
1. INTRODUCTION
Orchids are a unique group of plants whose beautiful and fragrant flowers have fascinated humans, and their medicinal properties have cured and saved human lives from several acute and chronic sufferings [1]. Aerides odoratum L. is a medicinal and ornamental orchid of the Eastern Himalayas found at altitudes of 800 m–1200 m. The leaf juice has antimicrobial properties and is used to cure boils in the ears and nose by tribal people [2,3]. Unfortunately, use of this orchid in pharmacology is nought because of its rare occurrence, slow growth, longer life cycle, and lack of artificial methods for large-scale propagation of superior variants [4].
Chemical-induced mutagenesis is one of the major breeding approaches for plant improvement. Ethyl Methane Sulfonate (EMS) has been recommended to improve cultivars of vegetatively propagated plants, especially ornamental plants. In vitro mutation induction in plants provides an efficient method for its rapid mass propagation and selection of better stress-tolerant morphological variants with improved phytochemical and osmolite contents [5]. The application of mutation techniques for improving both seed and vegetatively propagated plants has been made easier by the advent of effective in vitro culture methods [6]. Mutation induction, combined with in vitro culture techniques, has become the only viable option for plant regeneration and improvement in many vegetatively propagated crops, notably slow-growing ornamental plants. Effective use of low-dose mutagens on a large number of propagules and the ease of handling plantlets are a few benefits of in vitro-induced mutation [7].
Using a potent chemical mutagen, like EMS, is one method of causing mutation. It is a highly recommended chemical mutagen due to its low cost and easy application methods; however, the temperature of the solution, treatment period, and concentration are the three most crucial factors when using EMS to induce mutation [8]. EMS typically results in point mutations in plants, but it can also cause chromosome loss or deletion. Consequently, EMS may be able to modify loci of specific interest without causing a significant number of closely related alterations. However, it can cause cell death either instantly or by mitotic death that occurs when the cell divides [9]. Research has shown that EMS can be used to produce mutants in a variety of crop plants [9-12] However, very few studies are available claiming the success of EMS-induced mutations in orchids [13,14]. Therefore, this study aimed to establish superior A. odoratum mutant lines in terms of morphology and osmolytes, pigments, and phenol contents.
2. MATERIALS AND METHODS
2.1. Seed Source and Establishment of in vitro Culture
The study design is illustrated in Figure 1. Mature pods of A. odoratum were collected from their natural habitat. They were surface sterilized by liquid detergent for 5–10 min and then washed thoroughly in distilled water and subsequently double-distilled water to remove all the traces of detergent from the pod. Further, pods were disinfected using mercuric chloride solution (0.5% w/v) and 70% ethanol. The sterilized pods were split and opened in a laminar air flow unit (LAF) with a sterile surgical blade [15]. Powdery seeds were kept on nutrient media MSBM (Murashige and Skoog, 1962) fortified with a combination of coconut water (10%), kinetin (4.65 μM), and casein hydrolysate (250 mg/l) to obtain optimum seed germination and protocorm formation. The cultures were incubated at standard in vitro culture conditions.
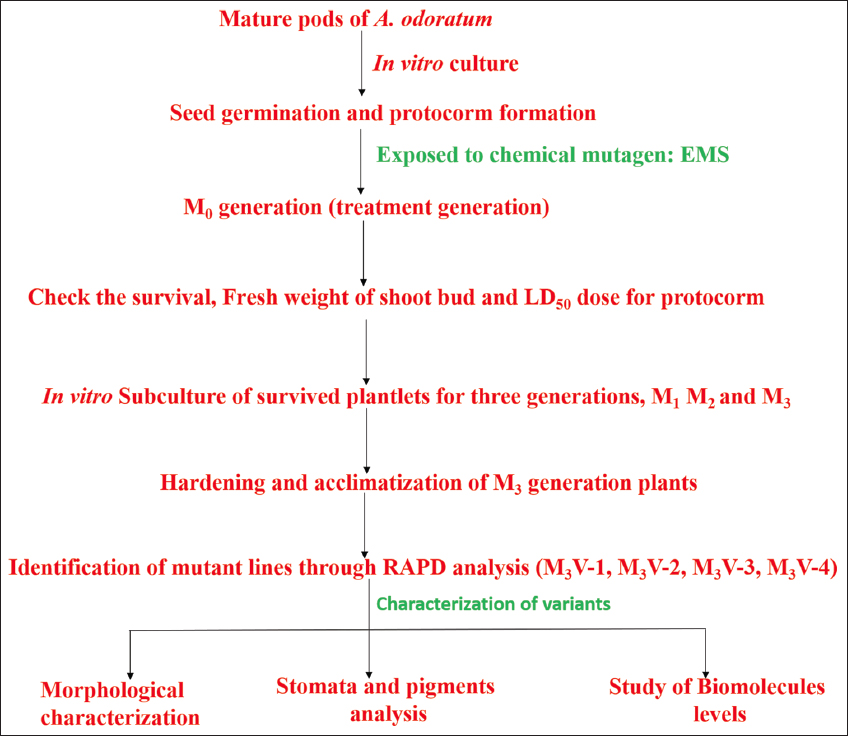 | Figure 1: A schematic diagram of experimental design for in vitro mutation breeding in A. odoratum. [Click here to view] |
2.2. In vitro Induction of Mutation
