosta J, Galarreta L. Observatorio de Commodities:Cacao. Peru:Ministerio de Desarrollo Agrario y Riego;2023.
6. Huamán O, Sánchez K, Armando C, Pari E, Acosta J, Galarreta L. Observatorio de Commodities:Cacao. Peru:Ministerio de Desarrollo Agrario y Riego;2020.
7. Guiltinan MJ. Cacao. In:Pua EC, Davey MR, editors. Transgenic Crops V. Biotechnology in Agriculture and Forestry. Vol. 60. Berlin, Heidelberg:Springer;2007. 497-518. Available from:https://link.springer.com/10.1007/978-3-540-49161-3_21 [Last accessed on 2023 Dec 01].
8. SodréGA, Sena AR. Cocoa propagation, technologies for production of seedlings. Rev Bras Frutic 2019;41:e782. [CrossRef]
9. Maximova SN, Young A, Pishak S, Guiltinan MJ. Field performance of Theobroma cacaoL. plants propagated via somatic embryogenesis. In Vitro Cell Dev Biol Plant 2008;44:487-93. [CrossRef]
10. Guillou C, Fillodeau A, Brulard E, Breton D, De Faria Maraschin S, Verdier D, et al. Indirect somatic embryogenesis of Theobroma cacao L. in liquid medium and improvement of embryo-to-plantlet conversion rate. In Vitro Cell Dev Biol Plant 2018;54:377-91. [CrossRef]
11. Altamura MM, Della Rovere F, Fattorini L, D'Angeli S, Falasca G. Recent Advances on Genetic and Physiological Bases of in vitro Somatic Embryo Formation. In:Germana MA, Lambardi M, editors. In VitroEmbryogenesis in Higher Plants. Methods in Molecular Biology. Vol. 1359. New York:Springer New York;2016. 47-85. Available from:https://link.springer.com/10.1007/978-1-4939-3061-6_3 [Last accessed on 2023 Dec 01]. [CrossRef]
12. Isah T. Induction of somatic embryogenesis in woody plants. Acta Physiol Plant 2016;38:118. [CrossRef]
13. Suprasanna P, Bapat VA. Differential Gene Expression during Somatic Embryogenesis. In:Mujib A, Šamaj J, editors. Somatic Embryogenesis. Plant Cell Monographs. Vol. 2. Berlin, Heidelberg:Springer-Verlag;2006. 305-20. Available from:https://link.springer.com/10.1007/7089_038 [Last accessed on 2023 Dec 01]. [CrossRef]
14. Garcia C, Marelli JP, Motamayor JC, Villela C. Somatic embryogenesis in Theobroma cacao L. In:Loyola-Vargas VM, Ochoa-Alejo N, editors. Plant Cell Culture Protocols. Methods in Molecular Biology. Vol. 1815. New York:Springer New York;2018. 227-45. Available from:https://link.springer.com/10.1007/978-1-4939-8594-4_15 [Last accessed on 2023 Dec 01]. [CrossRef]
15. Adu-Gyamfi R, Wetten A, Marcelino Rodríguez López C. Effect of cryopreservation and post-cryopreservation somatic embryogenesis on the epigenetic fidelity of cocoa (Theobroma cacao L.). PLoS One 2016;11:e0158857. [CrossRef]
16. Florez SL, Erwin RL, Maximova SN, Guiltinan MJ, Curtis WR. Enhanced somatic embryogenesis in Theobroma cacao using the homologous BABY BOOM transcription factor. BMC Plant Biol 2015;15:121. [CrossRef]
17. Alburqueque-Vasquez NJ, Hernández-Amasifuen AD, Pineda-Lázaro AJ, Fernández Ponce JN, Guerrero-Abad JC, García L, et al. Response of the transcription factor BABY BOOM of Arabidopsis thaliana L. in the formation of embryogenic calluses of cocoa leaves (Theobroma cacao L.). Sci Agropecu 2023;13:359-66. [CrossRef]
18. Fister AS, Landherr L, Maximova SN, Guiltinan MJ. Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in theobroma cacao. Front Plant Sci 2018;9:268. [CrossRef]
19. Chen JT, Chang WC. Induction of repetitive embryogenesis from seed-derived protocorms of Phalaenopsis amabilisvar. Formosa shimadzu. In Vitro Cell Dev Biol Plant 2004;40:290-3. [CrossRef]
20. Deng W, Luo K, Li Z, Yang Y. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci 2009;177:43-8. [CrossRef]
21. Maximova SN, Alemanno L, Young A, Ferriere N, Traore A, Guiltinan MJ. Efficiency, genotypic variability, and cellular origin of primary and secondary somatic embryogenesis of Theobroma cacao L. In Vitro Cell Dev Biol Plant 2002;38:252-9. [CrossRef]
22. Ciridhar P, Indu EP, Ravishankar GA, Chandrasekar A. Influence of triacontanol on somatic embryogenesis in Coffea arabica L. and Coffea canephora P. ex Fr. In Vitro Cell Dev Biol Plant 2004;40:200-3. [CrossRef]
23. Von Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult 2002;69:233-49. [CrossRef]
24. Te-Chato S, Hilae A. High-frequency plant regeneration through secondary somatic embryogenesis in oil palm (Elaeis guineensis Jacq. var. Tenera). J Agric Technol 2007;3:345-57.
