
ARTICLE HIGHLIGHTS
Grafting, intercropping, and resistant materials reduce the incidence of iron deficiency.
New chelate molecules or fertilizers stabilize iron in calcisols.
Organic complexes stimulate the soil microbiome by promoting iron bioavailability.
Nanotechnology provides iron in a constant and continuous way in the soil–plant system.
The use of iron nutrition strategies reduces iron deficiency and produces better-quality food.
1. INTRODUCTION
Iron (Fe) is an essential micronutrient for plants that can occur as Fe2+ or Fe3+. Its function is to act as an enzymatic cofactor and participate in cellular respiration and photosynthetic processes, chlorophyll biosynthesis, electron transport, and cell division and expansion. It is also useful in the process of the source-sink translocation of photoassimilates [1,2].
Iron is abundant in the earth’s crust. However, a large proportion of Fe is precipitated in the soil in forms that are not very available to plants, such as insoluble ferric (Fe[III]) hydroxide (Fe[OH]3) – a very common form in calcisols. These soils are of great economic relevance because they cover approximately 25?30% of the world’s agricultural area and are characterized by high pH, calcium carbonate (CaCO3) and bicarbonate (HCO3-) contents, and oxidative redox potentials, and low organic matter (OM) content [3,4]. These conditions cause low Fe availability in the soil and lead to Fe immobilization–precipitation in the apoplast [5,6], resulting in iron deficiency chlorosis (IDC). Iron deficiency chlorosis is characterized by the interveinal chlorosis of young leaves, chlorophyll loss, whitish-yellow discoloration, growth reduction, the deformation of young leaves, yield reduction, senescence, and tissue death [4] [Figure 1]. Likewise, this condition modifies nitric oxide homeostasis, reduces antioxidant system activity, and induces the accumulation of reactive oxygen species (ROS), all of which affect the photosynthetic system [1,7]. In addition, light perception and signaling processes in the coordination of Fe homeostasis alter the morphophysiology of plants, and because calcisols are associated with semiarid and arid areas (with high solar radiation rates), the incidence and level of Fe deficiency can be exacerbated [8].
 | Figure 1: Iron deficiency symptoms in tomato plants grown in calcisol soil (picture by Fabián Pérez-Labrada). [Click here to view] |
The low availability of Fe in calcisols leads to inadequate plant development. To improve crop growth in these soils, several alternatives in the management of Fe nutrition have been implemented and studied, from the use of tolerant plant varieties/hybrids, fertilizers, and chelates to the application of nanotechnology, new molecules, and genetic engineering tools [9,10]. This review represents an effort to show the main characteristics and potential of common strategies for the management of Fe nutrition in calcisols, demonstrate the efficient and sustainable use of Fe under calcisol conditions, and establish a critical perspective from which to approach unresolved or unaddressed aspects that should lead to the development of future research into mitigating IDC and producing better-quality crops.
2. IRON IN CALCISOL SOILS
Iron is abundant in soils (≈30 mg/kg) and can occur as water-soluble Fe, adsorbed Fe, Fe strongly adsorbed on carbonate, metal-organic complex Fe, Fe bound to manganese oxide, organically bound Fe, and residual Fe, although the most abundant form commonly found is Fe oxide, in the form of its main species – Fe(OH)2+, Fe(OH)30, and Fe(OH)4- [11]. However, in the case of calcisols containing more than 15% CaCO3, this compound may be deposited in hard and impermeable layers, or it may accumulate in the pores of soil particles when dissolved in the soil CaCO3, resulting in alkaline conditions due to the increase in HCO3- (CaCO3 + H2O ↔ Ca2+ + HCO3- + OH-), buffering the pH to between 7.5 and 8.5. This condition generates a saturation of Ca2+ and magnesium (Mg2+) bases in the cation exchange sites, leading to Fe adsorption and precipitation reactions [12]. Calcisols may have adequate Fe contents, but this is in a form unavailable to transfer to plants [Figure 2].
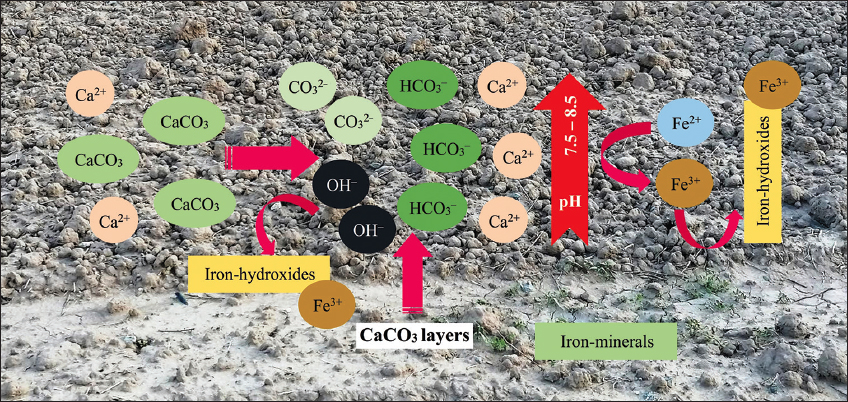 | Figure 2: Behavior of iron in calcisol soils. [Click here to view] |
These soils have a low soluble and exchangeable Fe content, are insoluble, and have difficulties with absorption and translocation [1,13] because the high pH causes the precipitation of Fe in the form of ferric hydroxides [mainly Fe(OH)30 – a chemical form that is not available to plants]. Similarly, soil redox conditions significantly affect Fe dynamics because, as the soil moisture content increases, “reduction microsites” are created in the vicinity of the soil particles – a condition that reduces the redox potential by the release of electrons and protons from the microbiome – causing an increase in Fe solubility. Drought conditions (low soil moisture content) increase the redox potential, generating an environment, in which Fe again precipitates as Fe(OH)30 (insoluble and unavailable to plants) [12,14]. Similarly, waterlogged conditions increase Fe2+ levels due to microbial metabolism, which uses Fe3+ as an electron acceptor [15], and likewise, CaCO3 dissociates into Ca2+ and CO32–, raising the pH of the soil solution, and this environment causes the oxidation of Fe2+ to Fe3+ [12,16,17].
3. ABSORPTION AND TRANSPORT
On average, plants contain up to 50–150 mg/kg dry weight of Fe in their leaf tissue, and a certain percentage is mobilized for seed accumulation during senescence [18]. These concentrations vary according to the carbon (C)-fixation pathway; for example, species with the Hatch-Slack pathway require more Fe than species using the Calvin–Benson–Bassham cycle [19]. To reach these requirements, plants present strategies to access soil Fe, including acidification?reduction?transport (Strategy I, non-grass plants) and Fe3+ chelation by hexadentate chelators and phytosiderophores (Strategy II, grass plants) [Figure 3] [9,20]. Plants under calcisol conditions tend to exploit these strategies to access Fe.
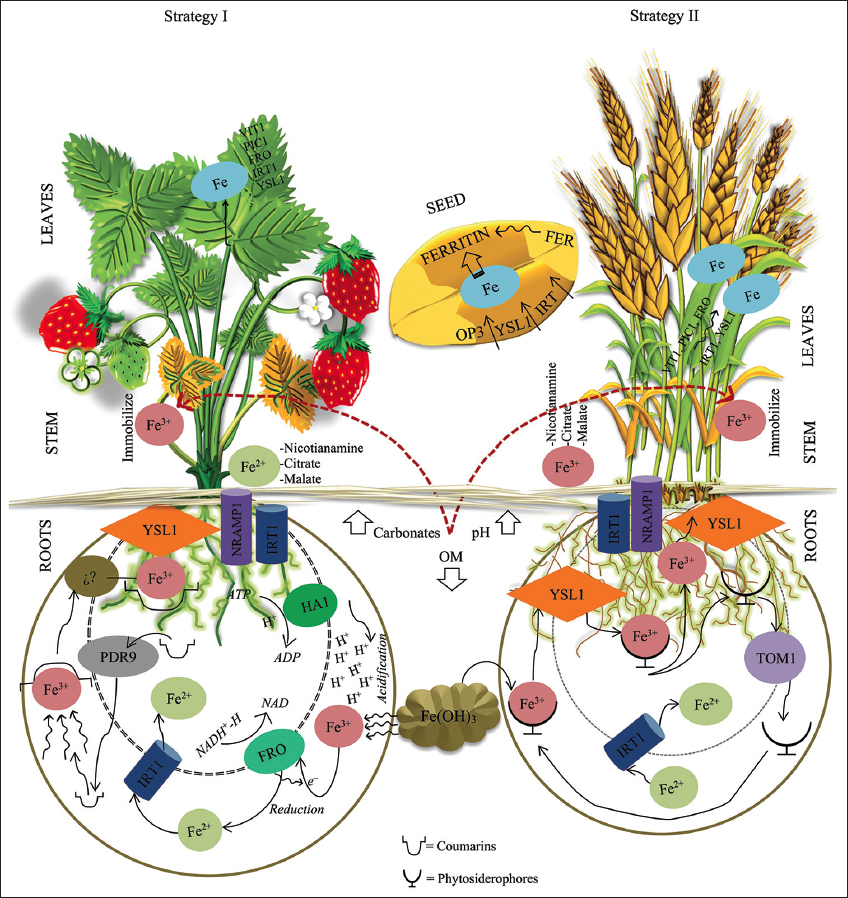 | Figure 3: Mechanisms of iron uptake and transport in plants. [Click here to view] |
In Strategy I, hydrogen (H+) is extruded by the enzyme H+?ATPase (HA1), which acidifies the rhizosphere and allows the solubilization of insoluble Fe3+ complexes. Then, phenolic compounds (mainly coumarins excreted by PLEIOTROPIC DRUG RESISTANCE 9 transporter, PDR9) or organic acids chelate the Fe3+, producing a “zone of optimum concentration” in the rhizosphere. The chelated Fe3+ complex is then reduced to Fe2+ by ferric reduction oxidase (FRO), and finally, the Fe2+ is transported into the plant by the zinc (Zn)-regulated transporter- and iron-regulated transporter (IRT)-related protein (ZIP) family, mainly IRT1, or by natural resistance-associated macrophage protein 1 (NRAMP1). These components are overexpressed in the outer membrane and in the early endosomes of the root system [4,9,18,21-24]. Transcription factors, such as the FER–like iron deficiency-induced transcription factor (FIT), basic helix–loop–helix (bHLH), and myeloblastosis (MYB), promote the activation of this strategy [25].