Doses and duration of treatment were decided based on a trial-and-error method. Ethyl methylsulfonate (SIGMA-Aldrich) stock solution (200 mg/mL) was prepared by adding 1 mL of sterile water to 200 mg of EMS and sterilized with a millipore filter (pore size: 0.01 μm) under LAF. In the sterilized culture media, the required volume of EMS was added to get different concentrations of mutagenic agents ranging from 0.025% to 5%. Further media with EMS were dispensed into the tubes or bottles containing nutrient media. Young protocorms (30 days old) were transferred to 120 bottles having different concentrations of EMS in mutagenic media [Figure 2]. 10 bottles of each EMS concentration to be studied were kept for different time intervals (2, 5, 7, and 10 days) to induce mutation. After the treatment of chemical mutagens, the protocorms were subcultured in the differentiation and multiplication media, MSBM in combination with Kn (4.65 μM) and TDZ (4.54 μM), and produced multiple shoots. The effect of mutagen on protocorms was recorded in terms of the percent survival of protocorms the increase or decrease in the number and weight of shoot bud initials. Further, half-strength MSBM supplemented with NAA (5.46 μM) was used to develop root initials. 10–20 plantlets of each treatment showing variation in the number and weight of shoots in M0 generation (treatment generation) were maintained for three subsequent generations: M1 (first generation after treatment), M2 (second generation after treatment), and M3 (third generation after treatment) under in vitro conditions. Leaf segments of M0, M1, and M2 generation seedlings were excised and subcultured on regeneration media in subsequent generations to obtain multiple shoots and roots. M3 generation mutants having strong and stout roots and 3–6 leaflets were hardened and acclimatized in the Orchidarium [7,10].
 | Figure 2: Protocorms subjected to mutagen treatment. [Click here to view] |
2.3. Evaluation of Mutants at M0 Generation
M0 generation-treated protocorms were evaluated for mutagenic effects based on variation in survival frequency and fresh weight (FW).
2.3.1. Percent survival of protocorms
Mutagen-treated protocorms were observed under a stereo microscope to find the effect of different doses of mutagen in terms of shrinkage, death, and/or shoot initiation, and percent survival was calculated.
2.3.2. FW
The treated protocorms were taken out of the culture bottles when they attained maximum growth, and the moisture was gently removed by blotting with filter paper and transferred to pre-weighed aluminum foil. The weight of the protocorm was determined with the help of an electronic balance.
2.4. Identification of Mutants through RAPD Analysis in M3 Generation
M3-generation 18-month-old plants were screened for molecular characterization. A total of 50 M3-generation plants, who were apparently looking morphologically different than the control, were used for molecular analysis. DNA was isolated from a hundred-day-old hardened mutant plant leaf using the standard CTAB method. Universal RAPD genetic marker: OPA-18 forward and reverse primers were used to amplify the DNA of control and mutant plants. Mutants were identified by looking for unique bands on the DNA gel picture.
2.5. Study of Morphological Parameters at the M3 Generation
Variations in morphological parameters such as plant height and weight, number of leaves, leaf length, breadth, and root length were studied among control and mutated plants in the M3 generation.
2.5.1. Mutation spectrum of chlorophyll at M3 generation
Mutant plantlets from the M3 generation were screened for changes in the chlorophyll spectrum caused by mutation. Chlorophyll mutants were categorized as follows [16]:
1.Albino: Leaves are white, devoid of chlorophyll and carotenoids, and typically have few plastids
2.Xantha: Yellow leaves with carotenoid pigments but little to no chlorophyll
3.Alboviridis: The rate at which plastids develop at the base and tips of leaves varies for each leaf and displays various hues in these sections.
4.Viridis: Their color is a pale green.
5.Tigrina: Display alternating narrow bands of green, yellow, or brown transverse stripes along the leaf in areas where pigment degradation has taken place.
6.Striata: These contain longitudinal stripes that alternate between green and yellow or white.
7.Maculata: Contains areas where carotene and/or chlorophyll have been damaged.
2.6. Stomatal Length (SL) and Density at M3 Generation
SL, μm was measured with the help of a scale present in the software from 30 stomata selected randomly. Stomatal density is a simple calculation of the number of stomata seen in a millimeter square of tissue. Grids of 100 μm2 were composed on the microscopic image to count the number of stomata and epidermal cells (E) within that. Stomatal density was calculated using the following formula: Stomatal Density = Number of Stomata present in 1 grid (S) X 100.
