
1. INTRODUCTION
Citrullus lanatus (Thunb) Matsum and Nakai, also known as watermelon, is a tropical fruit of the Cucurbitaceae family [1]. It is a monoecious warm-season crop that grows in a trailing and prostatic manner [2]. Watermelon fruits contain thick skin (exocarp) with varying pigmentation, a solid or striped look, a fleshy mesocarp, and an endocarp that can range in color from white to yellow [3,4]. The crop performs best in regions with year-round high temperatures in general [5,6].
The demand for Watermelon and its derivatives is on the rise due to its refreshing taste, high nutritional value, and wide culinary applications [7]. Watermelons are excellent sources of carotenoids, Vitamins A, B6, C, lycopene, and antioxidants [8-10]. The frequent inclusion of watermelon in a diet schedule can help to prevent the development of cancerous cells which could be implicated in the relative abundance of lycopene in watermelon fruits [8].
Despite the overwhelming nutritional and medicinal benefits of watermelons, little effort has been made to grow the crop in the derived savanna agroecology of southeastern Nigeria. The bulk of its production is concentrated in the drier savanna agroecological zones of Northern Nigeria, from where it is transported to other parts of the country [7,11-13]. This situation has led to the increased cost of watermelon in southern Nigeria.
Few attempts to grow the crop in the derived savanna agroecology of southeast Nigeria have been successful [7,14], although yield and quality have been significantly reduced by fungal diseases, with wilt disease the most predominant within the agroecology. Watermelon production is often limited by biotic and abiotic factors, including fungi pathogens. Fungal diseases constitute one of the most limiting biotic factors in watermelon production globally, and could significantly reduce yield and quality, and eventually cause significant economic losses to the farmer [15-19].
To combat these fungal diseases and ensure optimal watermelon production, farmers commonly employ synthetic fungicides. Fungicides commonly act by inhibiting the growth and reproduction of fungi, thereby minimizing disease progression. However, the efficacy of a fungicide may vary depending on several factors, including fungi species, the stage of development, and the fungicide spraying regime. Different varieties could respond differently to fungal pathogens due to varying levels of tolerance. Some varieties may possess genes that confer resistance to specific fungal pathogens, thus reducing the need for extensive fungicide use. Understanding the interaction between variety selection and fungicide spraying regimes on watermelon performance is crucial for optimizing watermelon production and disease management.
Previous reports on disease management in the cucurbits and in particular watermelon production, had largely focused on the efficacy of fungicides applications and doses of fungicides application on the management of Podosphaera xanthii, the causative organism of powdery mildew [16-18,20-26]. Studies involving varietal response to fungicide spraying regimes are scarce in the literature. The study hypothesized that watermelon varieties could respond differently to varying fungicide spraying regimes in the management of Fusarium wilt. Hence, the objectives of the study were: To assess the effect of fungicide spraying regimes on disease incidence and severity, growth, and yield components of three varieties of watermelon; and to identify and characterize the fungal causal organism of Fusarium wilt of watermelon in the derived savanna agroecology of southeast Nigeria.
2. MATERIALS AND METHODS
2.1. Experimental Site
The experiment was conducted at the Teaching and Research Farm, and the Pathology Laboratory of the Department of Crop Science, University of Nigeria, Nsukka, between September and November 2022. The location is between latitude 6° 86´ North and longitude 7° 40’ East at an altitude of about 450 m above sea level in South-eastern Nigeria.
2.2. Experimental Design and Treatments
The experiment was laid out as a 3 × 4 factorial in a randomized complete block design with two factors A and B. Factor A comprised the three varieties of watermelon (Red Diamond, Sweet Sangaria, and Jubaly), while factor B was the four fungicide spraying regimes (weekly, bi-weekly, tri-weekly and control). The three varieties of watermelon were sourced from the Department of Crop Science Seed Bank, University of Nigeria, Nsukka. Fungiforce Gold fungicide (Metalaxyl 8% + manxozeb 64% WP) and Laraforce Gold insecticide (Lambda-cyhalothrin 5% + imidacloprid 15% SC) were used for fungi and insect management. Poultry manure was obtained from the Department of Animal Science Research Farm, University of Nigeria Nsukka (UNN).
2.3. Land Preparation and Planting
Land measuring 23 m by 15 m was used for the experiment. The field was cleared, tilled, and partitioned into three blocks separated by a 1 m pathway. Each block consisted of 12 plots of size 6 m2. A blanket dose of 10 t/ha poultry manure was applied 2 weeks before planting. Two seeds were sown per stand at a plant spacing of 1 × 1 m, which was eventually thinned down to 1 at 2 weeks after planting (WAP). Laraforce Gold insecticide (Lambda-cyhalothrin 5% + imidacloprid 15% SC) was sprayed to control cutworms 1 week after planting. Scarecrows and guard rows were made to prevent attack by guinea fowl or other rodents.
2.4. Spraying Regime
The spraying of Fungiforce Gold fungicide (Metalaxyl 8% + manxozeb 64% WP) started exactly 3 WAP and was sprayed on every plot except the control. Hence, the weekly plots were sprayed a total of nine times, bi-weekly plots a total of five times, and tri-weekly plots a total of three times, covering a 9-week spraying schedule.
