
1. INTRODUCTION
Stevia rebaudiana Bertoni is a natural sweetener commonly known as a honey leaf, candy leaf or sweet leaf by the local people of Paraguay [1]. It is an economically important plant in medicine, food, and beverage industries [2], used to sweeten tea and herbal teas in different countries including Morocco [3]. It is an herbaceous native to Paraguay and grown commercially in many parts of the world [4]. In China, 20 000 ha are covered under stevia cultivation (the first producer in the world), while the chief market is in Japan (2000 tons of dry leaves/an) [5]. This plant is the best alternative to cane or beet sugar [6] and has numerous biological activities including anti-hyperglycemic, anti-inflammatory, hypotensive, antitumor, and antimicrobial [7]. It is a valuable medicinal plant of the family Asteraceae [8,9], producing zero-calorie diterpene glycosides in its leaves [10], which are widely used as a substitute for sugar since they are about 300 times sweeter than sucrose [4]. More than 38 steviol glycosides were detected on stevia leaves [11]. Stevioside is the most abundant compound in leaves (10–12% in dry weight) [12], while rebaudioside A is the sweetest and devoid of aftertaste [13]. Propagation of new varieties of stevia with higher Reb-A content is the overriding concern of many research groups [14]. However, the purpose of stevia selection is to obtain maximum quantities of plants, in a certain area with a relatively high content of sweetened substances [15]. Therefore, an effective protocol of mass production is very necessary for the cultivation of Stevia [14].
Conventional propagation of S. rebaudiana occurs by cuttings and seeds [16]. Propagation by seeds promotes heterogeneity in Stevia resulting in variability in the chemical composition and content of steviol glycosides, by modifying their quality, quantity, and growth [17]. Furthermore, due to self-incompatibility, seed-grown plants present heterogeneous phenotypes and low germination rates [18]. In addition, vegetative propagation through stem cutting has become unproductive, due to the unavailability of sufficient plant material in all seasons, and at the appropriate physiological stage, which limited the number of plants, that can be obtained [15]. Due to the above-mentioned difficulties, plant tissue culture provides a very useful and efficient technique for the mass production of S. rebaudiana [7] since it is able of producing a large number of plants that are similar in composition and steviol glycoside content [19,20], uniform, disease-free, and true-to-type at a fast rate. Stevia is previously micropropagated with direct organogenesis, through nodal segments [21], and shoot tip segments [22]. At present, in vitro culture of nodal explants is a viable alternative, for propagation of the species, but further standardization is required to achieve high success at field level [23]. Several elements, including genotype, explant type, medium composition, and culture circumstances, substantially encourage the efficacy of this approach. Consequently, a customized protocol must be developed for the genetic material that is available for specific regions [24]. This work aims to develop a rapid and efficient protocol for the mass production, of a selected plant of S. rebaudiana vigorous, and to improve its homogeneity through the techniques of in vitro multiplication by micropropagation of nodal explants on Murashige and Skoog (MS) medium supplemented with different plant growth regulators (PGRs).
2. MATERIALS AND METHODS
2.1. Plant Material and Culture Conditions
The experiment was carried out at the Regional Center for Agricultural Research in Kenitra (Morocco). Nodal explants were collected from two genotypes: genotype 1 (G1) and genotype 2 (G2) cultured in a greenhouse under controlled conditions at CCRA, INRA, Kenitra. They were thoroughly washed under running tap water, disinfected with 20% sodium hypochlorite solution (12°) for 25 min, and rinsed 3 times with distilled water. Then planted in culture tubes with solid MS medium (1962) containing 30 g/L of sugar, 7 g/L of agar and enriched with different combinations and concentrations of PGRs (Kin, 6-Benzylaminopurine [BAP], IAA) alone and in combination [Table 1]. The pH of the medium was adjusted to 5.8. Nodal segments (with a single axillary bud) of about 0.5–1 cm were cultured vertically in test tubes containing the different culture media incubated in a culture room with a temperature of 27 ± 2°C with 16 h of light and 8 h of darkness.