2.7. Estimation of Chlorophyll and Carotenoids at M3 Generation
Chlorophyll and carotenoids were estimated using the modified Holm method [17]. The concentration of chlorophyll was calculated by Porra’s (2002) equation [18]. Carotenoid concentration was calculated using Lichtenthaler and Wellburn’s equation [19].
2.8. Extraction of Sugars and Amino Acids at M3 Generation
Leaf samples that had been oven-dried at 60° C were blended with 80% hot ethanol, centrifuged for 10 min at 2000 rpm, and the supernatant was poured off. The residue mixture was then treated with 3 mL of 80% ethanol, and recentrifugation was performed. To guarantee that all sugars and amino acids were recovered, the extraction process was done twice. The residue was saved to estimate starch. A boiling-water bath at 45°C was used to cool and evaporate the supernatant, leaving a Petri dish-like residue that was eluted with 5 mL of 20% ethanol before being subjected to analysis for sugars and amino acids.
2.8.1. Quantification of total sugars at M3 generation
Quantification of total sugars was made by following the “anthrone method.” Using a standard curve that was plotted using a known quantity of glucose, the concentration of total sugars was determined [20].
2.8.2. Reducing sugar estimation at M3 generation
The DNSA method was used to estimate the amount of reduced sugars. Based on a standard curve that was produced with a known quantity of glucose, the amount of reducing sugars was determined [21].
2.8.3. Estimation of amino acids at M3 generation
Amino acid content was estimated by the “Ninhydrin method”. The concentration of free amino acids was calculated from a standard curve with a known concentration of glycine [22].
2.8.4. Estimation of total phenols at M3 generation
Concentrations of total phenol were estimated in M3 generation plant tissues using the modified method proposed by Waterman and Mole (1994) and expressed in terms of gallic acid equivalent (mg g–1 of dry mass) [23].
2.9. Statistical Analysis
The percent of data was calculated in MS Excel (2019) software. The data obtained were subjected to a one-way ANOVA in SPSS software, Version 26. The mean values, standard deviation, and level of significance were calculated.
3. RESULTS AND DISCUSSION
3.1. Effect of EMS Treatment on M0 Generation
The effect of EMS on survival frequency was scored based on shoot bud initiation after 60–80 days of treatment. It was found that the survival percentage of protocorms decreased with increasing concentration and time duration of EMS exposure. Protocorms [Figure 2] exposed to different concentrations of EMS for 2 days showed more than 50% survival frequency, ranging from 89 ± 7% to 55 ± 8%, and the maximum FW of shoot buds ranged between 40 ± 7 mg and 20 ± 5 mg. There was no significant difference in survival frequency or FW between treated and control plantlets. Hence, this duration was found ineffective in changing the survival frequency and weight of the shoot bud because shorter exposure is usually unproductive for the uptake and penetration of EMS into protocorms and seeds under in vitro conditions [24,25].
Treatment of protocorms with different doses of EMS for 5 days was found to be most effective in causing mutagenic effects. The frequency of protocorm survival ranged between 81 ± 2% and 2 ± 2% at doses of 0.025% and 0.5%, respectively. EMS concentrations of 0.3% showed an approximate 50% survival frequency at 5 and 7 days of treatment, whereas a drastic reduction in survival was noted with further increases in EMS concentrations. EMS (0.5%) was found to be lethal and showed only 2% survival frequency for 5-day treatment and zero survival for longer-term treatment. Radical variations in the FW of shoot buds on EMS treatments for 5 and 7 days of exposure were noted, which may be due to the mutagenic effect. The effect of EMS on A. odoratum is evident by the increasing concentration and duration of treatment. Similar doses were noted as effective in other studies on orchids as well [7,14]. Maximum FW of 60 ± 7 mg was attained at EMS (0.1%) and minimum 4 ± 3 mg at EMS (0.5%) for 5 days of treatment, whereas it varied from 37 ± 7 mg to 0 for 7 days of treatment [Table 1]. Variation in FW as an effect of different doses of EMS was also observed in orchids [7] and other species [26].
Table 1: Effect of different doses and time duration of EMS treatment on the survival frequency and fresh weight of shoot buds.