In Strategy II, the 2’-deoxymugineic acid secreted by the protein transporter of mugineic acids 1 (TOM1) chelates the Fe3+ to obtain a Fe3+–phytosiderophore complex. Subsequently, the transporter yellow stripe-like 1 (YSL1) located in the root epidermis and IRT1/2 capture the complex and mobilize it toward the root cells [4,9,10,21,24].
Excretion of the catecholic metabolite sideretin from the coumarin fraxetin functions as an annex mechanism in Fe uptake by a redox cycle (electron transport) in the root system of plants (probably in a greater proportion in Strategy I than Strategy II plants) [26].
In the root epidermis, Fe is mobilized through the apoplastic pathway (where the Caspary band and endodermis regulate its mobilization) until it is loaded into the xylem (mainly by transporters of the YSL family) or loaded into the phloem (as Fe2+ nicotianamine by YSL transporters) for subsequent translocation and histolocalization through a source-sink mechanism in chelated form – that is, Fe citrate, Fe cysteine, Fe histidine, Fe malate, Fe nicotianamine, Fe aspartate, Fe glutathione, and Fe glutamate – or with IMAI peptides (IRON MAN) [4,21,24,27]. In leaf tissue, it is loaded into the mesophyll cells, probably by YSL and IRT1. In addition, FRO may be activated to reduce the chelated complex. Once inside the cell, it can be redirected and stored mainly in the chloroplast by the protein components complex containing permease in chloroplast 1, a nickel–cobalt transporter, and mitoferrin–like 1; in the vacuole by vacuolar iron transporter (VIT1), VIT-like, and NRAMP; into the mitochondria by FRO and mitoferrins; or be redirected to the seed by oligopeptide transporter 3 (OPT3) and YSL1, where it is stored as FERRITIN in the vacuole by B3 transcription factors, including FUSCA3 (FUS3), leafy cotyledon2, and abscisic acid insensitive 3 [13,28]. Similarly, in the chloroplasts and mitochondria, the enzyme ferrochelatase allows the binding of Fe2+ in protoporphyrin IX, producing heme proteins or Fe?sulfur (S) groups [24,28,29].
With foliar Fe application, the presence of negative charges on the cuticle facilitates the diffusion of Fe (which has a positive charge) or FeO (nanoparticles [NPs]) through the cell wall. There is a certain degree of similarity between the van der Waals radius of chelates and Fe sulfate (FeSO4) and the aqueous pores, such that the Fe can penetrate through the stomata as easily as through the apparent free spaces. The Fe then rapidly penetrates into the vascular bundles, where, depending on the cell pH, it binds to anions and/or ligands (nicotianamine, deoxymugineic acid, and Fe transport proteins) or reacts with nitric oxide, forming nitrosyl, and dinitrosyl complexes (mainly under Fe-deprived conditions) for subsequent cell compartmentalization, as mentioned above [20].
The characteristics of calcisol soils (mainly their high pH and high HCO3– contents) and their Fe-deprived conditions significantly affect the plant physiology because the reduction in tissue Fe content generates a decrease in chlorophyll synthesis and alters the carbonic anhydrase, ribulose-1,5-bisphosphate carboxylase activase and phosphoenolpyruvate carboxylase (PEPC) activity, gas exchange (net photosynthesis), and chlorophyll fluorescence [30-34], and these alterations impair plant growth rate and production.
Iron deficiency in plants grown in calcisol soil leads to oxidative stress through an increase in the synthesis of ROS (H2O2, O2•, HO) and malondialdehyde [33]. This oxidative environment promotes proline concentration while reducing the synthesis of catalase, superoxide dismutase, and peroxidase because Fe is a constituent of these enzymes [30]. This enzymatic response varies according to the genetic resource and strategy used in Fe management.
The physiological and biochemical response observed in plants with IDC varies according to the level and timing of the Fe deprivation, both in Strategies I and II. In both cases, there is a molecular response that allows the mechanisms of Fe acquisition to be regulated. In this sense, transcription factors allow signaling and modulation. In plants developed in calcisol soils, the transcription factors FER, FIT, bHLH, and MYB, and the transcript expression of NRAMP3 and yellow stripe 1 (YS1)/YSL, are crucial for detecting Fe deprivation in the plant, thereby allowing this promotion and modulation of the signaling [25]. These transcription factors regulate the expression of the gene AHAs (encoding the ATPase protein in the plasma membrane), FRO (encoding the FRO protein), IRI1 (encoding the IRT protein), PEPC (synthesizing the PEPC1), YS1/YSL (encoding the Fe(III)-phytosiderophore complex transporter), and TOM1 (synthesizing the phytosiderophore transporters) [35-38]. These authors cited differential modifications in the expression of the genes in tomato (LeIRT1, LeFRO1, LePEPC1), rice (OsIRT1), barley (HvYS1), and maize (ZmYSI), as well as their heterologous nature in different plant organs.
4. MANAGEMENT STRATEGIES
To mitigate the incidence and levels of IDC in crops and to avoid inadequate plant development, different alternatives for its management have been investigated and implemented that can contribute to sustainable crop production in calcisols [9,10].
4.1. Grafting
Grafting is a practice based on the union of a plant of particular interest (graft) on another plant (rootstock) that presents resistance characteristics and is compatible with grafting. This technique has been reported in grapevine cultivars developed in calcisols to avoid IDC (given that grafting + quality rootstock → Fe accessibility and uptake and thus ↓ IDC) by promoting Fe uptake strategies through the modification of 3-hydroxymugineic acid, 3-desoxymugineic acid, and stearyl ferulate exudation patterns [39]. For example, in a pear cultivar (Pyrus bretschneideri cv. Rehd) grafted onto a dwarf quince (Cydonia oblonga cv. Molino) grown in a calcisol, a high susceptibility to IDC was found, whereas a P. bretschneideri cv. Rehd grafted onto a Pyrus betulifolia showed a mechanism of Fe activation in the leaves [40].
The positive response to the use of grafting in the management of Fe nutrition may be due to an alteration in the FRO activity or by exudation of low-molecular-weight compounds from the root system of the rootstock. However, in calcisols, the high bicarbonate concentration may lead to alkalinization of the pH in the xylem, causing inefficiency in the Fe transport or precipitation. This condition increases the concentration of inactive Fe in the apoplast of Fe-deficient leaves compared to non-deficient leaves (Fe-chlorosis paradox). In this context, using rootstocks can minimize the increase in apoplastic pH, stabilize the FRO activity, and mitigate the Fe precipitation in the leaf tissue. An appropriate selection of the rootstock–cultivar combination is necessary to initially facilitate the greatest survival and enhance the characteristics of the plant species; for example, the use of various Prunus rootstocks has led to agronomic alterations and modifications in the quality of the graft fruit in addition to a differential response in IDC and the foliar Fe content [41-44]. The graft’s physiological response may have resulted from histological and genetic interactions with the rootstock [45]. The graft-rootstock behavior, Fe content, yield, and incidence of chlorosis depend on the level of CaCO3 in the soil [46].
4.2. Intercropping Systems
Intercropping systems (a method that consists of sowing/developing more than one crop in a given area at the same time) have positive effects on the Fe nutrition derived from the interrelationships occurring in the root system of the plants. In an intercrop of peanut (Arachis hypogea L. cv. Luhua14) and maize (Zea mays L. cv. 4 Nongda108), the Fe content was improved by facilitating the root acquisition of Fe through modifications in the Fe transporter (AhNRAMP1) and divalent metal ion transporter [47]. Those authors pointed to an increase in methionine synthesis (derived from increased homocysteine S-methyltransferase and serine acetyltransferase 1) possibly related to an increased release of phytosiderophores. In another study, the intercropping of peanut (Strategy I) and maize (Strategy II) led to reductions in IDC and an increase in Fe in the peanut. This response was due to the AhYSL1 gene expressed in the root system of the peanut allowing the uptake of the Fe(III) phytosiderophore complex located in the rhizosphere of the maize [48]. Similarly, planting wheat, alfalfa, and maize in calcareous soil, in combination with chemical fertilization or manure, modified and increased the concentration of available Fe (mainly in the alfalfa) and promoted higher concentrations of Fe associated with CaCO3 and OM and lower contents of Fe associated with soil minerals [49]. Contrastingly, a peanut–corn intercropping system, developed in calcareous soil (pH 7.92), showed strong interspecific interactions that promoted both N and Fe acquisition, which may be because the corn root system under a Fe deficit increased the release of Fe phytosiderophore complexes in the rhizosphere zone. This availability allowed the peanut to absorb Fe, increasing the activity of FRO and specific transporters [50].
Intercropping systems positively promote the physicochemical and biochemical characteristics of the soil, rhizosphere, and microbiome [51], improving Fe absorption processes in calcisols, with crop1 + crop2 + cropn stimulated plant?plant relationships leading to stability of the plant-soil system, and thus to Fe bioavailability. However, for this strategy to be more efficient, it is necessary that the plants used present only slight complementation to avoid competition (e.g., combining a species with a high solar radiation demand with a species that demands less), that the water requirements are adequate, and, above all, that the allelopathy is analyzed, with the objective of enhancing the combination of crops beyond leguminous–non-leguminous pairs.