Table 1: MS medium supplemented with different concentrations and combinations of growth regulators.
| PGRs | PM0 | PM1 | PM2 | PM3 | PM4 | PM5 | PM6 | PM7 | PM8 | PM9 | PM10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| IAA (mg/L) | - | - | - | - | 0.5 | 0.5 | - | - | - | - | - |
| BAP (mg/L) | - | 0.5 | 1 | 2 | 1 | - | 1 | 1.5 | 2 | - | - |
| KIN (mg/L) | - | - | - | - | - | - | 0.25 | 0.25 | 0.25 | 0.25 | 0.5 |
2.2. Subcultures
After 4 weeks, the nodal segments of the regenerated shoots were subcultured in the same fresh proliferation medium (PM) for further multiple shoot induction. Subcultures were done every 4 weeks.
2.3. Root Induction
For rooting, individual shoots (more than three internodes) were implanted in vitro on MS basal medium with 0.5 mg/L of IAA or Naphtalenacetic acid (NAA) [Table 2].
Table 2: Rooting medium.
| PGRs | M1 | M2 | M3 |
|---|---|---|---|
| IAA (mg/L) | - | 0.5 | - |
| NAA (mg/L) | - | - | 0.5 |
NAA: Naphtalenacetic acid.
2.4. Acclimatization of Regenerated Plants
Rooted plants were washed with distilled water to eliminate the fixed medium. They were then moved to pots containing peat (60%) and sand (40%) and placed in mini greenhouses with 50–60% of relative humidity and 27 ± 2°C of temperature. Thus, the pots were covered with transparent plastic film for 2 weeks. The potted plants were transferred to the greenhouse for a better establishment for 1 month before being transplanted in the fields.
2.5. Statistical Analysis
Twenty cultures were raised for each treatment. The data obtained were processed using the “SAS” software. The comparison of the means relating to the responses of the explants was carried out using the Duncan test for all the treatments. The significance level of 5% is accepted for all analyses.
3. RESULTS
3.1. Shoots Multiplication and Proliferation
The statistical analysis shows that the PGRs combination had a significant effect on the number of days required for buds break and shoot emergence from the explant (P = 0.000) for both genotypes studied. Moreover, the buds started responding by buds break and the shoots appeared after 8 days of culture on PM0, PM9, and PM10 and after 12 days on the other mediums. Concerning the percentage of shoot induction, the difference is significant between both genotypes, and the highest percentage (G1 = 91.47% and G2 = 85%) was recorded in the PM0 followed by the medium containing 0.5 mg/L of Kin (PM10) with a percentage of 82% [Figure 1].
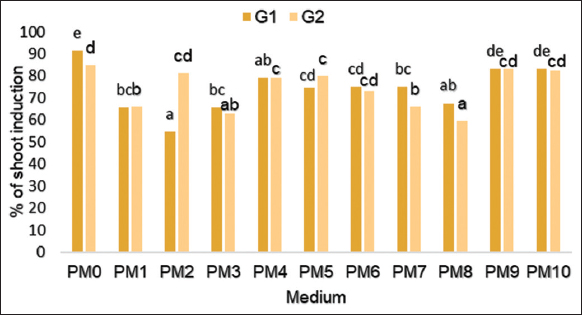 | Figure 1: Effect of different growth regulators on the percentage of shoot induction from the culture of nodal segments of Stevia rebaudiana in vitro. [Click here to view] |
The interaction between the PM and both genotypes for the number of shoots produced per explant is highly significant (P = 0.0015*). For G1, in vitro plants propagated on MP7 (MS+1.5 mg/L BAP + 0.25 mg/L Kin) medium has developed a maximum number of shoots per culture (2.9 shoots/explants), while for G2 a higher number of shoots (3.30 shoots/explants) was obtained on PM9 medium (MS + 0.25 mg/L Kin) [Figure 2].
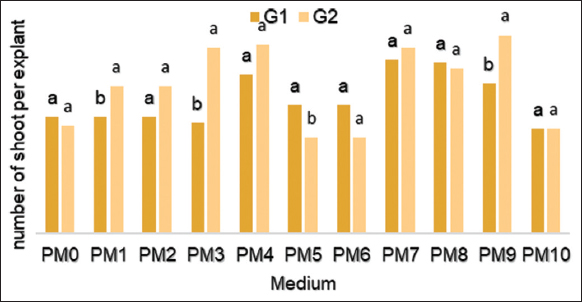 | Figure 2: Effect of different concentrations of hormones on the number of shoots/explants from the culture of nodal segments of Stevia rebaudiana. [Click here to view] |
The variance analysis showed that the length of new axillary shoots was significantly influenced by different combinations and concentrations of PGRs and by genotypes (P < 0.0001). The highest average length was recorded on PM0 and PM5 mediums [Figure 3] for both genotypes. The G1 grows more vigorously with an average length of 9.21; 9.22 cm respectively on the PM0 and PM5 media against 7.73; 6.5 cm for G 2.