| EMS doses (%) | Treatment for 2 days | Treatment for 5 days | Treatment for 7 days | Treatment for 10 days | ||||
|---|---|---|---|---|---|---|---|---|
| Survival frequency (%) | Fresh weight of shoot bud | Survival frequency (%) | Fresh weight of shoot bud | Survival frequency (%) | Fresh weight of shoot bud | Survival frequency (%) | Fresh weight of shoot bud | |
| Control | 85±5 | 32±3 | 85±5 | 32±3 | 85±5 | 32±3 | 85±5 | 32±3 |
| 0.025 | 89±7 | 33±5 | 81±2 | 43±5 | 85±9 | 37±7 | 50±3 | 31±7 |
| 0.05 | 82±8 | 37±7 | 78±9 | 52±6 | 81±7 | 48±8 | 74±9 | 38±9 |
| 0.75 | 79±11 | 34±4 | 72±11 | 56±6 | 79±7 | 41±7 | 88±6 | 36±5 |
| 0.1 | 75±9 | 36±9 | 75±10 | 60±7 | 72±11 | 43±2 | 67±3 | 30±4 |
| 0.125 | 82±8 | 38±5 | 78±9 | 46±9 | 66±8 | 40±8 | 57±3 | 26±7 |
| 0.15 | 85±9 | 32±6 | 67±14 | 50±11 | 62±6 | 33±2 | 51±7 | 21±9 |
| 0175 | 80±12 | 39±9 | 68±9 | 38±6 | 60±11 | 31±6 | 48±5 | 21±3 |
| 0.2 | 88±14 | 40±7 | 69±8 | 34±4 | 56±13 | 28±5 | 40±3 | 17±5 |
| 0.225 | 92±8 | 36±8 | 61±9 | 33±5 | 52±6 | 31±7 | 41±4 | 14±3 |
| 0.25 | 85±9 | 30±4 | 58±9 | 42±6 | 36±3 | 38±9 | 29±6 | 12±8 |
| 0.275 | 87±12 | 25±4 | 55±11 | 36±8 | 46±4 | 36±5 | 6±2 | 11±6 |
| 0.3 | 88±11 | 36±4 | 50±12 | 30±9 | 48±4 | 30±4 | 18±3 | 17±3 |
| 0.325 | 82±8 | 39±4 | 40±12 | 33±5 | 33±4 | 29±5 | 14±9 | 16±5 |
| 0.35 | 85±12 | 36±8 | 38±9 | 32±6 | 32±3 | 26±5 | 11±3 | 13±8 |
| 0.375 | 88±3 | 33±8 | 32±11 | 26±6 | 27±1 | 19±5 | 14±3 | 16±4 |
| 0.4 | 81±11 | 34±16 | 30±6 | 20±5 | 21±9 | 31±7 | 10±4 | 15±4 |
| 0.425 | 70±7 | 36±5 | 25±8 | 13±8 | 13±2 | 26±5 | 5±4 | 7±3 |
| 0.45 | 64±9 | 24±6 | 10±4 | 19±6 | 11±4 | 23±8 | 0 | 0 |
| 0.475 | 55±8 | 20±5 | 8±3 | 10±9 | 1±1 | 11±3 | 0 | 0 |
| 0.5 | 60±7 | 23±8 | 2±2 | 4±3 | 0 | 0 | 0 | 0 |
* All the data is mean±SD.
EMS doses of 0.3% and 0.5% were noted as LD50 and lethal doses, respectively [Table 1]. EMS treatment for 10 days showed deleterious effects of mutagenesis, resulting in a decrease in FW of shoot buds and <50% protocorm survival except at doses of 0.025% to 0.15% [Figures 3-5]. A similar genotoxic effect of EMS was noted in a wide range of plant species, including Eclipta alba [27], rose [28], citrus [29], and mungbean [30]. Further, the time required for shoot bud formation was exponentially increased with increasing the concentration of EMS in the present study. The delay in seed germination as well as the reduction in seed germination percentage with increasing ethyl methanesulfonate doses were also reported in crop plants such as lentil and wheat [31,32].
 | Figure 3: Survival of protocorms at LD50 Dose, EMS (0.3%). [Click here to view] |
 | Figure 4: Shoot bud formation at Lethal Dose, EMS (0.5%). [Click here to view] |
 | Figure 5: Development of axenic plantlet at M0 generation. [Click here to view] |
Surviving plantlets were maintained for three generations. M3-generation plantlets [Figure 6] were maintained at culture conditions for 100 days. Finally, 501 axenic plantlets [Figure 7] were obtained by treatment with different EMS doses for various time durations in M3 generation. These axenic plantlets were hardened and acclimatized to a potting mixture containing broken brick pieces, peat, and small pieces of charcoal (1:1:1 w/v) and maintained in Orchidarium for 18 months with a 75% survival frequency [Figure 8].
 | Figure 6: Development of axenic plantlet at M3 generation. [Click here to view] |
 | Figure 7: M3 generation plantlet ready for hardening. [Click here to view] |
 | Figure 8: M3 generation, 300 days old hardened plants. [Click here to view] |
3.2. Identification of Mutant Lines Using RAPD Analysis
RAPD analysis is one of the simplest and most accurate methods to identify epigenetic and somaclonal variations among variants. It has been used for the identification of mutations in Allium sativum L [33] and genetic stability in Rhynchostylis retusa [34]. Four mutant lines were established among variants of A. odoratum, which showed unique RAPD bands [Figure 9]. These mutant lines were denoted as M3V-1-M3V-4. Mutant line M3V-1 was obtained from 0.05% EMS treatment for 5 days, whereas M3V-2 was produced from 0.075% EMS treatment for 5 days. M3V-3 was formed from 0.1% EMS treatment for 5 days, and the M3V-4 mutant was obtained using EMS (0.05%) for 7 days. Similar results were noted in a study by Srivastava et al. [7].