4.3. Fertilizers
The use of fertilizers is common practice in relation to ferric nutrition due to their rapid response. However, the qualities of the fertilizer must be verified to avoid its precipitation when it comes into contact with soil particles. Likewise, its relationship with S must be accounted for, and excessive applications of copper (Cu) and Mn must be avoided because Fe absorption can decrease if there is an accumulation in the roots [52,53].
Due to its solubility and acceptable cost, the most widely used inorganic fertilizer for supplying Fe is FeSO4. Several studies have demonstrated the benefit of its use, with applications of 12 kg/ha on flour wheat in soils with pH 7.5?7.7 improving the Fe and starch content in the grain, while a combination of 12 kg/ha FeSO4 and 10 kg/ha ZnSO4 increased productivity and the Zn, Fe, Ca, and starch content [2]. Similarly, the use of foliar-applied FeSO4 has been effective in reducing IDC in the leaves of orange trees (Citrus sinensis L. cv. Thomson Navel, grafted on Citrus limon L.) grown in calcareous soil (pH 7.9) due to the remobilization of inactivated Fe in the apoplast, thus avoiding its oxidation and precipitation [54]. Paspalum scrobiculatum cv. CO3 plants developed under calcisols did not show IDC when 50 kg/ha of FeSO4 was applied to the soil in addition to foliar spraying at 0.5% at 30 and 50 days after planting [55]. Another example is the combined use of FeSO4 and biochar, which promoted root biomass and photosynthesis by improving the Fe and ferritin content and possibly reducing the soil pH [56]. Similarly, applications of S and bentonite granules to calcisol soil (inoculated with Thiobacillus thioparus) reduced the soil pH (by ≈–0.3 units) by Thiobacillus-mediated S oxidation, thereby improving the growth parameters and increasing the Fe uptake, in addition to the Mn, Cu, S, and phosphorus (P) in corn plants [57].
In addition to ferrous sulfate, using nitrogen fertilizers (ammonium [NH4+] sulfate, NH4+ nitrate, or urea) allows localized soil acidification, promoting greater Fe availability [58]. However, it should be taken into account that the uptake of N?NO3- is associated with H+ cotransport systems or OH- antiporters, so the rhizospheric pH could increase, reducing the Fe availability, and the excreted H+ could be neutralized by HCO3- [18]. When N?NH4+ encounters the soil, it is deprotonated (loses a H+), inducing a localized reduction in pH; however, the resulting ammonia (NH3) can easily volatilize. In this situation, a N?NO3-/N?NH4+ balance is suggested to allow the proper activity of nitrate transporter 1, which makes the reuse of apoplastic Fe more efficient [59].
The combined application of mineral fertilizers in saline-alkaline soils can be a management strategy for stimulating soil aggregation, the content of amorphous Fe oxides, and the distribution of C-aromatic, the latter being capable of forming Fe-aromatic complexes [60]. Another alternative is the application of ceramic frits (of fused, tempered, and granulated ceramic composition) that can increase the available Fe content in the soil (pH 8.25). Using these frits on kiwifruit, melon, and zucchini plants increased the foliar Fe content in the melon plants [61]. According to these authors, Fe release increased under alkaline conditions, suggesting its potential use as a slow-release inorganic fertilizer in calcisol soils. Recently, fertilizers coated with organic acids and amino acids (citric, humic, fulvic, and salicylic acids, and glycine) have been developed to allow a controlled release of nutrients, with granules coated with citric acid (10% w/v), especially allowing a greater availability of S and micronutrients, such as Fe, in calcisols [62].
To achieve sustainable Fe administration in calcisols, the 4Rs perspective is recommended. The 4Rs nutrient stewardship framework is based on four interrelated principles – the right source, right rate, right time, and right place [63]. This involves selecting the most adequate source for providing the Fe (preferably acid-reaction fertilizer/s), verifying the ideal dose based on the plant developmental stage, establishing the adequate moment(s) for application, and determining whether drench, edaphic, or foliar application should be used. This method can provide greater certainty in reducing IDC in the plants developed in this type of soil. Similarly, using biodegradable materials (palm and pistachio cellulose) to coat slow-release Fe fertilizers is effective and plausible [64].
4.4. Chelates and Novel Chelates
The application of conventional synthetic chelates, such as ethylenediaminetetraacetic acid (EDTA), ethylenediamine-N-N’bis(o-hydroxyphenyl acetic) acid (EDDHA) in different racemic ratios, and N-N’bis(hydroxyphenyl) ethylenediamine-N-N-N’-diacetic acid (HBED), is a common and effective agricultural practice used to prevent and/or mitigate IDC [21,65]. However, different application times are required for its efficiency, and ideally, before the first symptoms of IDC have manifested [18]. Applying HBED/Fe3+ chelate in the early stages of plant development is the most appropriate time for achieving a greater accumulation of Fe in the plant and in the soil [66]. In A. hypogea L., the foliar application of Fe-EDTA (0.1 mM) at 45, 60, and 90 days after sowing improved the pod number in plants developed in calcisols due to a possible reduction in the apoplastic pH that allowed Fe stabilization, mobilization, and metabolism [5]. Alternatively, applying Fe chelates to date palms grown in calcisols improved their fruit yield [67]. Similarly, the foliar application of Fe-EDDHA to tomato plants (in a calcisol with pH 7.9) improved their growth and yield [68], while applications of 4.8% of the same chelate to the tomato plants led to increased efficiency in Fe uptake. Thus, this may be a useful way of mitigating IDC, while a dose of 6.0% may serve as a long-term nutrition strategy [69]. To achieve efficiency in the use of chelates, the pH and carbonate content (buffering capacity) of the soil must be considered; otherwise, only short-term results will be obtained. Some chelates, such as EDDHA, lose stability due to the retention of their isomers in the soil particles [18].
Another alternative is the combined application of chelates and fertilizers, for example, applications of Ca nitrate (Ca[NO3]2] (50 g/tree) and Fe chelate (6%, 30 mL/tree) in Malus domestica Borkh cv. Red Delicious grown in calcisols improved the yield and fruit quality by optimizing the postharvest quality [70]. In addition, ferric fertilizer sprays (particularly citric acid + Fe2[SO4]3 + Fe-EDTA-Na) can increase the yield and Fe content in potato tubers grown on alkaline soil (pH 8.3?8.4), improving the photosynthetic pigments and leaf gas exchange [71]. In addition, in Italian lemon, the direct soil application of Fe-EDDHA + urea (1 month after the onset of sprouting) was found to increase the Fe content in the leaf tissue and the yield [72]. Although the use of these chelates increases the Fe content, reduces the IDC, and improves the fruit quality, their efficiency may be reduced by precipitation in the soil, and the low translocation or accumulation in the cuticle through foliar application [19].
Considering that conventional chelates can be highly persistent in the environment, alternative molecules have been sought to chelate the Fe. In this sense, N,N-dihydroxy-N,N’-diisopropyl hexanediamide (DPH) and azotochelin have been applied to soybean to investigate their stability in improving Fe nutrition as a long-term source of Fe, and inducing dissolution in calcareous soil [73]. It has also been reported that ethylenediamine disuccinic acid ([S,S]-EDDS/Fe) chelate applied to calcisol increased the Fe content in soybean plants similarly to the application of a Fe-EDTA chelate [74]. One of the qualities of the EDDS chelate is its low durability in soil, although this causes it to have a lower reactivity with the Ca in CaCO3, such that Fe is made available. In another study, it was found that the use of Fe chelate tris (3-hydroxy-1-(H)-2-methyl-4-pyridinonate) Fe(III), applied to soybean grown in calcisols on two occasions (the V3 and V4 phenological stages) at a dose of 5.5 mM, improved the chlorophyll content and root biomass in addition to the Fe content in the trifoliate leaves by promoting a system of absorption, redistribution, and accumulation of Fe in the plant, correcting the IDC [75].
In other studies, knowledge of the functionality of 2’-deoxymugineic acid (Strategy II) has been used to synthesize a more stable and economically accessible analog. In this sense, the compound proline-2’-deoxymugineic acid + Fe can be mobilized toward the plant as a Fe chelate by Fe(III)-2’-deoxymugineic acid transporters, reducing the Fe chlorosis in rice plants, and even applications of proline-2’-deoxymugineic acid chelate Fe found in calcisols enhanced its effect by up to 10-fold compared to Fe-EDDHA [76]. This molecule has also been tested on Strategy I plants (e.g., cucumber) grown in calcareous substrates (pH 9.1), where a reduction in IDC was observed along with an increase in Fe in the shoots due to its high reducibility [77]. Similarly, its application as proline-2’-deoxymugineic acid in peanuts grown in this type of soil can dissolve the insoluble Fe in the rhizosphere and stimulate AhYSL1 gene expression, thus improving the Fe nutrition, yield, and nutrient content in the grain [78]. Finally, the application of deferoxamine combined with Fe3+ to the roots of Cinnamomum camphora grown in saline–calcisol soil (pH 8. 2) alleviated the IDC by increasing the leaf Fe content by stimulating its uptake and transport system (CcbHLH, CcFRO, CcNramp, CcOPT, CcIRT, and CcVIT), as well as generating a higher solubility and mobility of the Fe in the plant due to the reduced activity of pectinesterase, which fixes Fe in cell walls [79].
Conventional chelates can cause contamination due to their high persistence in the soil and water fields, so their use in the context of the 4Rs is recommended, for example, selecting the right chelate source, right dose, right time, and right place. It is also to reduce and/or mitigate interesting to promote the use of novel chelates under a similar approach in both cases to reduce and/or mitigate plant IDC sustainably.