2.5. Data Collection
Data were collected weekly on days to 1st, 50, and 100% emergence (DTE), days to 1st, 50, and 100% flowering (DTF), number of nodes (NN) per plant, length of vine, and fruit weight from 3 sample plants per plot. Disease incidence and severity were recorded on a per-plot basis.
2.6. Determination of Phenology, Growth, Yield, and Disease Parameters
All growth and yield traits were determined according to the procedures used by [7]. Disease incidence for each plot was determined by counting the total number of plants sampled and the number of plants showing symptoms and calculated using the formula below:
 |
Scoring of disease severity was done using a 5-point scale at the onset of fruiting [27].
Severity Estimation (%) Scale Interpretation
0 No infection
1 Slight infection
2 Moderate infection
3 Severe infection
4 Completely infected and death.
 |
2.7. Morphological and Molecular Characterization of Fungi Isolates
Leaf samples were randomly selected from each treatment and properly cleaned in sterile water. Using flame-sterilized forceps, the diseased tissues were divided into small pieces of 2–5 mm squares and put into sterile Petri dishes containing a 0.1% mercuric chloride solution.
Potato dextrose agar medium comprising 200 g of peeled potatoes, 20 g of dextrose, 20 g of agar, and 1000 mL of water was prepared and sterilized in an autoclave at 121°C for 15 min at 15 psi. The tissue pieces were surface sterilized using 1% sodium hypochlorite solution for 30–60 s before rinsing twice or three times in sterile water. Three tissue pieces per plate were aseptically added to Petri dishes with a nutritional medium (potato dextrose agar medium) that was treated with streptomycin sulfate. The plates were incubated between 25 and 27°C, or room temperature [28]. Based on colony and physical traits including color and form, isolated fungi were identified. The Mycological Atlas of Robert and Ellen was used to confirm and authenticate the morphological traits and appearance of the fungal isolations.
Fungal staining was prepared for slide cultures. Cover slip-sized PDA agar blocks were cut and set on a clean microscopic slide over a sterile “V-shaped” glass rod in a Petri dish. The fungus was then inoculated along the margins of the agar block. The agar was covered with a sterile coverslip, and the apparatus was then incubated at room temperature. After the fungus had grown, the coverslip was carefully removed, stained with lactophenol blue, and examined using an Olympus bright field microscope (Model BX43) equipped with a 10- and 40-objective lens.
DNA was extracted using the Zymo Research Mini Quick-DNA™ Fungal/Bacterial Miniprep Purification Kit, with catalog number D6005. DNA Amplification and Agarose gel electrophoresis. The isolate’s ITS region was amplified in a MiniAmp plus Thermal Cycler (Thermo Fisher Scientific) using the fungi universal oligonucleotide primers. The polymerase chain reaction (PCR) mixture contained 12.5 μL of 1X Master mix with standard buffer, 0.5 μL (10 μM) of each of the forward and reverse primers, 3 μL of the extracted DNA, and 8.5 μL of sterile nuclease-free water to make up to 25 μL of reaction volume. One Taq Quick load 2X Master Mix with Standard Buffer (New England Biolabs, MA, USA) was used for the PCR, which is composed of 20 mM Tris-HCl, 1.8 mM MgCl2, 22 mM NH4Cl, 22 mM KCl, 0.2 mM DNTPS, 5% glycerol, 0.06% IGEPAL CA-630, 0.05% Tween 20, Xylene Cyanol FF, Tartrazine, and 25 units/mL of Taq DNA polymerase. This was vortexed at low speed and placed in a thermal cycler machine [Table 1] [29].
2.8. Cycling Parameters and Primers Used
Two primers ITS-1 (TCCGTAGGTGAACCTGCGG), and ITS-4 (TCCTCCGCTTATTGATATGC) were used. Initial denaturation was done at 95°C for 2 min, and then at 94°C for 30 s. Annealing was done at 55°C for 2 min, extended at 72°C for 1 min, and finally extended at 72°C for 10 min. This process was repeated for 30 cycles.
The PCR products were examined on a 1.5% Agarose gel stained with ethidium bromide (1 g/mL), electrophoresed at 110 volts for 45 min, and then seen under a UV transilluminator. As a DNA molecular weight marker, a 100 bp DNA ladder (New England Biolabs, USA) was employed.
The BigDye Terminator v3.1 cycle sequencing kit was used for sequencing the amplified fragments on an Applied Biosystems Genetic Analyzer 3130 × l sequencer per the manufacturer’s instructions. FinchTv software was used to modify the resulting FASTA format sequences [30].
