
1. INTRODUCTION
Orchids, renowned for their captivating beauty and remarkable ecological adaptations, are a diverse and enigmatic group of flowering plants. Among the thousands of orchid species inhabiting various ecosystems worldwide, the terrestrial orchid Malaxis rheedei, syn Malaxis versicolor stands out as an intriguing subject of study [1]. It is an endangered species with medicinal value [2]. This species, known for its distinctive morphology and intriguing ecological interactions, has always fascinated botanists, ecologists, and conservationists. It is a terrestrial or lithophytic erect species. The stems are sheathed, swollen at the base, and purplish green. The flowers of M. rheedei are maroon, bracts lanceolate, dorsal sepals linear, and petals recurved or incurved [1]. Its pseudobulbs are used as a tonic and form one of the ingredients of important ayurvedic medicine known as “Astavarga” which has therapeutic value [3].
M. rheedei is a species of orchid native to various parts of Asia, including India. It is endemic to Silent Valley, Mysore, Hassan, Thalgoppa (Kodagu), Tarave forests, Charmadi ghats, Kotigehar, Kemmangundi, Dakshina Kannada in India [4]. It engages a complex symbiotic relationship with mycorrhizal fungi beneath the forest canopy or within its preferred habitat as it is a known quality of terrestrial orchids [5]. This relationship is critical for its survival and reproductive success, making it a compelling model for understanding orchid–fungus interactions and the broader ecology of terrestrial [6].
The mycorrhizal association in orchids, including M. rheedei, is a striking example of mutualism, where the orchid and the associated mycorrhizal fungi exchange essential nutrients and support each other’s growth and reproduction [7]. Orchids depend on these fungal partners, especially for nutrient acquisition, as they often grow in nutrient-poor soils [8]. The most prevalent orchid mycorrhizal fungus in temperate and tropical areas are particularly members of the Tulasnellaceae family [9]. As a member of the fungal family Tulasnellaceae, Tulasnella calospora is known for its specific associations with orchids and its ability to facilitate their growth and development [10]. Tulasnella calospora is instrumental in assisting orchid seeds in germinating and developing into seedlings by providing the necessary nutrients and a suitable environment [11]. Several different consequences might result from the interaction of mycorrhiza with the orchid host. A post-embryonic plant structure called a mycorrhizal orchid protocorm, which has cells with intracellular fungi hyphae (pelotons) inhabited by them, is the result of a favorable association [12]. Usually, viable mycorrhizal protocorms grow into photosynthetic seedlings, but occasionally, they may become overtaken by the fungus’ mycelium and decayed hyphae [13].
Investigating the specific mycorrhizal associations with M. rheedei and understanding how these interactions influence the morphology of the roots of plants are a central focus of this study.
2. MATERIALS AND METHODS
2.1. Study Location
Roots of M. rheedei (Lindl.) orchid species were collected from two different areas in the Western Ghats. Collection was done in September to identify plants at vegetative and reproductive stages. Non-flowering plants were considered vegetative and flowering plants were considered at the reproductive state of plants. The characteristics of the location of the study are presented in Table 1.
Table 1: Characteristics of collection sites.
| S. No. | Collection sites | Latitude | Longitude | Vegetation type |
|---|---|---|---|---|
| 1. | Kogar, Shimoga | 14.1483 | 74.9581 | Evergreen and semi-evergreen climax forests |
| 2. | Z-point, Chikmagalur | 13.5474 | 75.7583 | Grassland slopes |
2.2. Sample Collection
Five individuals each attaining vegetative and reproductive growth stages were selected and their root samples were collected. When collecting, care was taken to avoid harming the roots. Roots were cleaned and kept until processing in a solution of FAA (formalin: 70% acetic acid: alcohol: 5:5:90).
2.3. Determination of Root Morphology
For the evaluation of root hair features, 10 randomly chosen root pieces (about 1 cm long) were put in water on microscope slides. ×20 was used to count the number of root hairs per centimeter of root length using a dissecting microscope. Using a compound microscope equipped with an ocular scale, measurements were taken at ×100 to determine the root hair’s length and width.