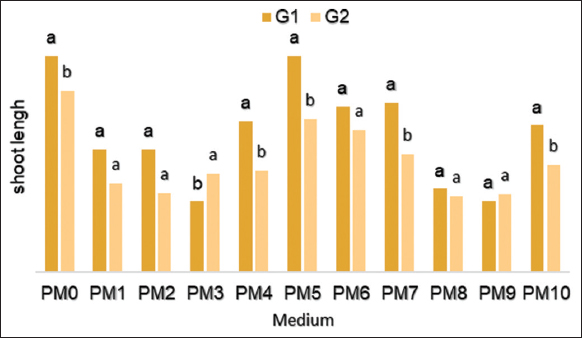 | Figure 3: Effect of different concentrations of hormones on the length of shoots of Stevia rebaudiana regenerated in vitro. [Click here to view] |
The number of internodes/vitro pousses was significantly impacted by the hormonal composition (P < 0.0001). While both cultivars showed no significant difference for this parameter. However, the highest number of internodes was found in the PM0 medium (5.75 internodes/explant for G1 and 4.70 for G2) [Figure 4]. In addition, the PM5 medium has favored the development of a large number of internodes for both genotypes, in contrast to the BAP-containing medium, which has regenerated the lowest number of internodes.
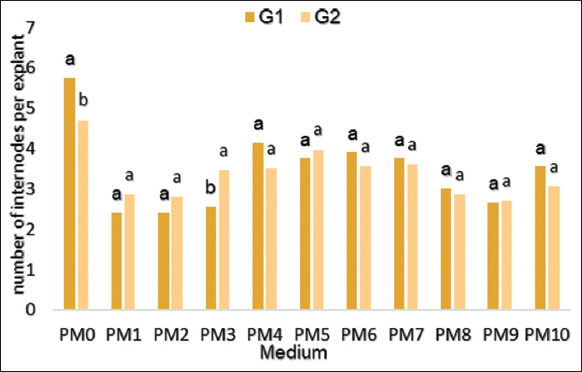 | Figure 4: Effect of different concentrations of hormones on the number of internodes per shoot of Stevia rebaudiana regenerated in vitro. [Click here to view] |
It should be mentioned that hormonal balance has influenced the appearance of regenerated shoots [Figure 5]. Hormone-free medium and mediums with IAA and Kin individually have regenerated relatively vigorous shoots with long internodal distances and large dark green leaves. As, BAP alone as well as in combination was found to induce a high number of short branching shoots with compacts, and green callus at the base [Table 3].
 | Figure 5: Morphological appearance of shoots of Stevia rebaudiana multiplied by buds break of nodal segments cultured on PM0, PM1, PM2, PM3, PM4, PM5, PM6, PM7, PM8, PM9, and PM10. [Click here to view] |
Table 3: Effect of different growth regulators on the formation of callus at the base of shoots obtained from the culture of the nodal segments of Stevia rebaudiana in vitro.
| Genotypes | MP0 | MP1 | MP2 | MP3 | MP4 | MP5 | MP6 | MP7 | MP8 | MP9 | MP10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| G 1 | - | +++ | +++ | +++ | ++ | + | ++ | ++ | ++ | + | + |
| G 2 | - | ++ | ++ | ++ | + | + | ++ | ++ | ++ | + | + |
-: No callus, +: Poor callus, ++: Medium callus, +++: Profuse callus.
3.2. Effect of Subcultures on Multiplication Rate
The multiplication rate, which represents the number of stems per explant and subculture after 4 weeks, was significantly influenced by different combinations and concentrations of PGR (P = 0.0001). Kin was more effective than IAA for shoot multiplication. The maximum mean number of shoots (2056.02 shoots/explant) occurred on PM10 medium (MS + 0.5 mg/L Kin) after the fourth subcultures. Furthermore, PM5 medium (MS + 0.5 mg/L IAA) has developed 1108.68 shoots/explants in the same period compared to the control who have regenerated only 764.55 shoots/explants [Table 4].