3. RESULTS
3.1. Effect of Variety and Spraying Regime on the Phenology of Watermelon
The effect of variety and spraying regime on the phenological attributes of watermelon is shown in Figure 1. The result showed that there was a significant difference (P < 0.05) in the varietal effect on phenological traits. Jubaly had the earliest days to 1% emergence (DTE-1%) at 4 days, while sweet sangaria was earliest in DTE-50% (4.5) and DTE-100% (6.67). In contrast, the red diamond was the latest in DTE-1% (5.67), DTE-50% (7.25), and DTE-100% (11.00). Jubaly variety also showed superiority in earliness to flowering with 28.00, 32.83, and 35.75 DTF, which was the earliest compared to red diamond (31.59 DTF-1%) and sweet sangaria (36.83 DTF-50%, 40.00 DTF-100%) which took longer days.
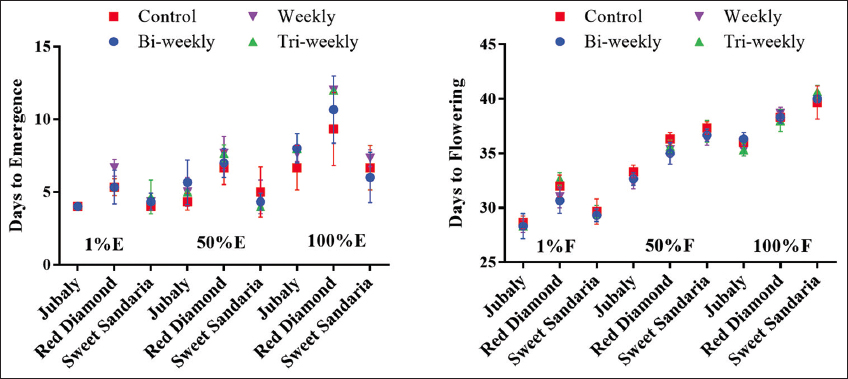 | Figure 1: Effect of variety and spraying regimes on the phenology of watermelon. E: Emergence; F: Flowering. The graph shows the mean values and standard deviations of 3 replicates. [Click here to view] |
The spraying regime significantly affected (P < 0.05) days to 50% flowering but not DTF-1% and DTF-100%. Weekly (34.67) and bi-weekly spray regimes (34.56) influenced shorter DTF-50% compared to tri-weekly (35.22) and control (35.67). Bi-weekly (29.44) and weekly (29.55) spray also showed a tendency for earliness in DTF-1% compared to tri-weekly (30.22) and control (30.11) spray regimes. However, this trend was not replicated in DTF-100, as all spraying regimes took an average of 40 days to 100% flowering [Figure 1].
The response of watermelon varieties to fungicide spraying regimes was similar (P > 0.05) in all phenological attributes studied. Varieties responded better to weekly and bi-weekly spraying regimes in DTF compared to tri-weekly and no-spray regimes, respectively. The interaction of Jubaly variety x weekly, bi-weekly, and tri-weekly spray regimes gave comparable results (28.33) in DTF-1% and was the earliest in contrast to the interaction of red diamond variety x tri-weekly (32.67) spray regime, which was the latest. Jubaly variety showed consistency for early flowering across all spraying regimes, flowering earlier than red diamond and sweet sangaria. The interaction of Jubaly variety x weekly spraying regime was earliest in attainment of DTF-50% (32.33), and DTF-100% (35.33) in contrast to the interactions of sweet sangaria variety x no-spray, and sweet sangaria variety x tri-weekly spraying regime, which were latest to attainment of DTF-50% (37.33) and DTF-100% (40.67), respectively [Figure 1].
3.2. Effect of Variety and Spraying Regimes on Growth Parameters of Watermelon
The effect of variety on the vine length of watermelon was significant (P < 0.05) across the weeks of investigation. Jubaly variety produced significantly longer vines (8.00) at 2 WAP, 4 WAP (34.86), 6 WAP (110.60), and 8 WAP (154.80) compared to the red diamond variety (6.59) which produced the shortest vine at 2 WAP, and sweet sangaria variety which consistently gave shorter vines at 4 WAP (27.43), 6 WAP (81.73), and 8 WAP (119.05) [Figure 2].
 | Figure 2: Effect of variety and spraying regimes on vine length of watermelon. The graph shows the mean values and standard deviations of 3 replicates. Bars with different letters are significantly different at P<0.05. [Click here to view] |
The spraying regime did not significantly affect (P > 0.05) the vine length of watermelon, as all spraying regimes gave comparable results. However, the bi-weekly spraying regime recorded longer vines at 2 WAP (7.55) and 4 WAP (30.90) compared to the tri-weekly spraying regime, which had the shortest vines at 2 WAP (6.8) and 4 WAP (28.31). At the later stage of the plant development, the weekly spraying regime recorded longer vine lengths at 6 WAP (94.08) and 8 WAP (151.94), while the control had the shortest vines with 91.09 and 116.79, respectively [Figure 2].
The different varieties showed similarities in their response to fungicide spraying regimes. Jubaly variety showed the best response to spraying regimes across the weeks of study. The interaction of Jubaly x weekly spraying regime recorded the longest vine at 2, 4, 6, and 8 WAP with 8.52, 38.22, 120.51, and 183.44, respectively. In contrast, it was the interaction of red diamond x tri-weekly (6.14), red diamond x no-spray (26.70), sweet sangaria x weekly (74.63), and red diamond x no-spray (102.33) that recorded the shortest vines at 2, 4, 6, and 8 WAP respectively [Figure 2].