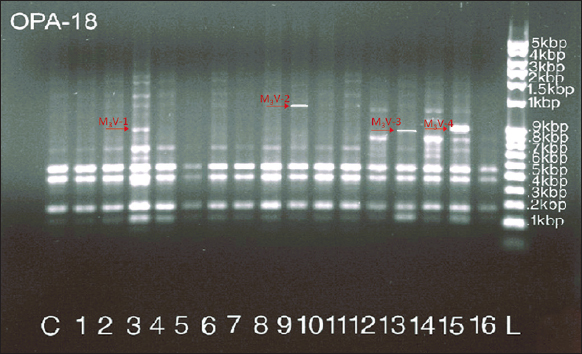 | Figure 9: RAPD analysis of hardened mutant plants. [Click here to view] |
3.3. Evaluation of Mutants in M3 Generation
Among four mutant lines (M3V-1-M3V-4) isolated from EMS treatment in M3 generation, superior morphological mutants M3V-1, M3V-2, and M3V-3 evolved from EMS 0.05%, 0.075%, and 0.1% treatments for 5 days, respectively, showed superior morphological, stomatal osmolytes, pigments, and phenol contents than control and were significantly diverse in morphology among them also [Table 2]. Among the three mutant lines, M3V-1 showed the least plant height (3.49 ± 0.03 cm), weight (549.83 ± 6.76 mg), root length (1.93 ± 0.09 cm), and number of leaves (3.39 ± 0.09) but the longest (3.99 ± 0.04 cm) and thinnest (1.27 ± 0.03 cm) leaf compared to the control [Figures 10 and 11]. It can also be distinguished by having maximum chlorophyll contents (4.42 ± 0.11 μg/mg) and total phenol (42.6 ± 0.6 mgGAE/g DW) [Table 2]. The variation among three morphologically superior mutants (M3V-1, M3V-2, and M3V-3) can be explained as mutations of genes, the breaking of tightly linked regions and crossing over within these regions, enhanced recombination, or an individual or a combination of two or more such effects [35,36].
 | Figure 10: Leaf of control A. odoratum. [Click here to view] |
 | Figure 11: Longer and thinner mutant leaf. [Click here to view] |
Table 2: Effect of EMS on the morphology of M3 generation plants in Aerides odoratum.
| EMS doses (%) | Plant height (cm) | Plant weight (mg) | Number of leaves | Root length (cm) | Leaf length (cm) | Leaf breadth (cm) | Length of stomata (mm) | Stomatal density |
|---|---|---|---|---|---|---|---|---|
| Control | 3.20±0.04 | 520.60±9.24 | 2.94±0.06 | 1.47±0.07 | 2.89±0.07 | 1.36±0.05 | 24.8±0.9 | 99.4±7.9 |
| 0.05% for 5 days (M3V-1) | 3.49±0.03 | 549.83±6.76 | 3.39±0.09 | 1.93±0.09 | 3.99±0.04 | 1.27±0.03 | 18.8±1.9 | 118.9±1.5 |
| 0.075% for 5 days (M3V-2) | 4.48±0.08 | 648.47±3.97 | 3.99±0.11 | 2.45±0.08 | 3.47±0.12 | 1.50±0.07 | 21.4±1.6 | 128.5±3.7 |
| 0.1% for 5 days (M3V-3) | 3.96±0.02 | 595.03±9.50 | 3.67±0.13 | 2.95±0.13 | 3.69±0.07 | 1.67±0.13 | 20.0±1.4 | 136.0±5.9 |
| 0.05% for 7 days (M3V-4) | 3.27±0.04 | 511.52±2.52 | 2.19±0.09 | 1.20±0.08 | 2.81±0.04 | 1.28±0.04 | 23.8±1.6 | 122.5±6.5 |
*All the data is mean±SD, P<0.05.
Mutant line M3V-2 showed maximum plant height 4.48 ± 0.08 cm, weight (648.47 ± 3.97 mg), number of leaves (3.99 ± 0.11), total soluble sugar (265.6 ± 5.6 μg/gDW), reducing sugar (228.5 ± 3.7 μg/gDW), and amino acids (161.2 ± 3.2 μg/gDW) [Table 3]. M3V-3 showed a maximum root length of 2.95 ± 0.13 cm and the broadest leaf (1.67 ± 0.13 cm) as significantly distinguishable morphological characters. Mutation by EMS is known to increase resistance to abiotic stress through the accumulation of osmolytes such as carbohydrates and amino acids. These compounds serve to maintain turgor pressure in the form of organic and inorganic compounds such as amino acids, sucrose, soluble sugars, reducing sugars, and various other solutes in the cytoplasm to increase the absorption of water from the soil [37,38]. In the present study, all morphological mutants showed an enhanced level of these osmolytes, which may be considered stress-resistant mutant lines for further studies.