Table 4: Effect of subcultures on the number of shoots/explant.
| Medium | S1 | S2 | S3 | S4 |
|---|---|---|---|---|
| MP0 | ||||
| G1 | 6.45±0.28 | 36.83±0.65 | 179.34±1.46 | 764.55±1.07 |
| G2 | 4.66±1.88 | 18.37±0.53 | 84.52±2.54 | 339.66±0.26 |
| MP5 | ||||
| G1 | 5.83±0.7 | 34.83±0.92 | 233.91±0.7 | 1108.68±0.6 |
| G2 | 5.6±0.56 | 23.15±3.32 | 87.17±3.2 | 316.49±0.17 |
| MP10 | ||||
| G1 | 6.75±1.76 | 47.4±0.57 | 268.95±0.07 | 2056.02±0.02 |
| G2 | 4±0 | 16±0 | 72±2.82 | 366±0.14 |
S: Subculture.
Variations were detected in the morphology of shoots in terms of length and number of internodes depending on subcultures. A significant increase in the number of internodes and shoot length was observed from the first subculture in the PM5 and PM10 mediums compared to the Control (PM0), which experienced a decrease in both parameters. On the contrary, an improvement in leaf size, leaf color, and rooting capacity was detected in the MP0 medium. The incorporation of BAP into the medium has influenced the shortening of the shoots relative to control plants and induced callus formations in the cut portion of the shoot. These calluses inhibited the development of shoots from the first subculture. As a result, subcultures were not carried out for these media (PM1, PM2, PM3, PM4, PM6, PM7, PM8, and PM9).
3.3. Roots Formation
The results of the data analysis indicated that the rooting medium had a very significant effect on the percentage of root induction. M1 medium (control) has stimulated the fastest of root emergences (7 days) with a higher percentage of root induction for both genotypes (G1 97.6% and G2 81.87%) [Table 5]. This medium stimulated the appearance of the longest root mean for both genotypes (3.32 cm for G1 and 2.5 cm for G2) [Figure 6] from the basal part of the shoot with no intervening callus. Statistical analysis revealed that the type of auxin used did not significantly influence the average number of roots. The longest roots mean was 6.16 roots/vitro plants more stimulated by IAA (M2) and was not significantly different with control (M1) which regenerated 5.95 roots/plants. The application of NAA decreased the root induction, number of roots, and root length.
Table 5: Effect of IAA and NAA on the rooting of shoot of Stevia rebaudiana in vitro.
| Medium | Composition | % of root induction | Number of roots/shoot | Length of roots | Days for rooting | |||
|---|---|---|---|---|---|---|---|---|
| G 1 | G 2 | G 1 | G 2 | G 1 | G2 | |||
| M1 | Without hormones | 92.51±2.73c | 81.87±5.8c | 5.95±2.17 | 3.18±2.03 | 3.32±0.65b | 2.5±0.71b | 7 days |
| M2 | IAA (0.5 mg/L) | 83.99±5.52b | 68.78±8.55b | 6.16±1.83 | 3.94±1.83 | 2.48±0.84a | 1.78±1.37a | 10 days |
| M3 | NAA (0.5 mg/L) | 61.24±1.55a | 40.83±5.73a | 5.33±3.34 | 3.03±2.00 | 2.26±1.38a | 1.36±0.95a | 8 days |
Values are mean ± SD of twenty replicates. Means with the same letter are not significantly different from each other using Tukeys’ honest significant difference.
 | Figure 6: Effect of rooting medium on the quality of newly formed roots of vitro plants cultured on M1, M2, and M3 media. [Click here to view] |
Acclimatization and Establishment in the Field
Micro shots with well-developed roots have been successfully acclimatized under the conditions described above or with an acclimatization rate exceeding 93% for both genotypes [Figure 7]. For rapid acclimatization, the use of a porous substrate allowing drainage and adequate aeration was recommended. After acclimatization under the greenhouse, a perfect establishment of 90% of the plants in the natural terrain was completed.
 | Figure 7: (a) Acclimatization of rooted in vitro plants, (b) in vitro propagated Stevia plantlets ready for field transfer, and (c) establishment of in vitro grown plant in the field. [Click here to view] |
4. DISCUSSION
The combination BAP-Kin showed a better proliferation of shoots in this study. MP7 medium (MS + 1.5 mg/L BAP + 0.25 mg/L KIN) was favorable for the induction of a maximum number of shoots for G1. While for G2, the highest number of shoots was obtained on MS medium with 0.25 mg/L Kin. This variability between both cultivars may be due to genetic factors. Similarly, several authors have shown that the addition of cytokinins such as BAP and Kin to media has a stimulatory effect on shoot development [1,25,26] and auxins like IAA favored rhizogenic formations [24]. Our results have shown that the addition of IAA in the medium has induced both shoots and subsequent rooting.