3.3. Effect of Variety and Spraying Regimes on Node Number of Watermelon
The varieties differed significantly (P < 0.05) in NN at 6 and 8 WAP [Figure 3]. There was no significant difference (P > 0.05) in NN at 4 WAP. Jubaly variety had higher NN (27.22) at 6 WAP, and at 8 WAP (38.11) in comparison with the sweet sangaria variety consistently had the least NN (22.17) at 6 WAP and (33.14) at 8 WAP.
 | Figure 3: Effect of variety and spraying regimes on number of nodes of watermelon. The graph shows the mean values and standard deviations of 3 replicates. Bars with different letters are significantly different at P<0.05. [Click here to view] |
The four spraying regimes influenced comparable (P > 0.05) NN. However, bi-weekly (24.93) and weekly (38.52) spraying regimes recorded higher NN at 6 and 8 WAP, respectively, than the no-spray treatment, which was consistent in recording the least NN at 6 WAP (23.56) and 8 WAP (34.13) [Figure 3].
The interaction effect of variety and spraying regime on NN was comparable at 4, 6, and 8 WAP [Figure 3]. The interaction of Jubaly x weekly spraying regime was also consistent in affecting the highest NN at 6 WAP (29.22) and 8 WAP (44.89). In contrast, the interaction of red diamond variety x no-spray had the least NN at 6 WAP (20.89) and 8 WAP (32.00). The varieties showed negligible differences from the four spraying regimes at 4 WAP.
3.4. Effect of Variety and Spraying Regimes on Yield Parameters of Watermelon
The effect of variety and spraying regime on fruit yield and yield traits of watermelons are shown in Figure 4. There was a clear varietal effect (P < 0.05) on fruit number and fruit weight, but not on fruit length and fruit width. The Jubaly variety was superior in fruit number (9.58) and fruit yield (38.04 t/ha) compared to the red diamond variety, which was the least in fruit number (6.42) and fruit yield (26.09 t/ha). There was no clear-cut varietal effect on fruit length and fruit width as all varieties showed comparable values.
 | Figure 4: Effect of variety and spraying regimes on fruit yield and yield components of watermelon. The graph shows the mean values and standard deviations of 3 replicates. Bars with different letters are significantly different at P<0.05. [Click here to view] |
Although a significant difference was not detected among spraying regimes in all yield parameters, weekly spray recorded higher fruit number (8.56) and fruit weight (48.21 t/ha) than no-spray with 7.00 and 35.09 t/ha mean fruit number and fruit weight, respectively. Fruit length and fruit width were similar in all four spraying regimes [Figure 4].
The interaction of variety and fungicide spraying regime did not significantly affect fruit yield and yield attributes of watermelon. However, the varieties followed a definite trend in their response to the fungicide spraying regime.
The interaction of Jubaly variety x weekly spray consistently recorded higher fruit number (11.00 cm), fruit width (22.37 cm), and fruit yield (48.11 t/ha) than the interaction of red diamond variety x control, which recorded the least values in fruit number (5.53), fruit width (19.20), and fruit yield (24.52). The interaction effect of red diamond variety x weekly spray produced the longest fruit (14.37 cm) compared to Jubaly x control, which was the least (9.42 cm) [Figure 4].
3.5. Effect of Variety and Fungicide Spraying Regime on Disease Incidence and Severity of Fusarium wilt of Watermelon
The effect of variety and fungicide spraying regime on disease incidence of Fusarium wilt of watermelon is shown in Figure 5. The varieties showed differences in their tolerance level of Fusarium wilt of watermelon. Red diamond showed higher tolerance to the disease by recording the least percentage incidence at 9 WAP (25%), 10 WAP (18%), and 11 WAP (30%) in contrast to the Jubaly variety, which showed higher susceptibility, recording the highest percentage incidences of 25%, 32%, and 35% at 9, 10, and 11 WAP, respectively.
 | Figure 5: Effect of variety and spraying regimes on disease incidence of watermelon. The graph shows the mean values and standard deviations of 3 replicates. Bars with different letters are significantly different at P<0.05. [Click here to view] |
Weekly and bi-weekly sprays showed similar effects on the incidence of Fusarium wilt recording the least percentage incidence (22%) at 9 WAP. However, at 10 and 11 WAP, the weekly spraying regime was more effective, recording the least percentage incidences of 28% and 55% compared to control with the highest incidences of 83% and 94%, respectively. In general, the varieties showed a higher preference for weekly spray, than bi-weekly, tri-weekly, and no-spray, respectively [Figure 5].