4.5. OM
Given that soil OM plays a crucial role in increasing Fe availability, the addition of organic sources (such as humic complexes) can stimulate soil microbial activity and/or induce the reduction of microsites – phenomena that can lead to bioavailable and stable Fe concentrations for prolonged periods in the soil and in the soil solution. A widely studied source of OM in soils is leonardite. In this context, applications of Fe fulvate (fulvic acid + Fe2SO4) in serrano chili grown in calcareous soil improved the growth and fruit quality in addition to inducing conditions for an optimal Fe content [80]. In the same context, humic substances adsorbed on weakly crystalline Fe oxides (ferrihydrite) can stabilize Fe in the soil, favoring its uptake. However, their effect varies depending on the type of strategy used by the plant. In wheat (Strategy II), an increase in Fe uptake was observed to result from the mobilization of Fe from the oxides in comparison to white lupin (Strategy I) [81]. In Glycine max, the application of a 2:1 mixture of Fe leonardite humates plus a chelating agent (Fe-HBED) provided available Fe to the chelating agent and subsequently to the plants, inducing a synergistic effect. Unlike the chelates, the Fe leonardite mixture maintained a stable humate supply. The same authors [81] noted an increase in the shuttle effect from a synergistic application of these compounds derived from the solubilization of the Fe3+ present in the soil and provided by the leonardite, as well as its subsequent chelation [82]. This was because humic substances can stabilize, solubilize, and provide plant-available ferric complexes [83].
Alternatively, weekly applications of humic complexes to calcareous soil (pH 8.5, 0.2% OM, 5 mg/kg Fe), in combination with a Fe-EDTA or Fe-EDDHA chelate, improved the Fe content in tomato plants [84]. In a study conducted over the course of 3 years (2017–2019) on Vitis vinifera L. cv. Tempranillo grafted on 41?B rootstocks and developed in calcareous soil (pH 8.265, 1.195% OM), applications of leonardite supplemented with ferrous sulfate heptahydrate increased the soil OM content, favoring root growth, slightly reducing the soil pH (≈ –0. 2 units), and improving the nutrient content of the soil [P, potassium (K), Fe, Mn, Cu, and Zn] and petiole (P, K, Fe, and Zn) [85]. The authors pointed out that the reduction in soil pH was because the Fe2+ in the sulfate was very labile in soils with good aeration and high pH because it reacted with oxygen (O2) to generate Fe3+, which hydrolyzed to form Fe(OH)3, releasing three H+ into the soil for each Fe3+. The FeSO4?7H2O produced H2SO4, Fe(OH)3 formed Fe2O3?nH2O, and H2SO4 was neutralized by CaCO3, thereby releasing Ca2+, SO24–, and HCO3-, the latter of which was able to dissociate into CO23– and H+. Finally, the foliar spraying of humic acid plus Fe on rice, soybean, and lettuce crops in soils with a pH of 8.44 resulted in increased growth, higher yields, better Fe accumulation in the aerial parts, and increased Fe mobilization to the seeds (rice and soybean) [86].
Using animal manure amendments (at an application rate of 10%) on pearl millet improved the Fe availability in calcareous soil (pH 8.5) and stimulated its uptake and accumulation in the plant. A reduction in IDC symptoms by stimulating photosynthesis and plant growth has also been reported [34].
4.6. Amino Acids and Low-Molecular-Weight Compounds
Amino and organic acids are molecules that allow metabolic activation in plants; their exogenous application can be a tool in managing Fe nutrition. Foliar applications of sulfate plus organic acids to Platanus orientalis (0.7% FeSO4) showed a better response than soil applications because they were able to influence the root system and rhizosphere of the trees [87]. In the same way, foliar applications of amino chelates (2.0% N-amino acids, 2.5% Zn, 2.0% Fe, 1.5% Mn, and 0.4% Cu) to Solanum lycopersicum L., Cucumis sativus L., and Phaseolus vulgaris L., grown on calcareous soils, improved the chlorophyll index and yield [88]. Contrastingly, applications of organic acids (1 mmol citrate, malate, and oxalate) and flavonoids can mobilize Fe in calcareous soils, inducing the dissolution of amorphous fractions, and allowing illite formation in C. sativus grown in calcareous soils (pH 8.0) [89].
To increase the biodegradation rate (concerning synthetic chelates), new complexes have been reported, such as the compound Fe3+ heptagluconate (in equimolar and 1:2 ratios), which gave greater stability to the Fe in the soil at high pH values as a result of the formation of polynuclear complexes [90]. Alternatively, FeSO4?7H2O, Fe glycine, Fe tyrosine, and Fe chitosan complexes have been developed. These were applied to the soil (20 mg Fe/kg) or leaves (0.25% w/v) of wheat in soil at pH 7.8 and were found to have improved the yield in addition to promoting Fe and N uptake and increasing the Fe concentration in the shoots. This response was derived from an improvement in the transport systems [91]. Applying these compounds implies lower energy costs for Strategy II plants, in addition to the fact that amino acids can participate in protein synthesis (e.g., Fe transport). Likewise, ferrous glycinate and ferrous citrate increased the Fe content through foliar application. Ferrous glycinate, in particular, induced greater chlorophyll biosynthesis and improved photosynthesis in Vigna [92]. Applying amino acids to soil-developed tomato (pH 7.17, available Fe 1.48 mg/kg) under greenhouse conditions promoted better yields, fruit weights and diameters, and higher concentrations of N, P, Fe, and Mn [93].
The application of various Fe sources to soybean grown in a calcisol can be considered a sustainable alternative in the management of Fe deficiency. Mining byproducts (Fe oxide and Fe-FeS2 oxide), in combination with thiols (glutathione, dithiothreitol, and thiophenol), increased the Fe in the plant and modified the content of reduced-oxidized glutathione, ascorbate, and antioxidant enzymes [94]. Mining byproducts are more effective than Fe-EDDHA chelate, probably due to their ability to release Fe, which can be solubilized and absorbed by the plant acquisition mechanism. Similarly, in tomato grown on calcisols, the foliar spraying of Fe glutamate improved the plant height and shoot and root fresh weights and increased the fruit number, yield, and quality, indicating that Fe glutamate can improve Fe nutrition by acting as a biostimulant [95].
Drench applications of nutrient solutions containing organic acids (citric or salicylic acid) and chelates (Fe-EDTA or Fe-EDDHA) to pot-grown tomato in calcisol soil (pH 8.5, 0.2% OM, and 5 mg/kg Fe) promoted Fe content in the shoot tissue. It increased the yield per plant [85,96]. Finally, applications of ascorbic and oxalic acid to cucumber plants grown in calcisol soil (pH 8, 10% CaCO3) appeared to reduce abiotic stress by histologically modifying the xylem duct and cortical cells [97].
4.7. Microorganisms
Plants tend to interact strongly with the rhizospheric microbiome to aid in their Fe uptake strategy. Likewise, some microorganisms under Fe deprivation can produce siderophores that chelate Fe3+ and induce its bioavailability. In this sense, the selection, propagation, and application of such microorganisms is a strategy that can contribute to Fe uptake in calcisols [98]. It has been shown that Azotobacter vinelandii and Bacillus subtilis, in combination with FeCl3?6H2O, can be a viable way to reduce IDC in soybean plants grown in calcisols [75,99]. The simultaneous application of growth-promoting rhizobacteria (Pseudomonas fluorescens T17?24 and B. subtilis P96) and Fe chelate (10 mg Fe-EDTA/kg soil) to sorghum grown in calcisol showed improvements in the morphological characteristics of the crop due to the dissolution of insoluble compounds, which increased the availability of Fe in the rhizosphere and expanding the Fe, Mn, Zn, Cu, and K contents in the plant [65].
Alternatively, microorganisms such as Pantoea agglomerans, Pseudomonas plecoglossicida, and Lactococcus lactis can synthesize pyoverdine siderophores (ferripyoverdines), catecholate, hydroxamate, and carboxylate, so that their application, in addition to modifying the plant ionome, can chelate Fe, facilitating its absorption, and increasing its content in the plant [100,101]. Similarly, soil applications of inocula of Glomus spp. microorganisms, Trichoderma asperellum, and Streptomyces avermitilis on strawberry plants (Strategy I) in calcisol were found to reduce IDC symptoms by increasing the Fe content in the different tissues. This was explained by an increase in Fe availability by the siderophores of the microorganisms [102]. In a 1:1 mixture of vermiculite and calcisol (pH 7.8), the individual or combined inoculation of Sphingobium fuliginis ZR 1?6 and Pseudomonas jessenii ZR 3?8 in soybean plants increased the FRO activity, allowing Fe3+ reduction, and improving Fe uptake and concentration in the tissues. This action was due to the production of indole-2-acetic acid, 1-aminocyclopropane-1-carboxylic acid deaminase, siderophores, and organic acids [103].
Rahnella aquatilis JZ-GX1 applied to C. camphora induced a substantial improvement in the chlorophyll and Fe2+ content in the leaves, in addition to increasing the activity of the superoxide dismutase, peroxidase, catalase, and ascorbate peroxidase enzymes [104]. The same authors pointed out the probability that this strain could secrete desferrioxamine in the rhizosphere and promote increased abundances of Glomus, Mortierella, Trichoderma, and Penicillium in calcisols at pH 8.2. Recently, it was found that the coinoculation of Piriformospora indica (phytopromotional fungus) and Azotobacter chroococcum WR5 (rhizobacterium) in Triticum aestivum L. altered the root system and enhanced Fe accumulation in plant tissues by increasing the root, stem, leaf, and grain transfer factors and the Fe transfer factors in the root?stem, root?leaf, and root?grain systems, as well as increasing the overexpression of ZIP-family transporter genes in the root system [105]. Similarly, Thiobacillus thiooxidans and Thiobacillus ferrooxidans in calcisols can increase the Fe concentration in soybean shoots [106].