2.4. Estimation of Orchid Fungal Colonization
To check for fungal colonization in the cortical cells, the plant material was dyed with tryphan blue and examined under a compound microscope for the presence of the structure of fungi in the cortical cells. The magnified intersection method was used to calculate root length having mycorrhizal colonization [14]. In addition, the number of intersections having pelotons and fungal hyphae was counted. Root length having highly condensed hyphal pelotons (intact and digested) was selected through observation and their assessment was done through transmission electron microscopy [15]. The percentage of cells available for colonization was calculated by counting the number of colonized cells in the cortex divided by the total cells seen ×100.
Each section’s percentage of the cortical volume filled by pelotons was graded using a modified version of the eight-step scale (12.5–100%) developed by Rasmussen and Whigham [16]. A circular cover glass had crosshairs on it that divided the circumference into eight equal portions starting in the center. Eight portions of the circle were divided by 100%, making each portion a total of 12.5%. The divisions were divided into four pieces, each of 3.125%. Five hand sections taken from various root parts of a species were scored. The length and width of the 50 cells and pelotons in the chosen portions were measured.
2.5. Fungal Isolation and Purification
Mycorrhizal fungi were isolated in accordance with conventional methods known as plating method. The roots were surface sterilized on the suitable media. Roots were split into 5–10 mm pieces, in an aseptic manner and plated onto potato dextrose agar (PDA) medium, then allowed to set for 2 days at a temperature of 25°C. Fungi started developing from the chopped ends to produce the pure culture. These fungal isolates were subsequently grown on PDA at 25°C for 4 weeks in typical diurnal conditions before being tried for identification [17]. The isolated fungi were purified using the hyphal tip method.
2.6. DNA Isolation and Amplification
The CTAB technique was modified slightly to extract genomic DNA from the fungal mycelial. We used the universal primers (forward and reverse) for fungal ITS to carry out a polymerized chain reaction (PCR) to create many copies of a specific DNA. In the present study, Universal primers (forward and reverse) for fungal genome, ITS forward – (5’ – TCC GTA GGT GAA CCT GCC G – 3’), and ITS reverse – (5’ – TCC TCC GCT TAT TGA TAT GC – 3’) sequences were used. The ITS sections were amplified using PCR reaction mixes in a volume of 50 μL. Standard PCR settings were used to amplify the ITS sections of the fungal DNA [18].
2.7. Sequence Analysis
The chain termination approach was used to sequence DNA using a genetic analyzer from Applied Biosystems. For phylogenetic analysis, MEGA software version 6.0 was utilized, whereas ClustalW was employed for sequence alignments. A phylogenetic tree was created using the neighbor-joining (NJ) method. P-distance substitution model and bootstrapping value 1,000 were used for calculations [19].
2.8. Statistical Analysis
Mean and standard deviation were calculated to analyze mycorrhizal colonization and root hair morphology data. Unpaired t-tests were used to compare continuous variables that were reported. The analysis was conducted using SPSS version 22 statistical software. A 5% alpha level was regarded as significant.
3. RESULTS
3.1. Study of Root Characteristics
Roots of M. rheedei [Figure 1a] collected from Shimoga and Chikmagalur were analyzed thoroughly using a dissecting microscope, binocular compound microscope, and transverse electron microscope. Each centimeter of root length was observed under a dissecting microscope for the number of root hairs present in it. We noted 15.63 ± 1.12 and 16.14 ± 2.01 root hairs in Shimoga and Chikmagalur orchid samples, respectively. The root hair’s length and width were measured using a binocular compound microscope and noted. Root hair’s length was 92.45 ± 9.32 μm and 91.12 ± 9.98 μm at Shimoga and Chikmagalur locations, respectively. The breadth was noted at 10.24 ± 1.45 μm and 9.58 ± 2.15 μm, respectively [Figure 1b]. These values were not statistically significant (P < 0.05), which concludes that variation in location does not have a significant impact on root characteristics.