The effect of variety and fungicide spraying regime on the severity of Fusarium wilt of watermelon is shown in Figure 6. Disease severity followed the same trend as was observed in disease incidence with the red diamond variety showing the least severity score compared to the Jubaly variety which recorded the highest severity score [Figure 6]. The severity score was also least in plots that received weekly spray compared to plots that received bi-weekly, tri-weekly, and no-spray respectively.
 | Figure 6: Effect of variety and spraying regimes on disease severity of watermelon. The graph shows the mean values and standard deviations of 3 replicates. Bars with different letters are significantly different at P<0.05. [Click here to view] |
3.6. Morphological and Molecular Characterization of the Fungi Isolate
A pinkish coloration was observed from the mycelia growth in the petri dish, and when viewed under the light microscope, floccose-shaped colonies; smooth, cylindrical, and branched hyphae; oval-shaped microconidia, and curved macroconidia were observed [Plate 1]. The basic local alignment search tool (BLAST) result showed 100% identity with Fusarium equiseti strain WZ-98 on the NCBI gene bank. Phylogenetic analysis using molecular evolutionary genetic analysis (MEGA) showed a high pairwise identity of the Isolates with sequences obtained from Nigeria, Africa, and other parts of the world [Figure 7].
 | Plate 1: Morphology of Fusarium equiseti spores as viewed in the microscope. [Click here to view] |
 | Figure 7: Maximum likelihood tree showing the relationship between the UNN isolate and other Fusarium equiseti isolated from watermelon in other countries. [Click here to view] |
Pairwise sequence comparison using the sequence demarcation tool (SDT) showed high similarity between the UNN isolate with NCBI accession number ORO31853.1 with isolates from other countries when compared with similarity to the outgroup Neurospora crassa [Figure 8].
 | Figure 8: Pairwise sequence comparisons of UNN isolate sequences of Fusarium equiseti and related sequences. [Click here to view] |
4. DISCUSSION
The findings of this study revealed significant effect of crop variety on various aspects of watermelon growth and development. Specifically, the study showed that watermelon variety had a significant influence on phenological characteristics, such as the time it took for the plants to emerge from the soil and the days until flowering occurred. Notably, the Jubaly variety outperformed the sweet sangaria and red diamond varieties in both of these critical phenological traits. Early emergence and flowering are highly desirable attributes for plant breeders seeking early-maturing varieties, as they are often indicative of early maturity in the crop [31]. The varietal effect was also evident in growth and yield-related traits, including vine length, node count, fruit number, and overall fruit yield. Once again, the Jubaly variety consistently exhibited superior performance in all these growth and yield traits when compared to the red diamond and sweet sangaria varieties. The longer vines [Figure 2] associated with the Jubaly variety resulted in an increased NN [Figure 3], ultimately translating into higher fruit production and greater fruit yield [Figure 4]. The observed variations in phenology, growth, and yield characteristics among the different varieties suggest underlying genetic differences and distinct ancestry among these watermelon varieties. These findings align with prior research conducted on watermelon and related crops by earlier researchers [32-35].
The effect of fungicide spraying regimes did not record any significant differences in phenology, growth, or yield components. However, plots subjected to weekly and bi-weekly fungicide spraying recorded marginally higher vine lengths, node counts, fruit numbers, and fruit weights. This effect could be attributed to the fungicide creating a more favorable growth environment by inhibiting the growth and proliferation of pathogenic fungi. Notably, the lowest disease incidence and severity were observed in plots subjected to weekly fungicide spray, in contrast to the untreated plots, which exhibited the highest disease values [Figures 5 and 6].
The study also recorded significant variation in the responses of various watermelon varieties to Fusarium wilt disease. The sweet sangaria variety in combination with a weekly spraying regime exhibited the lowest disease incidence and severity, while the interaction of the Jubaly variety with no fungicide spraying recorded the highest disease incidence and severity values.
Synthetic chemicals have been demonstrated as highly effective in controlling watermelon diseases, particularly when applied consistently to achieve the desired level of disease control. These chemicals are generally safe when applied in low concentrations [36,37]. It is advisable to select pesticides with low residual effects, typically having a half-life of <16 days, as they tend to accumulate less in the environment due to their shorter persistence [38]. Additionally, regulations such as pre-harvest intervals are commonly in place to ensure that crops or livestock products are not harvested immediately after treatment, allowing time for residue concentrations to decrease to safe levels before harvest. Fungicide application is known to inhibit or suppress the activities of phytopathogenic fungi, leading to lower disease incidence and severity, ultimately resulting in improved crop performance. Although statistical significance was observed primarily in the timing of flowering, the fungicide-treated plots consistently exhibited earlier flowering stages, including 1%, 50%, and 100% flowering, compared to the untreated plots. Moreover, the treated fields demonstrated better growth and yield attributes, aligning with the findings of previous studies on other crops like cucumber, tomatoes, and the control of diseases like blossom end rot [39-41].
Despite the Jubaly variety showing higher disease incidence and severity values, it still outperformed the other varieties across most growth and yield parameters evaluated. This could be attributed to inherent host immunity within the genetic makeup of the Jubaly variety [42]. Environmental factors, including relative humidity, temperature, and the saprophytic nature of the organism, may also have contributed to these outcomes which corroborates Aba et al. [39] and Dauda et al. [43] who had previously reported the incidence of Phytophthora blight of cucumber and Fusarium wilt of green pepper and radish, respectively, in the neighboring fields of the experimental plots. Since most solanaceous plants belong to the host range of the Fusarium pathogen, one could safely say that the source of inoculum is from the soil and that there is a disease build-up of the Fusarium pathogen in the environment.