Table 3: Effect of EMS on osmolytes, pigments, and total phenol concentration of M3 generation leaf in Aerides odoratum.
| EMS doses (%) | Total soluble sugar (mg/gDW) | Reducing sugar (mg/gDW) | Amino acids (mg/gDW) | Total Chlorophyll (mg/mg) | Carotenoid (mg/mg) | Total phenol (mgGAE/g DW) |
|---|---|---|---|---|---|---|
| Control | 174.5±4.4 | 123.2±6.7 | 88.4±5.5 | 3.22±0.05 | 1.50±0.09 | 20.5±0.8 |
| M3V-1 | 199.7±3.8 | 145.1±5.1 | 139.1±8.4 | 4.42±0.11 | 2.32±0.15 | 42.6±0.6 |
| M3V-2 | 265.6±5.6 | 228.5±3.7 | 161.2±3.2 | 3.91±0.14 | 2.62±0.10 | 35.5±0.3 |
| M3V-3 | 223.4±6.2 | 180.7±4.8 | 107.4±4.7 | 4.19±0.17 | 1.91±0.09 | 29.5±1.4 |
| M3V-4 | 188.0±3.4 | 131.3±2.3 | 89.1±6.4 | 3.02±0.18 | 1.19±0.07 | 21.8±3.2 |
*All the data is mean±SD, P<0.05.
The length of stomata in M3V-1, M3V-2, and M3V-3 was shorter, but the stomatal density was significantly greater than the control. For M3V-4, although the length of the stomata was shorter than that of the control, the difference was not significant. However, the stomatal density was significantly higher than the control in M3V-4. Further, an increase in chlorophyll, carotenoid, and total phenolic contents was noted as an important impact of mutation in all species. Similar effects were noticed in many stress-resistant crops, including orchids [7,17,39,40].
The M3V-4 mutant line showed the most distinguishing feature, the viridis chlorophyll spectrum in leaves. The leaf was thickest, having spots where chlorophyll and/or carotene had been destroyed. Less quantity of chlorophyll (3.02 ± 0.18 μg/mg) and carotenoid (1.19 ± 0.07 μg/mg) compared to the control estimation confirmed the morphological mutation in the M3V-4 mutant line [Table 3]. The chlorophyll variegation in leaves after EMS treatment is common to change and seen in different ornamentals, which may be due to the high specificity of EMS for mitochondrial and plastid genome mutations [7,41,42]. Our investigation of improved stomatal density, chlorophyll, and carotenoid contents in the M3V-4 mutant line also supports the formation of viridis variegation in the leaf [Figure 12].
 | Figure 12: Thinner mutant leaf with viridis chlorophyll spectrum. [Click here to view] |
4. CONCLUSION
The use of low EMS doses on protocorms of orchids can generate superior variants of orchids in terms of morphological and biomolecular characteristics, which may be considered stress-resistant mutant lines for further studies. Four mutant lines (M3V1-M3V4) were established based on RAPD analysis of the leaf DNA of 300-day-old hardened plants. M3V-1 (developed from 0.05% EMS for 5 days of treatment) was noted as the best among all mutant lines based on morphological analysis, whereas M3V-2 (generated from 0.075% EMS for 5 days treatment) was noted as the most superior in terms of having the highest quantity of osmolytes, pigments, stomata, and phenol. M3V-4 (0.05% EMS for 7 days) showed the viridis chlorophyll spectrum as the most distinguishing factor. EMS (0.1%) for 5 days was responsible for the creation of M3V-3, which had maximum root length, leaf breadth, and carotenoids. The present study developed the first method for the in vitro mutation of A. odoratum, which provides a framework for implementing mutation breeding and improving the orchid species. Future studies on identifying and encoding genes responsible for these mutations can help in directly introducing agronomically important features into orchid plants.
5. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be authors as per the International Committee of Medical Journal Editors (ICMJE) requirements and guidelines.
6. FUNDING
There is no funding to report.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. DATA AVAILABILITY
The authors confirm that the data supporting the findings of this study are available within the article.
10. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
11. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Teoh ES, Teoh ES. Aerides Lour. Orchid Species from Himalaya and Southeast Asia Vol. 1 (A-E). Berlin:Springer;2021. 19-30. [CrossRef]
2. Jyotshna BS, Kavitha KR, Srivastava D. Orchids:Indispensable members of horticulture in India. Pract Res Hortic 2023;10:59-73.
3. Jhansi K, Rahamtulla M, Kishore I, Khasim S. Phytochemical and therapeutic potential of Aerides odorata lour. (Orchidaceae):An overview. Phytochem Compos Pharm Med Plants 2023;2:201.