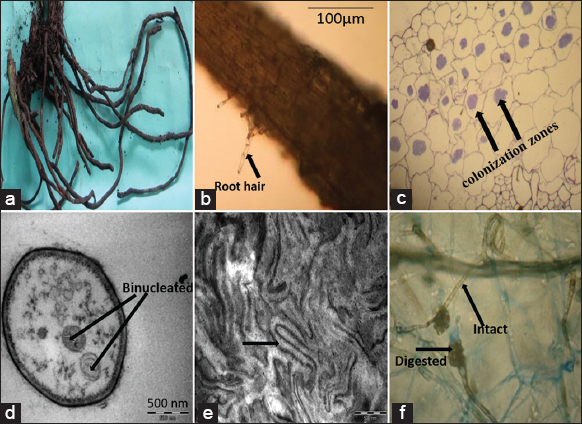 | Figure 1: Microscopic study of Entada rheedii roots. (a) Roots of M. rheedei; (b) transverse section of root and root hair; (c) colonization zone of fungal pelotons in cortex; (d) binucleated hyphae; (e) hyphae in the form of peloton (f) intact and digested pelotons. [Click here to view] |
3.2. Colonization of Roots
Fungal colonization was seen in the transverse section of the root cortical region. Root length with total colonization in the form of pelotons was approximately 91%. The diameter of the roots varied between 2 and 8 mm in the test species. The fungus’ hyphae commonly invade the roots through the root hairs or the root surface itself. Colonization was seen in the cortex of roots of all ages, both vegetative and reproductive. Hyphae appeared in the form of patches in the cortex [Figure 1c], taking up to 62% of the space [Table 2]. In reproductive age roots, some deformation of root hairs cortex appeared to be related to endophytes within their roots in the areas of colonized roots. We noted a 30% difference in root length between the degenerating peloton and the intact peloton [Table 2]. We also noted width of the vascular bundles was narrower than the parenchymatous cortex [Table 2].
Table 2: Root hair characteristics with and without pelotons.
| S. No. | Characteristics | Kogar, Shimoga | Z-point, Chikmagalur |
|---|---|---|---|
| 1. | Root hair number | 15.63±1.12 | 16.14±2.01 |
| 2. | Root hair length | 92.45±9.32 | 91.12±9.98 |
| 3. | Root hair breadth | 10.24±1.45 | 9.58±2.15 |
| 4. | Root length with degenerating peloton | 55.76±7.67 | 54.32±6.89 |
| 5. | Root length with the intact peloton | 39.67±7.89 | 38.41±6.55 |
| 6. | Root length with total colonization | 91.34±6.57 | 91.98±7.98 |
| 7. | Ratio of intact and lysed pelotons | 1.65±0.21 | 1.70±0.19 |
| 8. | Percent cortical volume occupied | 61.84±8.57 | 63.74±7.77 |
TEM studies noted that vegetative hypha was approximately 2.5–4 μm wide, hyaline, septate, binucleate [Figure 1d], constricted at branch points, sclerotia minute, mycelium composed of loosely arranged clusters of monoid cells which were hyaline, ellipsoidal to spherical, 7.5–14.2 × 6.5–10.8 μm, and in short branched or unbranched chains. In cells with degenerating peloton intact nuclei were seen.
TEM analysis also that M. rheedei root hairs did not remain straight and cylindrical and got distorted after the entry of fungal hyphae. Hence, we confirmed that the fungus entered into the roots only through the root hairs. Pelotons of loosely arranged clumps of hyphae were seen within the root’s cortical cells. Although the colonization of the fungal hyphae is a continual process, we could not see that all of the cortex’s cells were colonized at once. We also noted that colonization was mainly because of pelotonic hyphae [Figure 1e]. Pelotons were observed in the middle and inner cortex. The colonization density was up to 85% in the cortex. Both intact and digested pelotons were observed [Figure 1f]. The lysis of the peloton was from the periphery to the center of the cortical cell (centripetal lysis). Although 62% of the cortical volume was occupied with peloton, colonization was absent in the vascular region [Table 2].