Microscopic observations of F. equiseti’s morphological characteristics in this study were consistent with the findings of Hami et al. [44] who reported prevalence of Fusarium oxysporum, Fusarium solani, and F. equiseti in Kashmir, Northern Himalayas, supporting the validity of the identification. Furthermore, molecular characterization confirmed the presence of Fusarium spp., specifically F. equiseti, within the pathogen. This information is valuable for biodiversity assessment, taxonomic decisions, and pest management planning. The BLAST analysis showed a 100% identity match with F. equiseti strain WZ-98 in the NCBI gene bank. The Fusarium isolate from the UNN was submitted and deposited at the NCBI Gene Bank under the accession number OR031853.1.
Phylogenetic analysis was conducted to compare the UNN sequence with sequences isolated from watermelon in other countries. Using (MEGA version 11) and the SDT, high similarity was observed between the UNN isolates and other isolates [Figures 7 and 8]. Specifically, the UNN isolate formed a clade with other F. equiseti isolates. Although it did not fall within the same clade as the out group N. crassa, it is possible that the out group shares some ancestral lineage with other species, despite appearing more distant. This report is in tandem with the observations of Hami et al. [44] and Srimali et al. [45] in other Fusarium species. However, closely related species clustered together, consistent with bootstrap values. Utilizing the SDT (SDT version 1.2) for pairwise sequence comparison revealed a high pairwise identity of approximately 94–100% between the UNN isolates and other isolates, in contrast to the out group N. crassa, which exhibited only a 44% similarity.
Recent research has also uncovered the presence of various Fusarium species in diverse crop species. For example, in Northeastern Israel, Kalman et al. [46] identified four Fusarium species among isolates obtained from contaminated Allium cepa samples exhibiting symptoms of Basal Rot. Aba Saad et al. [47] observed significant variability among strains of Fusarium species isolated from a wide array of agricultural and ornamental crops in Lebanon; they reported the first-ever prevalence of several Fusarium species on Lebanese host plants. Rabaaoui et al. [48] identified numerous Fusarium strains from Date Palms (Phoenix dactylifera) isolates displaying symptoms of Leaf wilt and sudden decline syndrome in Tunisia. In addition, in a comprehensive survey conducted by Torbati et al. [49], approximately 80 Fungicolous Fusarium isolates were found to be associated with 36 host species and 32 fungal genera. The prevalent hosts were primarily categorized under the sub-kingdom Mucoromycota.
5. CONCLUSION
The findings of this research provide invaluable insights into the response of watermelon varieties to fungicide spraying regimes and identified F. equiseti as the pathogen causing the wilt disease. However, future research should be channeled toward addressing the limitations and expanding on the findings to develop more effective and sustainable strategies for managing Fusarium wilt in watermelon cultivation. For instance, additional isolates should be sequenced for a more thorough analysis to give a complete picture of the genetic diversity within the pathogen population. More research is also required to determine how different watermelon varieties respond to Fusarium wilt, as identification of tolerant varieties would grossly reduce the need for fungicide use. Research collaborations with colleagues from various institutions and locations can offer a more comprehensive view of Fusarium wilt and its control. This is true because sharing information and resources can result in studies that are more thorough and significant.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. FUNDING
This research did not receive any funding from public or private organizations.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All data generated in this study are included in the manuscript.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Dube J, Ddamulira G, Maphosa M. Watermelon production in Africa:Challenges and opportunities. Int J Veg Sci 2020;21:211-9. [CrossRef]
2. Boualem A, Lemhemdi A, Sari MA, Pignoly S, Troadec PC, Choucha FA, et al. The andromonoecious sex determination gene predates the separation of Cucumis and Citrullus genera. PLoS One 2016;11:e0155444. [CrossRef]
3. Bahari M, Rafii MY, Saleh GB, Latif MA. Combining ability analysis in complete diallel cross of watermelon (Citrullus lanatus (Thunb.) Matsum. and Nakai). ScientificWorldJournal 2012;2012:543158. [CrossRef]
4. Munisse P, Jensen BD, Andersen SB. Genetic differentiation of watermelon landraces in Mozambique using microsatellite markers. Afr J Biotechnol 2013;12:5513-21.
5. Uguru MI. Crop Production:Tools, Techniques, and Practice. Nsukka:Fulladu Publishing Company;1996. 4-15.
6. Lawal AB. Response of cucumber to intercropping with maize and varying rates of farmyard manure and inorganic fertilizer. Agric Environ 2000;2:78-83.
7. Ezeh IN, Ukwu UN, Uzoigwe DA. Comparative effects of two manure types and their combinations on growth and yield of watermelon (Citrullus lanatus (Thunb) Matsum and Nakai) in Nsukka, Southeast Nigeria. Asian J Adv Agric Res 2021;17:30-41. [CrossRef]
8. Guner N, Wehner TC. The genes of watermelon. HortScience 2004;39:1175-82. [CrossRef]
9. Jensen BD, Toure FA, Hamattal AG, NantouméAD. Watermelons in the sand of Sahara:Cultivation and use of indigenous landraces in the Tombouctou region of Mali. Ethnobotany Res Appl 2011;9:151-62. [CrossRef]
10. USDA NASS 2018. Available from:https://usda.mannlib.cornell.edu/usda/current/vegesumm/vegesumm-02-13-2018.pdf [Last accessed on 2023 Aug 14].