4. Paraste VK, Sarsaiya S, Mishra UC, Sourabh P. A comprehensive review on global research trends on Aerides genus with reference to Aerides odorata species. J Appl Biol Biotechnol 2023;11:55-62. [CrossRef]
5. Sarsu F, Penna S, Nikalje GC. Strategies for screening induced mutants for stress tolerance. In:Mutation Breeding for Sustainable Food Production and Climate Resilience Singapore. Berlin:Springer Nature Singapore;2023. 151-76. [CrossRef]
6. Hwang D, Jeong HJ, An JT, Kwon JK, Kang SY, Kang BK. Development of an Ethyl Methane Sulfonate (EMS) Mutant Lines in C. annuum. In:Breakthroughs in the Genetics and Breeding of Capsicum and Eggplant. Proceedings of the XV EUCARPIA Meeting;2013. 505.
7. Srivastava D, Gayatri MC, Sarangi SK. In vitro mutagenesis and characterization of mutants through morphological and genetic analysis in orchid Aerides crispa Lindl. Indian J Exp Biol 2018;56:385-94.
8. Talebi AB, Talebi AB, Shahrokhifar B. Ethyl methane sulphonate (EMS) induced mutagenesis in Malaysian rice (cv. MR219) for lethal dose determination. Am J Plant Sci 2012;3:1661-5. [CrossRef]
9. Siddique MI, Back S, Lee JH, Jo J, Jang S, Han K, et al. Development and characterization of an ethyl methane sulfonate (EMS) induced mutant population in Capsicum annuum L. Plants (Basel) 2020;9:396. [CrossRef]
10. Arisha MH, Shah SN, Gong ZH, Jing H, Li C, Zhang HX. Ethyl methane sulfonate induced mutations in M2 generation and physiological variations in M1 generation of peppers (Capsicum annuum L.). Front Plant Sci 2015;6:399. [CrossRef]
11. Nasri F, Zakizadeh H, Vafaee Y, Mozafari AA. In vitro mutagenesis of Chrysanthemum morifolium cultivars using ethyl methanesulphonate (EMS) and mutation assessment by ISSR and IRAP markers. Plant Cell Tissue Organ Cult 2021;149:1-7. [CrossRef]
12. Lapjit C, Pan IC, Chen YM, Tseng MJ. Induction of mutagenesis in Erycina pusilla by ethyl methanesulfonate and sodium azide. Horticulture NCHU 2015;40:81-93.
13. Kurniadi AS, Irawati F, Putra SE, Hardjo PH. Induction of protocorm-like bodies (PLBs) Phalaenopsisspp. hybrids mutation through ultraviolet irradiation (UV254) and ethyl methane sulfonate (EMS). Agriprima J Appl Agric Sci 2023;7:1-5. [CrossRef]
14. Khairum A, Hualsawat S, Chueakhunthod W, Tharapreuksapong A, Tomsantia B, Yenchon S, et al. Selection and characterization of in vitro-induced mutants of Dendrobium “Earsakul”resistant to black rot. In Vitro Cell Dev Biol Plant 2022;58:577-92. [CrossRef]
15. Mahendran G, Bai VN. Mass propagation of Satyrium nepalense D. Don.-a medicinal orchid via seed culture. Sci Hortic 2009;119:203-7. [CrossRef]
16. Girija M, Dhanavel D. Induced chlorophyll mutation in cowpea (Vigna unguiculata L. Walp.). Int J Curr Res 2013;2:136-40.
17. Kurniawan FY, Amelia DE. Chlorophyll and carotenoid level comparisons of pigeon orchid (Dendrobium crumenatum) in water and light stress treatment. Indones J Sci Educ 2021;5:44-8. [CrossRef]
18. Porra RJ. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth Res 2002;73:149-56. [CrossRef]
19. Deepti S, Gayatri MC, Sitikantha S. Morphological and biochemical changes in pollinated flowers of different Aerides species. Curr Bot 2013;4:33-7.
20. Soni DK, Shahi SK, Khandel P, Mahobiya D, Singh R, Yadaw RK, et al. Extraction and estimation of chlorophylls from epiphytic orchids and their antioxidants scavenging activity analysis. Plant Arch 2018;18:2448-52.
21. Jain A, Jain R, Jain S, Jain A, Jain R, Jain S. Quantitative analysis of reducing sugars by 3, 5-dinitrosalicylic acid (DNSA method). In:Basic Techniques in Biochemistry, Microbiology and Molecular Biology:Principles and Techniques. Berlin:Springer;2020. 181-3. [CrossRef]
22. López-Hidalgo C, Meijón M, Lamelas L, Valledor L. The Rainbow Protocol:A Sequential Method for Quantifying Pigments, Sugars, Free Amino Acids, Phenolics, Flavonoids and MDA from a Small Amount of Sample. Chichester, UK:John Wiley &Sons, Ltd.;2021. 34-45.