3.3. Isolation of Fungi
The colony on PDA was pale white to cream, margin submerged, glabrous, with concentric zonation. The colony grew to 9 cm in diameter after 9 days of incubation. We yielded one fungal isolate from the roots of M. rheedei at both locations.
3.4. Molecular Characterization of Fungus
DNA was extracted and purified by the modified CTAB method from the fungal samples at both locations. The amplification success of loci, namely, ITS1-5.8S-ITS2 was 95%. ITS sequence (581 bp) data of the fungal isolate from the roots of M. rheedei showed a close (95%) resemblance to many Tulasnella species on NCBI BLAST. The sequence has been submitted to NCBI GenBank with Accession No. KC920478. Higher bootstrap values were seen in the NJ-based ITS sequencing tree, which was also associated with other Tulasnella species. The phylogenetic tree of the fungal isolate is given in Figure 2. It showed 98% identity with Tulasnella calospora, hence identified as T. calospora.
 | Figure 2: Phylogenetic tree of Tulasnella calospora isolated from the roots of Entada rheedii. [Click here to view] |
4. DISCUSSION
According to the fact that mature orchids are mycorrhizal dependent and frequently contain mycorrhizal roots or tubers [20], we collected M. rheedei from various sites in the current research to find differences in their morphology and mycorrhizal relationship. We found the same fungal association at both locations and non-significant variations in root morphology, it proves that the specificity of the orchid-mycorrhizal association is not location dependent.
Our study found distortion of root hairs of M. rheedei roots, which made it evident that the root hairs served as the main point of entrance for the fungus. This supports Sathiyadash et al. observations of eight terrestrial orchids that the fungal entrance into roots was always restricted to the portion of the roots called root hairs [14].
The fungus commonly penetrates the root hairs near their tips, occasionally just slightly away from the tips and infrequently at their bases. Root hairs often lose their straight cylindrical shape and become crooked and bent to varying degrees after the introduction of the fungal hyphae, imitating the pattern of colonization including the nodulating bacteria [21]. Depending on the location of the hyphal entrance, the alteration of the root hair may just reach the tip or may continue to various lengths in the direction of the base [22]. In the present study, the deformation of the root hairs cortex can be considered a sign of the presence of fungi.
Members of Tulasnellaceae are noted as the key mycorrhizal symbionts of several orchids such as Cypripedium, Chiloglottis Dendrobium nobile, Liparis japonica, and Ophrys [23-28]. The mycorrhizal status of the M. rheedei species is being examined in the present study for the first time. Phylogenetic analysis of isolated fungi found that T. calospora is associated with the medicinal orchid M. rheedei. Similar to our study, this fungus has been isolated, sequenced, and phylogenetically analyzed from many orchid species using ITS primers, such as Bipinnula fimbriata by Steinfort et al. [29], Cymbidium goeringii and C. faberi by Yu et al. [30], and Paphiopedilum barbigerum by Tian et al. [31]. A case study by Hampejsová et al. (2022) noted that this fungal species is responsible for the accumulation of certain disease–resistant molecules in orchid tubers, which might increase the value of the tubers for pharmaceutical and therapeutic uses [32].
5. CONCLUSION
The study concludes that M. rheedei exhibits a robust mycorrhizal association, with 91% root cortex colonization by the fungal hyphae. Phylogenetic analysis of isolated fungi identified an association of the fungus T. calospora with M. rheedei roots. The morphological analysis reveals that intact and degraded peloton patches occupy up to 62% of the cortical space, influencing root length. This study on mycorrhizal association may help conservationists in the conservation and restoration of orchid species. This information is also important for researchers to find novel ways for the cultivation, propagation, and identification of bioactive compounds. Future research on mycorrhizal associations in M. rheedei may focus on elucidating the molecular mechanisms governing the specificity of interactions between the orchid and T. calospora.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to the conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. FUNDING
There is no funding to the report.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. DATA AVAILABILITY
All data generated during the study is available in the article.
10. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
11. REFERENCES
1. Muthukumar T, Rajendran A, Priyadharsini P, Sarvalingam A. A new species of Malaxis (Orchidaceae:Epidendroideae) from the Southern Western Ghats, India. Webbia 2015;70:65-70. [CrossRef]
2. Renjini Haridas MS, Sindhu S, Thomas B. Potential bioactive components of Malaxis rheedii Sw. (Orchidaceae). Int J Exp Pharmacol 2016;6:11-5.
3. Haridas R, Thekkan S, Manorama S. Determination of total antioxidant activity of Malaxis rheedii Sw (Orchidaceae). J Pharmacogn Phytochem 2016;5:132-5.
4. Rao SK, Raja K, Swamy K, Kumar D, Singh RA, Bhat KG. Flora of Peninsular India;2019. Available from:https://flora-peninsula-indica.ces.iisc.ac.in/herbsheet.php [Last accessed on 2023 Sep 12].
5. Li T, Yang W, Wu S, Selosse MA, Gao J. Progress and prospects of mycorrhizal fungal diversity in orchids. Front Plant Sci 2021;12:646325. [CrossRef]
6. Hegde SM, Krishnaswamy K. Terrestrial orchid diversity in Sagara Taluk of Shimogga District central Western Ghats Region of Karnataka. J Plant Sci Res 2023;1:15-17.
7. Ma X, Kang J, Nontachaiyapoom S, Wen T, Hyde KD. Non-mycorrhizal endophytic fungi from orchids. Curr Sci 2015;10:72-87.
8. Rasmussen HN, Ras Rasmussen HN, Rasmussen FN. Orchid mycorrhiza:Implications of a mycophagous life style. Oikos 2009;118:334-45. [CrossRef]
9. Yang Q, Xu L, Xia W, Liang L, Bai X, Li L, et al. Mycorrhizal compatibility and germination-Promoting activity of Tulasnella species in two species of orchid (Cymbidium mannii and Epidendrum radicans). Horticulturae 2021;7:472. [CrossRef]
10. Brundrett MC. Understanding the roles of multifunctional mycorrhizal and endophytic fungi. In:Microbial Root Endophytes. Berlin, Heidelberg:Springer;2006. 281-98. [CrossRef]
11. Girlanda M, Segreto R, Cafasso D, Liebel HT, Rodda M, Ercole E, et al. Photosynthetic Mediterranean meadow orchids feature partial mycoheterotrophy and specific mycorrhizal associations. Am J Bot 2011;98:1148-63. [CrossRef]
12. Swarts ND, Dixon KW. Terrestrial orchid conservation in the age of extinction. Ann Bot 2009;104:543-56. [CrossRef]
13. Dearnaley JD, Martos F, Selosse MA. 12 Orchid mycorrhizas:Molecular ecology, physiology, evolution and conservation aspects. In:Fungal Associations. Berlin, Heidelberg:Springer;2012. 207-30. [CrossRef]
14. Sathiyadash K, Muthukumar T, Uma E, Pandey RR. Mycorrhizal association and morphology in orchids. J Plant Interact 2012;7:238-47. [CrossRef]
15. Ma X, Nontachaiyapoom S, Jayawardena RS, Gentekaki E, Zhou S, Qian Y, et al. Endophytic Colletotrichum species from Dendrobium spp. in China and northern Thailand. MycoKeys 2018;43:23-57. [CrossRef]
16. Rasmussen HN, Whigham DF. Phenology of roots and mycorrhiza in orchid species differing in phototrophic strategy. New Phytol 2002;154:797-807. [CrossRef]