11. Schippers RR. African Indigenous Vegetables:An Overview of the Cultivated Species. London, UK:University of Greenwich, Natural Resources Institute;2020. 214.
12. Anikwe MA, Agu JC, Ikenganyia EE. Agronomic evaluation of four exotic tropical varieties of watermelon (Citrullus lanatus L.) in two agro-environments in Nigeria. Int J Plant Soil Sci 2016;10:1-10. [CrossRef]
13. Eifediyi EK, Remison SU, Ahamefule HE, Azeez KO, Fesobi PO. Performance of watermelon (Citrullus lanatus L) in response to organic and NPK fertilizer. Agric Environ 2017;9:5-18. [CrossRef]
14. Oraegbunam CJ, Njoku OM, Imoh ON, Obalum SE, Onyia VN, Atugwu AI, et al. Agronomic performance and adaptability of three varieties of watermelon (Citrullus lanatus) on sandy loam soil in derived savanna. Agro Sci J Trop Agric Food Environ Ext 2016;15:46-50. [CrossRef]
15. Keinath AP, DuBose VB. Evaluation of fungicides for prevention and management of powdery mildew on watermelon. Crop Prot 2004;23:35-42. [CrossRef]
16. Keinath AP, DuBose VB. Controlling powdery mildew on cucurbit rootstock seedlings in the greenhouse with fungicides and biofungicides. Crop Prot 2012;42:338-44. [CrossRef]
17. Kousik CS, Donahoo RS, Webster CG, Turechek WW, Adkins ST, Roberts PD. Outbreak of cucurbit powdery mildew on watermelon fruit caused by Podosphaera xanthii in Southwest Florida. Plant Dis 2011;95:1586. [CrossRef]
18. Kousik CS, Brusca J, Turechek WW. Diseases and disease management strategies take top research priority in the watermelon research and development group members survey (2014 to 2015). Plant Health Prog 2016;17:53-8. [CrossRef]
19. Reetu, Tomar M. Watermelon:A valuable horticultural crop with nutritional benefits. Popular Kheti 2017;5:5-9.
20. McGrath MT. Efficacy of fungicides with resistance risk for cucurbit powdery mildew and fungicide sensitivity of Podosphaera xanthii in New York. Phytopathology 2013;103:93.
21. Candido V, Campanelli G, Viggiani G, Lazzeri L, Ferrari V, Camele I. Melon yield response to the control of powdery mildew by environmentally friendly substances. Sci Hortic 2014;166:70-7. [CrossRef]
22. Cerkauskas RF, Ferguson G. Management of powdery mildew (Podosphaera xanthii) on greenhouse cucumber in Ontario. Can J Plant Pathol 2014;36:22-37. [CrossRef]
23. Kousik CS, Ikerd JL. Evidence for Cucurbit Powdery Mildew Pathogen Races based on Watermelon Differentials. In:Havey M, Weng Y, Day B, Grumet R, editors. Proceedings of Cucurbitaceae. Alexandria, VA:American Society of Horticultural Science;2014. 32-4.
24. Keinath AP. Efficacy of fungicides against powdery mildew on watermelon caused by Podosphaera xanthii. Crop Prot 2015;75:70-6. [CrossRef]
25. Barickman TC, Horgan TE, Wilson JC. Efficacy of fungicide applications and powdery mildew resistance in three pumpkin cultivars. Crop Prot 2017;101:90-4. [CrossRef]
26. Kousik CS, Ikerd JL, Mandal M. Relative susceptibility of commercial watermelon varieties to powdery mildew. Crop Prot 2019;125:104910. [CrossRef]
27. Nwufo MI, Ihejirika GO. Influence of inter-cropping and removal of diseased leaves on incidence and severity of leaf spot disease of Telfairia occidentalis Hook f. caused by Phoma sorghina. Life Sci J 2008;5:81-3.
28. De Wet MM, Brink HG. Fungi in the bioremediation of toxic effluents. In:Sharma VK, Shah MP, Parmar S, Kumar A, editors. Fungi Bio-prospects in Sustainable Agriculture, Environment and Nano-technology. Cambridge:Academic Press;2021. 407-31. [CrossRef]
29. Zymo Research. Zymo Research Instruction Manual 2023. Available from:https://files.zymoresearch.com/protocols/_d4068_d4069_quick-dna_miniprep_plus_kit.pdf [Last accessed on 2023 Aug 14].
30. BigDye Quick Reference 2016;BigDye™ Terminator v3.1 Cycle Sequencing Kit. Available from:https://assets.thermofisher.com/TFS-Assets/LSG/manuals/MAN0015666_BigDyeTerminator_V3_1_CycleSeq_QR.pdf [Last accessed on 2023 Aug 14].