The joint application of Bacillus megaterium and Ca superphosphate (69, 207, and 345 kg/ha) led to a reduction in the damage caused by a high soil carbonate content (10.8?11.3%, pH 7.19?7.77), and improved the content of Fe, Cu, Zn, and Ca in the leaf and root tissues [107]. Similarly, biochar applied with plant growth-promoting rhizobacteria and mycorrhizal fungi under wheat seed at sowing under rhizobox conditions improved the availability of Fe, Zn, and P in the soil, thereby increasing the OM [108]. The increase in Fe bioavailability was due to the synthesis of microbial siderophores that could promote Fe solubility and stability, with the biochar functioning as a niche for microbial proliferation due to its high porosity and through the interaction of different Fe fractions with the biochar and soil [109]. Microbiome–soil?plant?root interactions under Fe-deficient conditions may depend on coumarin excretion [110].
4.8. Bioengineering: Plant Breeding
Selective crop management allows desirable and interesting traits to be obtained (conventional breeding). However, genetic engineering (gene manipulation) allows a more efficient and rapid response. In both cases, the objective is to understand and improve the mechanisms of Fe uptake, translocation, and utilization at the transcriptomic level in IDC-tolerant/resistant genotypes [18,111,112]. For example, the introduction of barley genome fragments containing the OsIRT1?refre1/372 promoter (increased chelate reductase capacity) and the 35S?OsIRO2 promoter (increased phytosiderophore secretion) into an Fe deficit–tolerant rice crop allowed enhancement of the gene expression related to phytosiderophore synthesis and ferric chelate reductase activity, enabling the efficient acquisition of the Fe present in a calcisol (pH 8.9), which allowed a 9-fold increase in yield compared to non-transgenic rice plants [113].
Some plant species or cultivars show a higher tolerance to Fe deficiencies. For example, Medicago truncatula line TN8.20 showed better performance when grown in a calcisol than the Jemalong line, which is possibly due to the differential capacity for Fe allocation to nodules, thereby enabling their symbiotic N-fixing functionality and the higher enzymatic activity of superoxide dismutase, catalase, and peroxidase [16]. Likewise, a higher relative tolerance to Fe deprivation conditions was found in the calcisols of durum wheat cultivars due to a higher Fe uptake and translocation capacity together with its efficient use [112]. In this regard, genetic engineering strategies have been proposed to exacerbate the synthesis of certain genes as well as their promoters (YS1 controlled by the heavy metal ATPase 2 promoter, transporter of mugeneic acid 1 controlled by ferric reductase defective like 1 promoter, ferritin, and nicotianamine synthase), which may correspond to an increased Fe content in maize and rice seeds [114]. The combined introduction knockdown of the OsHRZ and OsIRT1?Rfre1/372 promoter (artificial gene) can lead to improved tolerance of Fe deficiency without reducing its accumulation. This approach has allowed plants to exhibit increased reductase activity at elevated pH levels (in calcisols), leading to Fe accumulation in rice seed and rice straw. Thus, the augmentation of transcription factors is a feasible tool [115].
Further potential for the genomic improvement of plants could be the genetic mapping of IDC-tolerant plants [116] or wild plants. The latter plants (Strategy I) developed in calcisols have been reported as presenting a different absorption and assimilation mechanism because they did not show a strong effect at high carbonate concentrations and soil pH values. This suggests the exudation of specific chemical reductants [117]. Consequently, genetic mapping techniques, and even clustered regularly interspaced short interspaced palindromic repeats (Cas9) [118], could present a way to improve the metabolic pathways that comprise such chemical reductants or exacerbating genes related to their uptake, transport (FRO and IRT1), and storage (Ferritin).
4.9. Nanotechnology
Nanotechnology (a tool that uses compounds <100 nm in size) allows environmentally friendly applications because very low concentrations are required to deliver the nutrient, which, due to its specific characteristics, will have completely different morphological and physiological effects than a particle at its normal scale [119]. Nanoscale Fe (nanoiron) shows rapid penetration, its release, and distribution being controlled and specific. When applied foliarly, it enters the plant through the stomata, hydathodes, and apparent free spaces, whereas, when applied to the roots, it enters through the youngest root cells. In both cases, it is mobilized, through apoplastic and symplastic pathways, to the vascular bundles for translocation by Fe transporters or by phagocytosis, pinocytosis, or endocytosis, can accumulate in the cell. This accumulation results in the supply of Fe for the required metabolic processes and stimulation of the plant antioxidant system [120,121].
Applications of Fe nanofertilizers synthesized from leonardite and containing ferrihydrite in their structure have produced a slow and continuous supply of Fe to soybean plants under calcisol conditions (pH 7.9) [122], while the application of nanoiron (15 mg Fe/kg) to soybean grown in slightly alkaline soil (pH 7.81) induced an increase in shoot-and-root fresh and dry weights, as well as higher leaf numbers, due to a high nutrient-use efficiency with respect to FeSO4?7H2O and Fe-EDDHA [123]. Similarly, under calcisols, cumin (Cuminum cyminum L.) plants responded positively to foliar applications (before flowering) of 1 g/L nanochelated Fe, which improved the yield and plant height [124]. Contrastingly, the application of 10% diammonium phosphate, nanocoated with KFeO2 and urea and 25% K2SO4, to wheat under calcisol-alkaline soil conditions appears to be promising, having improved the growth and development and increased the Fe uptake in plants [125]. These authors pointed out that the positive responses may have resulted from the nanocoating causing a slow release of nutrients. In another case, foliar nutrition with Zn oxide (ZnO) NPs (1000 mg/L) and ascorbic acid (250 mg/L) in sweet potato plants (Beauregard) grown in calcisols (pH 7.19?7.77) promoted higher Fe, Mn, P, and Cu contents and improved tuber yields, perhaps by stimulating the root area, phytohormone synthesis, and ROS scavenging, possibly triggered by the high carbonate content (10.8–11.3%) in the soil [126]. The application of green Fe NPs to the soil increases Fe release and thus its uptake, as well as the growth and photosynthetic pigment content of sorghum plants. This behavior is due to Fe polyphenol complexes (from green NPs) and the reduction of Fe3+ to Fe2+, causing high Fe availability in the soil while reducing the precipitation of Fe oxides-hydroxides in the roots [127].
Using nanotechnology in conjunction with humic substances may be a promising way of reducing Fe-deficiency problems in calcisols and alkaline-reactive soils, in addition to presenting low to no toxicity levels in the environment and maximizing the physiological changes caused by humic substances [83]. In this sense, the foliar application of Fe oxide NPs supplemented with humic acid (Fe?NPs?HA) to geranium plants grown in alkaline soils (pH 7.66) improved their yield and stimulated metabolite accumulation as a result of a higher Fe content, which caused an increase in cellular energy and activation of the enzyme nitrate reductase. This prevents alterations caused by Fe deficiency to a greater extent than sprays of EDDHA, ferrous sulfate, and even Fe NPs [31]. Similarly, foliar applications of fulvic substances in combination with Fe in the form of nanoferrihydrite to spinach improved the total leaf Fe and chlorophyll contents, and the plant fresh weight [32]. This combination allowed for a faster penetration into the leaf. In plant tissues, these substances can induce morphological changes and affect the activity related to nutrient uptake by the plasma membrane [83], which is maximized by Fe at the nanometer scale [31]. The use of zero-valent nanoiron could be an alternative strategy for managing Fe nutrition in these soils, as opposed to FeSO4 and Fe-EDDHA applications, for improving grain Fe concentrations [128].
The foliar application of g-Fe2O3 nanomaterials to soybean has been presented as a sustainable and highly efficient strategy for mitigating Fe deficiency; this nanomaterial induces significantly greater growth than Fe-EDTA by increasing the biomass, nodules, N fixation, yield, and free amino acid content in the seed [129]. These positive effects may be due to an increase in energy supply, reduction in ROS, and phytohormonal modulation. However, it will be necessary to apply this technology at the field level in calcisols soils. It should be noted that through the reduction of Fe NPs through the Maillard reaction, efficient, and environmentally friendly systems that are highly tolerant to alkaline environments have been developed [130].
Combinations of these strategies have been reported as being suitable tools for improving plant Fe nutrition by promoting Fe availability in calcisols [131-139] [Table 1]. Plant chlorosis can be alleviated by organic management practices, coupled with proper water management. This method reduces the soil pH, which affects the availability, mobilization, and uptake mechanisms of Fe [132-136]. It is possible that these practices can supply Fe to plants in the form of natural chelates, slowly and continuously [136], generating an increase in active Fe in the plant tissues. This increase, mainly in the apoplast, alters the source-sink distribution rate, alleviating IDC [137].
Table 1: Summary of research on the combination of strategies for ferric nutrition management in calcareous soils.
| Strategies | Findings | References |
|---|---|---|
| Biochar impregnated with Fe. | Increases Fe availability, Fe content, and fresh and dry weight of the soybean plant. | [131] |
| Foliar spraying of Fe + no–tillage system or flattening soil without the use of mulch. | Promotes the FRO activity of the root system, alleviating chlorosis on bread wheat plant. | [132] |
| Manure applications plus aeration system. | Improve soil Fe and its use in the leaves of pears plants. | [133] |
| Irrigation with carbonated water. | Improves the morphological development of the plant and nutraceutical quality of the grape berry. | [134] |
| Foliar and basal application of FeSO4 together with citric acid sprays and applications of enriched organic compounds | Yield promotion, improved availability of Fe, in groundnut. | [135] |
| NPK in combination with Fe (as enriched farmyard manure) and Bacillus. | Improve peanuts yield. | [136] |
| Trunk injection and FeSO4. | Increased chlorophyll and active iron content in leaves, improving weight, volume, and yield on peach trees. | [137] |
| Biochar with foliar chelate and nano chelate. | Induces positive response of vegetative attributes and photosynthetic pigments of spikenard plants. | [138] |
| Inoculation (bacterial strains) and grafting. | Increased plant growth, leaf area, and iron of peach plants. | [139] |
However, because agroecosystems are network-responsive systems [140], it is necessary to verify the pH and the Fe, carbonate, and OM contents of calcisols, in addition to determining the crop(s) of interest for an adequate selection of the most relevant strategies for enhancing interactions for sustainable Fe nutrition management.