BAP was reported to induce better shoot multiplication of Stevia in vitro in several studies. However, this cytokinin can affect the quality of in vitro regenerated plants. In our study, the incorporation of BAP in the medium has induced fairly, short, and branched shoots with compact callus at the base. On the contrary, the development of comparatively longer multiple shoots with large-size leaves and a higher number of internodes was obtained in plants grown on PM0 (without hormones) and PM5 (MS + 0.5 mg/L IAA). These results are in agreement with Ben Mahmoud et al. [5] who have stated that BAP-containing media gave stunted shoots with low acclimatization rates. Similarly, Al-Taweel [27] have found that BAP at 2.5 mg/L was more effective to encourage shoot proliferation, but these shoots were very thin and vitrified, while in vitro plants growing on control media were long with a higher number of leaves. The effectiveness of the control (hormone-free medium) compared with BAP and Kin for the development of shoots with significant length and a high number of leaves was also reported by Hassanen and Khalil [28].
It was clearly shown in this study, that MS medium with different types and concentrations of PGRs has succeeded inducing shoot proliferation from nodal explants of stevia. We have found that IAA and KIN were the most suitable PGRs for multiple shoot regeneration by continuous subcultures, and kinetin at 0.5 mg/L was more effective to increase the multiplication rate. However, cytokinins stimulate the cell division of plants and play a role in the release of lateral buds dormancy, lateral growth of shoots, and control of the cell cycle [29]. Several authors noted that KIN was more effective in stevia multiplication [30], while BAP was reported to induce less shoot proliferation and has stimulated calluses formation that has stopped the growth of shoots in repeated subcultures. These results are in agreement with Al-Taweel [27] who have demonstrated that the use of BAP has produced the maximum number of short, vitrified, and very thin shoots, irrelevant to subculture completion and with Ibrahim et al. [31] who have shown that the multiplication of S. rebaudiana without BAP is better for producing normal vitro plants. On the contrary, some researchers have observed that the addition of BA and NAA to the MS medium is more effective for the multiplication of axillary buds by repeated [32].
Regarding the rooting of stevia, our results showed that IAA at 0.5 mg/L induced a high mean number of roots (6.16 roots/shoots) with no significantly different from control (M1) which regenerated 5.95 roots/plants. The longest roots (3.32 cm) were developed on a hormone-free medium from the basal part of the shoot with no callus formation; these roots were good enough for their successful establishment in soil. These findings are in line with Hossain et al. [33] who reported that ½ MS media without any hormone is the best for shoot elongation. Several researchers [8,9,14,34] have mentioned the effectiveness of IAA as a rooting hormone. Ben Mahmoud et al. [5] noted that the vitro plants rooted on the MS medium with IAA showed a more interesting rhizogenic induction compared to the control and confirmed that the ability of plants to form roots on the MS medium without growth regulators could be attributed to the plant richness in endogenous auxins.
5. CONCLUSION
The successful tissue culture of S. rebaudiana affords a system that is efficient for the multiplication of this valuable medicinal plant. The best culture medium for multiplication and root formation was found and large-scale production of stevia from nodal segments was achieved. The protocol presented in this work allows the production of thousands of clonal plants from a nodal explant by continuous culturing shoot propagules over the course of 6 months. The multiplication rate achieved in this work is significantly high to be commercially significant for the mass production of S. rebaudiana under local conditions. Finally, the selection of new varieties of stevia with high levels of Rebaudioside A and without an aftertaste and their multiplication on a large scale according to this protocol is essential.