31. Trachsel S, Burgueno J, Suarez EA, San Vicente FM, Rodriguez CS, Dhliwayo T. Interrelations among early vigor, flowering time, physiological maturity, and grain yield in tropical maize (Zea mays L.) under multiple abiotic stresses. Crop Sci 2017;57:229-42. [CrossRef]
32. Gichimu BM, Owuor BO, Dida MM. Agronomic performance of the three most popular commercial watermelon cultivars in Kenya as compared to one newly introduced cultivar and local landrace grown on dystric nitisols under sub-humid tropical conditions. ARPN J Agric Biol Sci 2008;3:65-70.
33. Kumar D, Sharma R. Effect of mulching on growth, yield and quality in different varieties of summer squash (Cucurbita pepo L.). Int J Curr Microbiol Appl Sci 2018;7:2113-9. [CrossRef]
34. Ukwu NU, Olasanmi B. Crossability among five cassava (Manihot esculenta Crantz) varieties. Mod Concepts Dev Agron 2018;2:1-6. [CrossRef]
35. Tegen H, Alemayehu M, Alemayehu G, Abate A, Amare T. Response of watermelon growth, yield, and quality to plant density and variety in Northwest Ethiopia. Open Agric 2021;6:655-72. [CrossRef]
36. Lechenet M, Dessaint F, Py G, Makowski D, Munier-Jolain N. Reducing pesticide use while preserving crop productivity and profitability on arable farms. Nat Plants 2017;3:17008. [CrossRef]
37. Hellar H. Pesticides Residues in Sugarcane Plantations and Environment after Long-term Use. Kilimanjaro Region, Tanzania:The Case of TPC Ltd.;2002.
38. Parween T, Jan S, Mahmooduzzafar S, Fatma T, Siddiqui ZH. Selective effect of pesticides on plant--a review. Crit Rev Food Sci Nutr 2015;56:160-79. [CrossRef]
39. Aba S, Eze SC, Ishieze PU, Omeje TE, Ugwuoke KI. Controlling Phytophthora blight of cucumber (Cucumis sativus) in a derived savannah, using different spray intervals of a registered fungicide. Plant Pathol Quarantine 2018;8:122-30. [CrossRef]
40. Mtui HD, Maerere AP, Bennett MA, Sibuga KP. Effect of mulch and different fungicide spray regimes on yield of tomato (Solanum lycopersicum L.) in Tanzania. Afr J Food Agric Nutr Dev 2015;15:9607-19. [CrossRef]
41. Miller NF, Standish JR, Quesada-Ocampo LM. Sensitivity of Fusarium oxysporum f. sp. niveum to prothioconazole and pydiflumetofen in vitro and efficacy for Fusarium wilt management in watermelon. Plant Health Prog 2020;21:13-8. [CrossRef]
42. McCarville JL, Ayres JS. Disease tolerance:Concept and mechanism. Curr Opin Immunol 2018;50:88-93. [CrossRef]
43. Dauda N, Ukwu UN, Osadebe VO, Ishieze UP, Ugwuoke KI. Environmental Impact on Incidence and Severity of Fusarium Wilt of Radish (Raphinus sativus L.) in Nsukka. In:Proceedings of the 39th Annual Conference of the Horticultural Society of Nigeria. Ibadan:Cocoa Research Institute of Nigeria;2021. 156-62.
44. Hami A, Rasool RS, Khan NA, Mansoor S, Mir MA, Ahmed N, et al. Morpho-molecular identification and first report of Fusarium equiseti in causing chilli wilt from Kashmir (Northern Himalayas). Sci Rep 2021;11:3610. [CrossRef]
45. Srimali PU, Manamgoda DS, Basnayake BM, Thushari AN, Salim N. Molecular characterization and pathogenicity of Fusarium species associated with Pokkah Boeng:An emerging disease of sugarcane in Sri Lanka. Sugar Tech 2022;24:1357-67. [CrossRef]
46. Kalman B, Abraham D, Graph S, Perl-Treves R, Meller Harel Y, Degani O. Isolation and Identification of Fusarium spp., the causal agents of onion (Allium cepa) basal rot in Northeastern Israel. Biology (Basel) 2020;9:69. [CrossRef]
47. Aba Saad C, Masiello M, Habib W, Gerges E, Sanzani SM, Logrieco AF, et al. Diversity of Fusarium species isolated from symptomatic plants belonging to a wide range of agri-food and ornamental crops in Lebanon. J Fungi 2022;8:897. [CrossRef]
48. Rabaaoui A, Dall'Asta C, Righetti L, Susca A, Logrieco AF, Namsi A, et al. Phylogeny and mycotoxin profile of pathogenic Fusarium species isolated from sudden decline syndrome and leaf wilt symptoms on date palms (Phoenix dactylifera) in Tunisia. Toxins (Basel) 2021;13:463. [CrossRef]
49. Torbati M, Arzanlou M, da Silva Santos AC. Fungicolous Fusarium species:Ecology, diversity, isolation, and identification. Curr Microbiol 2021;78:2850-9. [CrossRef]