5. CONCLUSION
In this review, we have summarized the present knowledge concerning calcisol conditions, including management strategies for minimizing the prevalence and/or incidence of IDC in crops. These included grafting, intercropping, the use of fertilizers and chelates (both conventional and novel), organic molecules, engineering, and nanomaterials. Each strategy has its complexities, but each offers the sustainable management of Fe nutrition in these types of soils.
The presented studies demonstrated how modifications are carried out at both the structural and functional levels in the soil?plant system, in terms of promoting Fe nutrition, increasing its bioavailability, stimulating microbiome activity, improving enzymatic activity, and stabilizing Fe for adequate absorption, transport, and assimilation.
Signaling molecules, such as hydrogen sulfide and nitric oxide, have been studied as alternatives to Fe nutrition, but need to be tested under calcisol conditions. In this context, we recommend implementing one or more strategies to improve plant growth and development in calcisols by reducing IDC, generating better-quality food, and improving soil sustainability.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to the conception, design, acquisition of data, analysis, and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. FUNDING
There is no founding to this report.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All data generated and analyzed are included within this research article.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Ahmadi H, Motesharezadeh B, Dadrasnia A. Iron chlorosis in fruit stone trees with emphasis on chlorosis correction mechanisms in orchards:A review. J Plant Nutr 2023;46:782-800. [CrossRef]
2. Hafeez MB, Ramzan Y, Khan S, Ibrar D, Bashir S, Zahra N, et al. Application of zinc and iron-based fertilisers improves the growth attributes, productivity, and grain quality of two wheat (Triticum aestivum) cultivars. Front Nutr 2021;8:779595. [CrossRef]
3. Shakeri S, Saffari M. The status of chemical forms of iron and manganese in various orders of calcareous soils and their relationship with some physicochemical and mineralogical properties. Commun Soil Sci Plant Anal 2020;51:2054-68. [CrossRef]
4. Chao ZF, Chao DY. Similarities and differences in iron homeostasis strategies between graminaceous and nongraminaceous plants. New Phytol 2022;236:1655-60. [CrossRef]
5. Akhtar S, Bangash N, Iqbal MS, Shahzad A, Arshad M, Ul-Hassan F. Comparison of foliar and soil applications for correction of iron deficiency in peanut (Arachis hypogaea L.). Pakistan J Bot 2019:51:1121-7. [CrossRef]
6. Tsai HH, Schmidt W. pH-dependent transcriptional profile changes in iron-deficient Arabidopsis roots. BMC Genomics 2020;21:694. [CrossRef]
7. Mira MM, Asmundson B, Renault S, Hill RD, Stasolla C. Suppression of the soybean (Glycine max) phytoglobin GmPgb1 improves tolerance to iron stress. Acta Physiol Plant 2021;43:147. [CrossRef]
8. Gavassi MA, Alves FR, Monteiro CC, Gaion LA, Alves LR, de Mello Prado R, et al. Photomorphogenic tomato mutants high-pigment 1 and aurea responses to iron deficiency. Sci Hortic 2023;307:111502. [CrossRef]
9. Martín-Barranco A, Thomine S, Vert G, Zelazny E. A quick journey into the diversity of iron uptake strategies in photosynthetic organisms. Plant Signal Behav 2021;16:1975088. [CrossRef]
10. Mehrotra R, Verma RK, Pal A. Iron deficiency chlorosis in aromatic grasses--A review. Environ Challenges 2022;9:100646. [CrossRef]
11. Liang S, Li YF, Tang J, Wang YJ, Li QY. Study on form distribution of soil iron in western Jilin and its correlation with soil properties. Glob Geol 2016;35:593-600.
12. Taalab AS, Ageeb GW, Siam HS, Mahmoud SA. Some characteristics of calcareous soils. A review. Middle East J Agric Res 2019;8:96-105.
13. Murgia I, Marzorati F, Vigani G, Morandini P. Plant iron nutrition:The long road from soil to seeds. J Exp Bot 2022;73:1809-24. [CrossRef]
14. Lindsay WL. Soil and plant relationships associated with iron deficiency with emphasis on nutrient interactions. J Plant Nutr 1984;7:489-500. [CrossRef]
15. de Marcos Lapaz A, Yoshida CH, Gorni PH, de Freitas-Silva L, de Oliveira Araújo T, Ribeiro C. Iron toxicity:Effects on the plants and detoxification strategies. Acta Bot Brasilica 2022;36:e2021abb0131. [CrossRef]
16. Kouas S, Slatni T, Chihaoui SA, Abdelly C, Mhadhbi H. Differential behavior of Medicago truncatula to calcareous soil is explained by modulation of stomatal responses, antioxidant activity, and iron use efficiency. Arab J Geosci 2021;14:1669. [CrossRef]
17. Tomasi D, Marcuzzo P, Nardi T, Lonardi A, Lovat L, Flamini R, et al. Influence of soil chemical features on aromatic profile of V. vinifera cv. Corvina grapes and wines:A study-case in Valpolicella area (Italy) in a calcareous and non-calcareous soil. Agriculture 2022;12:1980. [CrossRef]
18. Merry R, Dobbels AA, Sadok W, Naeve S, Stupar RM, Lorenz AJ. Iron deficiency in soybean. Crop Sci 2022;62:36-52. [CrossRef]
19. Buturi CV, Mauro RP, Fogliano V, Leonardi C, Giuffrida F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021;10:223. [CrossRef]
20. Malhotra H, Pandey R, Sharma S, Bindraban PS. Foliar fertilisation:Possible routes of iron transport from leaf surface to cell organelles. Arch Agron Soil Sci 2020;66:279-300. [CrossRef]
21. Dey S, Regon P, Kar S, Panda SK. Chelators of iron and their role in plant's iron management. Physiol Mol Biol Plants 2020;26:1541-9. [CrossRef]
22. Sarashgi A, Puschenreiter M, Baune M, Paffrath V, Oburger E, Giehl RF, et al. Does the exudation of coumarins from Fe-deficient, soil-grown Brassicaceae species play a significant role in plant Fe nutrition?Rhizosphere 2021;19:100410. [CrossRef]
23. Vélez-Bermúdez IC, Schmidt W. How plants recalibrate cellular iron homeostasis. Plant Cell Physiol 2022;63:154-62. [CrossRef]
24. Liang G. Iron uptake, signaling, and sensing in plants. Plant Commun 2022;3:100349. [CrossRef]
25. Fan Z, Wu Y, Zhao L, Fu L, Deng L, Deng J, et al. MYB308-mediated transcriptional activation of plasma membrane H+-ATPase 6 promotes iron uptake in citrus. Hortic Res 2022;9:uhac088. [CrossRef]
26. Rajniak J, Giehl RF, Chang E, Murgia I, von Wirén N, Sattely ES. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat Chem Biol 2018;14:442-50. [CrossRef]
27. Cui M, Gu M, Lu Y, Zhang Y, Chen C, Ling HQ, et al. Glutamate synthase 1 is involved in iron-deficiency response and long-distance transportation in Arabidopsis. J Integr Plant Biol 2020;62:1925-41. [CrossRef]
28. Grant-Grant S, Schaffhauser M, Baeza-Gonzalez P, Gao F, Conéjéro G, Vidal EA, et al. B3 transcription factors determine iron distribution and FERRITIN gene expression in embryo but do not control total seed iron content. Front Plant Sci 2022;13:1031. [CrossRef]
29. Sági-Kazár M, Solymosi K, Solti Á. Iron in leaves:Chemical forms, signalling, and in-cell distribution. J Exp Bot 2022;73:1717-34. [CrossRef]
30. Rukmani N, Chitdeshwari T, Malarvizhi P, Maheswari M, Karthikeyan S, Marimuthus S. Changes in biochemical constituents and antioxidant enzyme activity in groundnut (Arachis hypogaea L.) by the addition of coated multi-nutrient fertilization in calcareous soil. J Appl Nat Sci 2022;14:1100-9. [CrossRef]
31. El-Sonbaty AE, Farouk S, Al-Yasi HM, Ali EF, Abdel-Kader AA, El-Gamal SM. Enhancement of rose scented geranium plant growth, secondary metabolites, and essential oil components through foliar applications of iron (Nano, Sulfur and Chelate) in alkaline soils. Agronomy 2022;12:2164. [CrossRef]
32. Turan M, Ekinci M, Kul R, Kocaman A, Argin S, Zhirkova AM, et al. Foliar applications of humic substances together with Fe/Nano Fe to increase the iron content and growth parameters of spinach (Spinacia oleracea L.). Agronomy 2022;12:2044. [CrossRef]
33. Barhoumi S, Ellouzi H, Krouma A. Functional analysis of the genotypic differences in response of pea (Pisum sativum L.) to calcareous-induced iron deficiency. Phyton Int J Exp Bot 2023;92:521-36. [CrossRef]
34. Krouma A. Potential of animal manure amendments in combating calcareous induced iron deficiency in pearl millet. Plant Stress 2023;7:100139. [CrossRef]