6. ACKNOWLEDGMENTS
The COSUMAR group in agreement with INRA, Kénitra, supported this research project. The authors gratefully acknowledge the COSUMAR group for having sanctioned the project for the micropropagation of S. rebaudiana in Morocco. The authors are also grateful to various individuals, in particular Dr. Abdelmjid Zouahri coordinator of the Research Unit of Environment and Conservation of natural resources INRA, CRRA of Rabat.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All data generated and analyzed are included within this article.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Asmono SL, Rahmawati, Sjamsijah N. The effect of murashige and skoog (MS) modified medium and several types of auxins on the growth of Stevia (Stevia rebaudiana Bertoni) in vitro. IOP Conf Ser Earth Environ Sci 2021;672:012001. [CrossRef]
2. Ghaheri M, Miraghaee S, Babaei A, Mohammadi B, Kahrizi D, Haghighi ZM, et al. Effect of Stevia rebaudiana Bertoni extract on sexual dysfunction in Streptozotocin-induced diabetic male rats. Cell Mol Biol (Noisy-le-grand) 2018;64:6-10. [CrossRef]
3. Khiraoui A, Bakha M, Amchra F, Ourouadi S, Boulli A, Al-Faiz C, et al. Nutritional and biochemical properties of natural sweeteners of six cultivars of Stevia rebaudiana Bertoni leaves grown in Morocco. J Mater Environ Sci 2017;8:1015-22.
4. Hossain MF, Islam MT, Islam MA, Akhtar S. Cultivation and uses of Stevia (Stevia rebaudiana Bertoni):A review. Afr J Food Agric Nutr Dev 2017;17:12745-57. [CrossRef]
5. Ben Mahmoud K, Ksaier M, Jemai N, Mehouachi T, Jemmali A study of the in vitro regeneration potential of an elite medicinal plant Stevia rebaudiana (Bert.). Ann l'INRA 2019;92:82-96.
6. Rokosa MT, Kulpa D. Micropropagation of Stevia rebaudiana plants. Ciên Rural 2019;50:e20181029. [CrossRef]
7. Ahmad A, Ali H, Khan H, Begam A, Khan S, Ali SS, et al. Effect of gibberellic acid on production of biomass, polyphenolics and steviol glycosides in adventitious root cultures of Stevia rebaudiana (Bert.). Plants (Basel) 2020;9:420. [CrossRef]
8. Ahmed MB, Salahin M, Karim R, Razvy MA, Hannan MM, Hossain M, et al. An efficient method for in vitro clonal propagation of a newly introduced sweetener plant (Stevia rebaudiana Bertoni.) in Bangladesh. Am Eur J Sci Res 2007;2:121-5.
9. Attaya AS. Direct shoot bud proliferation protocol from Stevia rebaudiana leaf culture for healthy biomass production. Egypt J Agron 2021;43:149-56. [CrossRef]
10. Akbari F, Arminian A, Kahrizi D, Fazeli A, Ghaheri M. Effect of nitrogen sources on gene expression of Stevia rebaudiana (Bertoni) under in vitro conditions. Cell Mol Biol (Noisy-le-grand) 2018;64:11-6. [CrossRef]
11. Libik-Konieczny M, Capecka E, Tuleja M, Konieczny R. Synthesis and production of steviol glycosides:Recent research trends and perspectives. Appl Microbiol Biotechnol 2021;105:3883-900. [CrossRef]
12. Verma R, Gena DD, Jat BL. In vitro propagation of Stevia rebaudiana Bertoni (a sweeting plant). World J Pharm Res 2016;5:666-85.
13. Pande SS, Gupta P. Plant tissue culture of Stevia rebaudiana (Bertoni):A review. J Pharmacogn Phytother 2013;5:26-33.
14. Yücesan B, Mohammed A, Büyükgöcmen R, Altug C,Kavas O, Gürel S, et al. In vitro and ex vitro propagation of Stevia rebaudiana Bertoni with high Rebaudioside-a content-a commercial-scale application. Sci Hortic2017;203:20-8.
15. Mehmed A, Enchev S, Bozhimirov S. Assessment of initial material for Stevia selection (Stevia rebaudiana B.). Bulg J Agric Sci 2021;27:536-40.