17. Zhu GS, Yu ZN, Gui Y, Liu ZY. A novel technique for isolating orchid mycorrhizal fungi. Fungal Divers 2008;33:123.
18. Srivastava D, Manjunath K. DNA barcoding of endemic and endangered orchids of India:A molecular method of species identification. Pharmacogn Mag 2020;16:S290-9. [CrossRef]
19. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6:Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 2013;30:2725-9. [CrossRef]
20. Kartzinel TR, Trapnell DW, Shefferson RP. Highly diverse and spatially heterogeneous mycorrhizal symbiosis in a rare epiphyte is unrelated to broad biogeographic or environmental features. Mol Ecol 2013;22:5949-61. [CrossRef]
21. Kumar SS, Krishnamurthy KV. The role of root hairs in the mycorrhizal association of the ground orchid, Spathoglottis plicata blume. Int J Parasitol 1998;10:15-7.
22. Anjali P, Madhuri K, Sunita P. Terrestrial orchid mycorrhiza and non-mycorrhizal endophytes from Kolhapur District (MS)-III. Int J Life Sci 2016;A7:89-97.
23. Ding R, Chen XH, Zhang LJ, Yu XD, Qu B, Duan R, et al. Identity and specificity of Rhizoctonia-like fungi from different populations of Liparis japonica (Orchidaceae) in Northeast China. PLoS One 2014;9:e105573. [CrossRef]
24. Roche SA, Carter RJ, Peakall R, Smith LM, Whitehead MR, Linde CC. A narrow group of monophyletic Tulasnella (Tulasnellaceae) symbiont lineages are associated with multiple species of Chiloglottis (Orchidaceae):Implications for orchid diversity. Am J Bot 2010;97:1313-27. [CrossRef]
25. Schatz B, Geoffroy A, Dainat B, Bessière JM, Buatois B, Hossaert-qMcKey M, et al. Acase study of modified interactions with symbionts in a hybrid Mediterranean orchid. Am J Bot 2010;97:1278-88. [CrossRef]
26. Shefferson RP, Weiss M, Kull TI, Taylor DL. High specificity generally characterizes mycorrhizal association in rare lady's slipper orchids, genus Cypripedium. Mol Ecol 2005;14:613-26. [CrossRef]
27. Shefferson RP, Bunch W, Cowden CC, Lee YI, Kartzinel TR, Yukawa T, et al. Does evolutionary history determine specificity in broad ecological interactions?J Ecol 2019;107:1582-93. [CrossRef]
28. Xing X, Ma X, Men J, Chen Y, Guo S. Phylogenetic constrains on mycorrhizal specificity in eight Dendrobium (Orchidaceae) species. Sci China Life Sci 2017;60:536-44. [CrossRef]
29. Steinfort U, Verdugo G, Besoain X, Cisternas MA. Mycorrhizal association and symbiotic germination of the terrestrial orchid Bipinnula fimbriata (Poepp.) Johnst (Orchidaceae). Flora 2010;205:811-7. [CrossRef]
30. Yu Y, Cui YH, Hsiang T, Zeng ZQ, Yu ZH. Isolation and identification of endophytes from roots of Cymbidium goeringii and Cymbidium faberi (Orchidaceae). Nova Hedwigia 2015;101:57-64. [CrossRef]
31. Tian F, Liao XF, Wang LH, Bai XX, Yang YB, Luo ZQ, et al. Isolation and identification of beneficial orchid mycorrhizal fungi in Paphiopedilum barbigerum (Orchidaceae). Plant Signal Behav 2022;17:2005882. [CrossRef]
32. HampejsováR, Berka M, BerkováV, JersákováJ, DomkárováJ, von Rundstedt F, et al. Interaction with fungi promotes the accumulation of specific defense molecules in orchid tubers and may increase the value of tubers for biotechnological and medicinal applications:The case study of interaction between Dactylorhiza sp. and Tulasnella calospora. Front Plant Sci 2022;13:757852. [CrossRef]