23. Waterman PG, Mole S. Analysis of Phenolic Plant Metabolites. Methods in Ecology. Oxford:Blackwell Scientific Publications;1994.
24. Sharma M, Tarafdar A, Pandey A, Ahmed S, Pandey V, Chobe DR, et al. Biotic stresses in food legumes:An update and future prospects. In:Genetic Enhancement in Major Food Legumes:Advances in Major Food Legumes. Berlin:Springer;2021. 149-96. [CrossRef]
25. Singh AK. Induced genetic variability in M3 generation of mungbean. J Food Legum 2009;22:162-5.
26. Yasmin S, Nehvi FA. Ethyl methane sulphonate-A potent mutagen for inducing genetic variability in saffron (Crocus sativus L.). Appl Biol Res 2017;19:164-71. [CrossRef]
27. Kumar G, Mishra R. Micronuclei assay as biomarker for ethyl methane sulphonate (EMS) genotoxicity in Eclipta alba (L.) Hassk. Indian J Genet Plant Breed 2020;80:318-23. [CrossRef]
28. Mehrabi MM, Taghizadeh M, Solgi M. Effect of EMS ethyl methane sulfonate (EMS) mutagen on Iranian rose (Rosa persica Michx) to generate morphological variation. Plant Prod 2022;45:335-46.
29. Bora L, Vijayakumar RM, Ganga M, Ganesan NM, Sarkar M, Kundu M. Determination of mutagenic sensitivity (LD50) of acid lime [Citrus aurantifolia (Christm.) Swingle] cv. PKM-1 to physical and chemical mutagens. Natl Acad Sci Lett 2024;47:73-7-7 [CrossRef]
30. Usharani KS, Kumar CA. Mutagenic efficiency and effectiveness of gamma rays and EMS and their combination in inducing chlorophyll mutations in M2 generation of Urdbean (Vigna mungo (L.) Hepper). Electron J Plant Breed 2015;6:210-7.
31. Shahwar D, Khan Z, Ansari MY. Evaluation of mutagenized lentil populations by caffeine and EMS for exploration of agronomic traits and mutant phenotyping. Ecol Genet Genom 2020;14:100049. [CrossRef]
32. Bahar B, Akkaya MS. Effects of EMS treatment on the seed germination in wheat. J Appl Biol Sci 2009;3:59-64.
33. Mahmoud AM, El-Maaty-Shereen A, Osman-Neama H, Al-Azab KF. Determination of suitable gamma-ray dose for mutation induction in garlic (Allium sativum L.) and identification of genetic diversity by RAPD and ISSR markers. Arab J Biotechnol 2019;22:25-42.
34. Oliya BK, Chand K, Thakuri LS, Baniya MK, Sah AK, Pant B. Assessment of genetic stability of micropropagated plants of Rhynchostylis retusa (L.) using RAPD markers. Sci Hortic 2021;281:110-8. [CrossRef]
35. Hassan MJ, Karim MM, Islam MA, Pramanik MH, Hossain MA. Changes in root porosity and water soluble carbohydrates in rice (Oryza sativa L.) under submergence stress. J Bangladesh Agric Univ 2019;17:539-44. [CrossRef]
36. Kaur S. Effect of mutagens on regeneration and growth of in vitro grown epicotyl segments of rough lemon seedlings (Citrus jambhiri Lush.). J Appl Nat Sci 2015;7:459-65. [CrossRef]
37. Widodo, Patterson JH, Newbigin ED, Tester M, Bacic A, Roessner U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J Exp Bot 2009;60:4089-103. [CrossRef]
38. Chen L, Duan L, Sun M, Yang Z, Li H, Hu K, et al. Current trends and insights on EMS mutagenesis application to studies on plant abiotic stress tolerance and development. Front Plant Sci 2023;13:1052569. [CrossRef]
39. Tang CJ, Luo MZ, Zhang S, Jia GQ, Sha TA, Jia YC, et al. Variations in chlorophyll content, stomatal conductance, and photosynthesis in Setaria EMS mutants. J Integr Agric 2023;22:1618-30. [CrossRef]
40. Rime J, Dinesh MR, Sankaran M, Shivashankara KS, Rekha A, Ravishankar KV. Evaluation and characterization of EMS derived mutant populations in mango. Sci Hortic 2019;254:55-60. [CrossRef]
41. Kaur N, Mittal RK, Sood VK, Soharu A. Studies on induced chlorophyll mutants in black gram (Vigna mungo L. Hepper). Himachal J Agric Res 2022;47:156-62.
42. Kim SH, Kim SW, Ahn JW, Ryu J, Kwon SJ, Kang BC, et al. Frequency, spectrum, and stability of leaf mutants induced by diverse g-ray treatments in two Cymbidium hybrids. Plants (Basel) 2020;9:546. [CrossRef]