16. Tang CN, Lamasudin DU, Abdullah WM, Moo CL, Chiew MS, Chai QY, et al. Enhanced in vitro shoot regeneration and biochemical properties of Stevia rebaudiana using chitosan. Sains Malaysiana 2021;50:667-76. [CrossRef]
17. Al-Taweel SK, Azzam CR, Karim MR, Abel-Aziz RM, Abou-Sreea AI, Esmat EF, et al. Salinity effects on gene expression, morphological, and physio?biochemical responses of Stevia rebaudiana Bertoni in vitro. Plants (Basel)2021;10:820. [CrossRef]
18. Rasouli D, Werbrouck S, Maleki B, Jafary H, Schurdi-Levraud V. Elicitor-induced in vitro shoot multiplication and steviol glycosides production in Stevia rebaudiana. S Afr J Bot 2021;137:265-71. [CrossRef]
19. Luwa?ska A, Perz A, Ma?kowska G, Wielgus K. Application of in vitro Stevia (Stevia rebaudiana Bertoni) cultures in obtaining steviol glycoside rich material. Herb Pol 2015;61:50-63. [CrossRef]
20. Melviana AC, Esyanti RR, Setyobudi RH, Mel M, Adinurani PG, Burlakovs J. Gene expression related to steviol glycoside synthesis produced in Stevia rebaudiana (Bert.) shoot culture induced with high far-red LED light in TIS RITA®bioreactor system. Sarhad J Agric 2021;37:1-8. [CrossRef]
21. Fatima B, Nazir Z, Usman M, Bokhari SA, Shumaila. Enhanced micropropagation in Stevia rebaudiana Bertoni. Biologia 2021;67:34-9.
22. Bhingradiya V, Modi N, Mankad A. Tissue culture of Stevia rebaudiana (Bert.) using MS and S &H Medium. Int J Res Anal Rev 2019;6:1189-1193.
23. Singh M, Saharan V, Dayma J, Rajpurohit D, Sen Y, Sharma A. In vitro propagation of Stevia rebaudiana (Bertoni):An overview. Int J Curr Microbiol Appl Sci 2017;6:1010-22. [CrossRef]
24. Javed MA, Habib I, Jamil MW, Anwar JM, Nazir S, Rahman SU, et al. Protocol optimization for efficient in vitro micro-propagation of Stevia rebaudiana Bertoni. Adv Life Sci 2019;6:100-5.
25. Abdul Razak UN, Ong CB, Yu TS, Lau LK. In vitro micropropagation of Stevia rebaudiana Bertoni in Malaysia. Braz Arch Biol Technol 2014;57:23-8. [CrossRef]
26. Su YH, Liu YB, Zhang XS. Auxin-cytokinin interaction regulates meristem development. Mol Plant 2011;4:616-25. [CrossRef]
27. Al-Taweel SK, Azzam CR, Khaled KA, Abdel-Aziz RM. Improvement of Stevia (Stevia rebaudiana Bertoni) and steviol glycoside through traditional breeding and biotechnological approaches. Sabrao J Breed Genet 2021;53:88-111.
28. Hassanen SA, Khalil RM. Biotechnological studies for improving of Stevia (Stevia rebaudiana Bertoni) in vitro plantlets. Middle East J Sci Res 2013;14:93-106.
29. Amien S, Azhari AZ, Bakti, Maulana H. The effect of shoot explant types of eleven Stevia (Stevia rebaudiana Bertoni) accessions on in vitro growth. JAgro2021;8:142-52. [CrossRef]
30. Das N, Tripathi N, Basu S, Bose C, Maitra S, Khurana S. Progress in the development of gelling agents for improved culturability of microorganisms. Front Microbiol 2015;6:698. [CrossRef]
31. Ibrahim IA, Nasr MI, Mohammed BR, El-Zefzafi MM. Nutrient factors affecting in vitro cultivation of Stevia rebaudiana. Sugar Technol 2008;10:248-53. [CrossRef]
32. Sharma S, Shahzad A. High frequency clonal multiplication of Stevia rebaudiana Bertoni, sweetener of the future. J Funct Environ Bot 2011;1:70-6. [CrossRef]
33. Hossain MA, Kabir AH, Jahan TA, Hasan N. Micropropagation of Stevia. Int J Sustain Crop Prod 2008;3:1-9.
34. Aziz RA, Al-Taweel K. Effect of plant growth retadants on Stevia (Stevia rebaudiana Bertoni) acclimatization produced in vitro. Plant Arch 2019;19:1275-84.