35. Ishimaru Y, Suzuki M, Tsukamoto T, Suzuki K, Nakazono M, Kobayashi T, et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J 2006;45:335-46. [CrossRef]
36. Yamagata A, Murata Y, Namba K, Terada T, Fukai S, Shirouzu M. Uptake mechanism of iron-phytosiderophore from the soil based on the structure of yellow stripe transporter. Nat Commun 2022;13:7180. [CrossRef]
37. Pérez-Labrada F, Benavides-Mendoza A, Juárez-Maldonado A, Solís-Gaona S, González-Morales S. Effects of citric acid and humic-like substances on yield, enzyme activities, and expression of genes involved in iron uptake in tomato plants. Horticulturae 2023;9:630. [CrossRef]
38. Zhao Y, Liu S, Li F, Sun M, Liang Z, Sun Z, et al. The low ferric chelate reductase activity and high apoplastic pH in leaves cause iron deficiency chlorosis in “Huangguan“pears grafted onto quince A grown in calcareous soil. Sci Hortic 2023;310:111754. [CrossRef]
39. Marastoni L, Lucini L, Miras-Moreno B, Trevisan M, Sega D, Zamboni A, et al. Changes in physiological activities and root exudation profile of two grapevine rootstocks reveal common and specific strategies for Fe acquisition. Sci Rep 2020;10:18839. [CrossRef]
40. Zhao Y, Sun M, Liang Z, Li H, Yu F, Liu S. Analysis of contrast iron chlorosis tolerance in the pear cv. “Huangguan“grafted onto Pyrus betulifolia and quince A grown in calcareous soils. Sci Hortic 2020;271:109488. [CrossRef]
41. Mestre L, Reig G, Betrán JA, Pinochet J, Moreno MÁ. Influence of peach-almond hybrids and plum-based rootstocks on mineral nutrition and yield characteristics of “Big Top“nectarine in replant and heavy-calcareous soil conditions. Sci Hortic 2015;192:475-81. [CrossRef]
42. Mestre L, Reig G, Betrán JA, Moreno MA. Influence of plum rootstocks on agronomic performance, leaf mineral nutrition and fruit quality of “catherina“peach cultivar in heavy-calcareous soil conditions. Spanish J Agric Res 2017;15:e0901. [CrossRef]
43. Font i Forcada C, Reig G, Mestre L, Mignard P, Betrán JA, Moreno MÁ. Scion ×rootstock response on production, mineral composition and fruit quality under heavy-calcareous soil and hot climate. Agronomy 2020;10:1159. [CrossRef]
44. Reig G, Garanto X, Mas N, Iglesias I. Long-term agronomical performance and iron chlorosis susceptibility of several Prunus rootstocks grown under loamy and calcareous soil conditions. Sci Hortic 2020;262:109035. [CrossRef]
45. Reig G, Iglesias I, Zazurca L, Torguet L, Martinez G, Miarnau X. Physiological and agronomical responses of “Vairo“almond and “Big Top“nectarine cultivars grafted onto different Prunus rootstocks and grown under semiarid mediterranean conditions. Agronomy 2022;12:821. [CrossRef]
46. Kangarshahi AA, Amiri NA. Performance of miyagawa satsuma mandarin raised on swingle citrumelo in calcareous soils. Int J Hortic Sci Technol 2023;10:161-72.
47. Dai J, Qiu W, Wang N, Nakanishi H, Zuo Y. Comparative transcriptomic analysis of the roots of intercropped peanut and maise reveals novel insights into peanut iron nutrition. Plant Physiol Biochem 2018;127:516-24. [CrossRef]
48. Qiu W, Wang N, Dai J, Wang T, Zuo Y. Physiological and molecular mechanisms of improved iron nutrition of peanut intercropping with maise. Chinese Sci Bull 2019;64:1129-36. [CrossRef]
49. Chen X, Wei X, Hao M, Zhao J. Changes in soil iron fractions and availability in the Loess Belt of Northern China after 28 years of continuous cultivation and fertilisation. Pedosphere 2019;29:123-31. [CrossRef]
50. Jiao N, Wang F, Ma C, Zhang F, Jensen ES. Interspecific interactions of iron and nitrogen use in peanut (Arachis hypogaea L.)-maise (Zea mays L.) intercropping on a calcareous soil. Eur J Agron 2021;128:126303. [CrossRef]
51. Dai J, Qiu W, Wang N, Wang T, Nakanishi H, Zuo Y. From leguminosae/gramineae intercropping systems to see benefits of intercropping on iron nutrition. Front Plant Sci 2019;10:605. [CrossRef]
52. Kaur H, Manchanda JS. Copper induced iron deficiency in wheat (Triticum aestivum L). J Plant Nutr 2019;42:2824-43. [CrossRef]
53. Kawakami Y, Bhullar NK. Potential implications of interactions between Fe and S on cereal fe biofortification. Int J Mol Sci 2020;21:2827. [CrossRef]
54. Rajaie M, Tavakoly AR. Iron and/or acid foliar spray versus soil application of Fe-EDDHA for prevention of iron deficiency in Valencia orange grown on a calcareous soil. J Plant Nutr 2018;41:150-8. [CrossRef]
55. Vetrivel M, Devi DD, Sivakumar R, Chinnasamy GP. Impact of different levels of iron on mitigation of iron chlorosis in varagu CO3 (Paspalum scrobiculatum L.). J Appl Nat Sci 2022;14:206-12. [CrossRef]
56. Ramzani PM, Khalid M, Naveed M, Ahmad R, Shahid M. Iron biofortification of wheat grains through integrated use of organic and chemical fertilisers in pH affected calcareous soil. Plant Physiol Biochem 2016;104:284-93. [CrossRef]
57. Souri B, Sayadi Z. Efficiency of sulfur-bentonite granules to improve uptake of nutrient elements by the crop plant cultivated in calcareous soil. Commun Soil Sci Plant Anal 2021;52:2414-30. [CrossRef]
58. Zebec V, Lisjak M, Jovi?J, Kujundži?T, Rastija D, Lon?ari?Z. Vineyard fertilisation management for iron deficiency and chlorosis prevention on carbonate soil. Horticulturae 2021;7:285. [CrossRef]
59. Ye JY, Zhou M, Zhu QY, Zhu YX, Du WX, Liu XX, et al. Inhibition of shoot-expressed NRT1.1 improves reutilization of apoplastic iron under iron-deficient conditions. Plant J 2022;112:549-64. [CrossRef]
60. Chen M, Zhang S, Liu L, Liu J, Ding X. Organic fertilisation increased soil organic carbon stability and sequestration by improving aggregate stability and iron oxide transformation in saline-alkaline soil. Plant Soil 2022;474:233-49. [CrossRef]
61. D'Amato R, De Feudis M, Troni E, Gualtieri S, Soldati R, Famiani F, et al. Agronomic potential of two different glass-based materials as novel inorganic slow-release iron fertilisers. J Sci Food Agric 2022;102:1660-4. [CrossRef]
62. Narayanasamy R, Thiyagarajan C, Pillai MP, Muthunalliappan M, Subburamu K, Subramanian M. Nutrient release from biodegradable polymer-coated multi-nutrient fertiliser granules in calcareous soils. Arab J Geosci 2023;16:53. [CrossRef]
63. Flis S, Bowman M. Soil health and 4R:What practices are working?Crop Soils 2021;54:21-5. [CrossRef]
64. Salari-Bardsiri N, Hejazi-Mehrizi M, Behrouz R, Naghavi H, Fekri M. Release kinetics and bioavailability of nutrients from a slow-release iron fertiliser coated with cellulose derived from waste palm branches and pistachio shells. Arab J Geosci 2023;16:120. [CrossRef]
65. Abbaszadeh-Dahaji P, Masalehi F, Akhgar A. Improved growth and nutrition of sorghum (Sorghum bicolor) plants in a low-fertility calcareous soil treated with plant growth-promoting rhizobacteria and Fe-EDTA. J Soil Sci Plant Nutr 2020;20:31-42. [CrossRef]
66. Martín-Fernández C, López-Rayo S, Hernández-Apaolaza L, Lucena JJ. Timing for a sustainable fertilisation of Glycine max by using HBED/Fe3+ and EDDHA/Fe3+ chelates. J Sci Food Agric 2017;97:2773-81. [CrossRef]
67. Ramezanpour MR, Farajpour M, Yousefian M. Enhancing the quality and quantity of date palm (Phoenix dactylifera) by micronutrients under calcareous soils with a high pH level. J Nat Fibers 2022;19:11749-62. [CrossRef]
68. Turhan A, Özmen N. Effects of iron fertilisation on plant growth, yield components and quality traits of industrial tomatoes. Mediterr Agric Sci 2022;35:1-6. [CrossRef]
69. Ay AY, Demirkaya S, K?z?lkaya R, Gülser C. The effects of two Fe-EDDHA chelated fertilisers on dry matter production and Fe uptake of tomato seedlings and Fe forms of a calcareous soil. Eurasian J Soil Sci 2022;11:259-65. [CrossRef]
70. Khakpour S, Hajizadeh HS, Hemati A, Bayanati M, Nobaharan K, Chelan EM, et al. The effect of pre-harvest treatment of calcium nitrate and iron chelate on post-harvest quality of apple (Malus domestica Borkh cv. Red Delicious). Sci Hortic 2022;304:111351. [CrossRef]
71. Zhang R, Zhang W, Kang Y, Shi M, Yang X, Li H, et al. Application of different foliar iron fertilisers for improving the photosynthesis and tuber quality of potato (Solanum tuberosum L.) and enhancing iron biofortification. Chem Biol Technol Agric 2022;9:79. [CrossRef]