25. Muilu-MäkeläR, Vuosku J, Hamberg L, Latva-MäenpääH, Häggman H, Sarjala T. Osmotic stress affects polyamine homeostasis and phenolic content in proembryogenic liquid cell cultures of Scots pine. Plant Cell Tiss Organ Cult 2015;122:709-26. [CrossRef]
26. Chagas K, Cipriano JL, Lopes JC, Schmildt ER, Otoni WC, Alexandre RS. The effects of an osmoregulator, carbohydrates and polyol on maturation and germination of 'Golden THB'papaya somatic embryos. An Acad Bras Ciênc 2018;90:3433-47. [CrossRef]
27. Driver JA, Kuniyuki AH. In vitro propagation of paradox walnut rootstock. HortScience 1984;19:507-9. [CrossRef]
28. Lloyd G, McCown B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot tip culture. Proc Int Plant Prop Soc 1980;30:421-7.
29. Gamborg OL. Aromatic metabolism in plants. II. Enzymes of the shikimate pathway in suspension cultures of plant cells. Can J Biochem 1966;44:791-9. [CrossRef]
30. Stasolla C, Van Zyl L, Egertsdotter U, Craig D, Liu W, Sederoff RR. The effects of polyethylene glycol on gene expression of developing white spruce somatic embryos. Plant Physiol 2003;131:49-60. [CrossRef]
31. Lincy AK, Remashree AB, Sasikumar B. Indirect and direct somatic embryogenesis from aerial stem explants of ginger (Zingiber officinale Rosc.). Acta Bot Croat 2009;68:93-103.
32. Viji M, Maheswari P, Karuppanapandian T, Manoharan K. Effect of polyethylene glycol and mannitol on somatic embryogenesis of pigeonpea, Cajanus cajan(L.) Millsp. Afr J Biotechnol 2012;11:10340-9. [CrossRef]
33. De Oliveira LF, Navarro BV, Cerruti GV, Elbl P, Minocha R, Minocha SC, et al. Polyamine- and amino acid-related metabolism:The roles of arginine and ornithine are associated with the embryogenic potential. Plant Cell Physiol 2018;59:1084-98. [CrossRef]
34. Al-Khayri JM, Al-Bahrany AM. Effect of abscisic acid and polyethylene glycol on the synchronization of somatic embryo development in date palm (Phoenix dactylifera L.). Biotechnology 2012;11:318-25. [CrossRef]
35. Martin D. Somatic embryogenesis:a biotechnological tool for in vitro propagation of guajava. Biotecnol Vegetal 2017;17:209-20.
36. Urrea AI, Atehortúa L, Gallego AM. Regeneración vía embriogénesis somática de una variedad colombiana élite de Theobroma cacao L. Rev Colomb Biotecnol 2011;13:39-50.
37. Al-Khayri JM, Al-Bahrany AM. Growth, water content, and proline accumulation in drought-stressed callus of date palm. Biol Plant 2004;48:105-8. [CrossRef]
38. Nadel BL, Altman A, Ziv M. Regulation of somatic embryogenesis in celery cell suspensions:2. Early detection of embryogenic potential and the induction of synchronized cell cultures. Plant Cell Tiss Organ Cult 1990;20:119-24. [CrossRef]
39. Díaz LC, Carmona OE, Beltrán JD. Optimization of in vitro conservation of Dioscorea spp germplasm by minimal growth. Rev Colomb Biotecnol 2015;17:32-9. [CrossRef]
40. Pinto G, Santos C, Neves L, Araújo C. Somatic embryogenesis and plant regeneration in Eucalyptus globulus Labill. Plant Cell Rep 2002;21:208-13. [CrossRef]
41. Cárdenas-Lara MA, Villegas-Monter Á. Osmotic potential of culture medium with different compounds for the in vitro propagation. Rev Fitotec Mex 2002;25:213-7.
42. Márquez-Martín B, Sánchez-Romero C, Perán-Quesada R, Barceló-Muñoz A, Pliego-Alfaro F. Factores Que Afectan a la Obtención de Embriones Somáticos Blanco-opacos de Aguacate. In:Proceedings V World Avocado Congress;2003. p. 97-102.
43. Barbón R, Borroto I, Mariana La O, Quiala E, Pérez M. Effect of sorbitol and 6-BAP in Swietenia mahagoni (L.) Jacq. somatic embryos maturation and germination Jacq. Biotecnol Vegetal 2013;13:51-5.