72. Puente-Ramírez JV, Rivera-Ortiz P, Silva-Espinosa JH, Andrade-Limas E. Chelate EDDHA to correct the iron deficiency in Italian lemon trees (Citrus limon (L.) Osbeck). Terra Latinoam 2022;40:e926. [CrossRef]
73. Ferreira CM, Sousa CA, Sanchis-Pérez I, López-Rayo S, Barros MT, Soares HM, et al. Calcareous soil interactions of the iron(III) chelates of DPH and Azotochelin and its application on amending iron chlorosis in soybean (Glycine max). Sci Total Environ 2019;647:1586-93. [CrossRef]
74. López-Rayo S, Sanchis-Pérez I, Ferreira CM, Lucena JJ. [S,S]-EDDS/Fe:A new chelate for the environmentally sustainable correction of iron chlorosis in calcareous soil. Sci Total Environ 2019;647:1508-17. [CrossRef]
75. Santos CS, Rodrigues E, Ferreira S, Moniz T, Leite A, Carvalho SM, et al. Foliar application of 3-hydroxy-4-pyridinone Fe-chelate [Fe(mpp)3] induces responses at the root level amending iron deficiency chlorosis in soybean. Physiol Plant 2021;173:235-45. [CrossRef]
76. Suzuki M, Urabe A, Sasaki S, Tsugawa R, Nishio S, Mukaiyama H, et al. Development of a mugineic acid family phytosiderophore analog as an iron fertiliser. Nat Commun 2021;12:1558. [CrossRef]
77. Ueno D, Ito Y, Ohnishi M, Miyake C, Sohtome T, Suzuki M. A synthetic phytosiderophore analog, proline-2'-deoxymugineic acid, is efficiently utilised by dicots. Plant Soil 2021;469:123-34. [CrossRef]
78. Wang T, Wang N, Lu Q, Lang S, Wang K, Niu L, et al. The active Fe chelator proline-2'-deoxymugineic acid enhances peanut yield by improving soil Fe availability and plant Fe status. Plant Cell Environ 2022;46:239-50. [CrossRef]
79. Kong WL, Wen TY, Wang YH, Wu XQ. Physiological and transcriptome analyses revealed the mechanism by which deferoxamine promotes iron absorption in Cinnamomum camphora. Int J Mol Sci 2022;23:9854. [CrossRef]
80. López-Salazar R, Peña-Ramos FM, Sánchez-Bernal F, Lozano-Cavazos CJ, Benavides-Mendoza A, González-Fuentes JA. Efecto de un fulvato de hierro sobre calidad y producción de frutos de chile Serrano. Rev Mex Cienc Agrícolas 2019;10:1367-78. [CrossRef]
81. de Santiago A, García-López AM, Recena R, Moreno MT, Carmona E, Delgado A. Adsorption of humic substances on ferrihydrite affects its use as iron source by plants. Agric Food Sci 2020;29:451-9. [CrossRef]
82. Cieschi MT, Lucena JJ. Leonardite iron humate and synthetic iron chelate mixtures in Glycine max nutrition. J Sci Food Agric 2021;101:4207-19. [CrossRef]
83. Zanin L, Tomasi N, Cesco S, Varanini Z, Pinton R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front Plant Sci 2019;10:675. [CrossRef]
84. Pérez-Labrada F, Benavides-Mendoza A, Juárez-Maldonado A, Solís-Gaona S, González-Morales S. Organic acids combined with Fe-chelate improves ferric nutrition in tomato grown in calcisol soil. J Soil Sci Plant Nutr 2020;20:673-83. [CrossRef]
85. Olego MÁ, Lasso MC, Quiroga MJ, Visconti F, López R, Garzón-Jimeno E. Effects of Leonardite amendments on vineyard calcareous soil fertility, vine nutrition and grape quality. Plants (Basel) 2022;11:356. [CrossRef]
86. Sharma S, Anand N, Bindraban PS, Pandey R. Foliar application of humic acid with fe supplement improved rice, soybean, and lettuce iron fortification. Agriculture 2023;13:132. [CrossRef]
87. Salahi B, Hadavi E, Samar SM. Foliar iron sulphate-organic acids sprays improve the performance of oriental plane tree in calcareous soil better than soil treatments. Urban For Urban Green 2017;21:175-82. [CrossRef]
88. Souri MK, Sooraki FY, Moghadamyar M. Growth and quality of cucumber, tomato, and green bean under foliar and soil applications of an aminochelate fertiliser. Hortic Environ Biotechnol 2017;58:530-6. [CrossRef]
89. Gattullo CE, Pii Y, Allegretta I, Medici L, Cesco S, Mimmo T, et al. Iron mobilisation and mineralogical alterations induced by iron-deficient cucumber plants (Cucumis sativus L.) in a calcareous soil. Pedosphere 2018;28:59-69. [CrossRef]
90. Islas-Valdez S, López-Rayo S, Arcos J, Menéndez N, Lucena JJ. Effect of Fe:ligand ratios on hydroponic conditions and calcareous soil in Solanum lycopersicum L. and Glycine max L. fertilised with heptagluconate and gluconate. J Sci Food Agric 2020;100:1106-117. [CrossRef]
91. Mirbolook A, Rasouli-Sadaghiani MH, Sepehr E, Lakzian A, Hakimi M. Fortification of bread wheat with iron through soil and foliar application of iron-organic-complexes. J Plant Nutr 2020;44:1386-403. [CrossRef]
92. Subramani M, Durairaj J, Thiyagarajan C, Muthumani J. Synthesis of iron chelates for remediation of iron deficiency in an alkaline and calcareous soil. J Appl Nat Sci 2021;13:149-55. [CrossRef]
93. Ek?i HS, Sönmez ?. Effects of amino acid applications on yield, growth and mineral nutrition of greenhouse tomato. J Elem 2022;27:545-57.
94. Gheshlaghi Z, Khorassani R, Abadia J. Two Fe mining sub-products and three thiol compounds alleviate Fe deficiency in soybean (Glycine max L.) grown in a calcareous soil in greenhouse conditions. Plant Soil 2023;482:469-90. [CrossRef]
95. Mazhar MW, Akram R, Shahid A. Foliar application of iron glutamate improves yield and growth of tomatoes compared to iron sulphate and L-glutamate. Int J Veg Sci 2022;28:511-20. [CrossRef]
96. Pérez-Labrada F, Benavidez-Mendoza A, Valdez-Aguilar LA, Robledo-Torres V. Citric acid in the nutrient solution increases the mineral absorption in potted tomato grown in calcareous soil. Pak J Bot 2016;48:67-74.
97. Çoban GA, Aras S. Effects of ascorbic and oxalic acids on cucumber seedling growth and quality under mildly limey soil conditions. In:Gesunde Pflanz. Berlin:Springer;2022. p. 1-8.
98. Dhankhar R, Gupta S, Gulati P. Insights on Plant-Microbe Interactions in Soil in Relation to Iron Dynamics. Vegetos. Berlin:Springer Nature Singapore;2022. 1-18. [CrossRef]
99. Ferreira CM, López-Rayo S, Lucena JJ, Soares EV, Soares HM. Evaluation of the efficacy of two new biotechnological-based freeze-dried fertilisers for sustainable fe deficiency correction of soybean plants grown in calcareous soils. Front Plant Sci 2019;10:1335. [CrossRef]
100. Lurthy T, Cantat C, Jeudy C, Declerck P, Gallardo K, Barraud C, et al. Impact of bacterial siderophores on iron status and ionome in pea. Front Plant Sci 2020;11:730. [CrossRef]
101. Abiraami TV, Suman A, Singh B, Aswini K, Annapurna K. Radiochemical evidence for the contribution of chemotyped siderophore producing bacteria towards plant iron nutrition. Curr Microbiol 2021;78:4072-83. [CrossRef]
102. Puglisi I, Brida S, Stoleru V, Torino V, Sellitto VM, Baglieri A. Application of novel microorganism-based formulations as alternative to the use of iron chelates in strawberry cultivation. Agriculture 2021;11:217. [CrossRef]
103. Roriz M, Pereira SI, Castro PM, Carvalho SM, Vasconcelos MW. Iron metabolism in soybean grown in calcareous soil is influenced by plant growth-promoting rhizobacteria - A functional analysis. Rhizosphere 2021;17:100274. [CrossRef]
104. Kong WL, Wang YH, Lu LX, Li PS, Zhang Y, Wu XQ. Rahnella aquatilis JZ-GX1 alleviates iron deficiency chlorosis in Cinnamomum camphora by secreting desferrioxamine and reshaping the soil fungal community. Front Plant Sci 2022;13:960750. [CrossRef]
105. Singh S, Annapurna K, Shrivastava N, Varma A. Symbiotic interplay of Piriformospora indica and Azotobacter chroococcum augments crop productivity and biofortification of Zinc and Iron. Microbiol Res 2022;262:127075. [CrossRef]
106. Daliran T, Halajnia A, Lakzian A. Thiobacillus bacteria-enhanced iron biofortification of soybean in a calcareous soil enriched with ferrous sulfate, mill scale, and pyrite. J Soil Sci Plant Nutr 2022;22:2221-34. [CrossRef]
107. Awad AA, Ahmed AI, Elazem AH, Sweed AA. Mitigation of CaCO3 Influence on Ipomoea batatas plants using Bacillus megaterium DSM 2894. Agronomy 2022;12:1571. [CrossRef]
108. Vahedi R, Rasouli-Sadaghiani MH, Barin M, Vetukuri RR. Effect of biochar and microbial inoculation on P, Fe, and Zn bioavailability in a calcareous soil. Processes 2022;10:343. [CrossRef]
109. Zahedifar M. Iron fractionation in the calcareous soils of different land uses as influenced by biochar. Waste Biomass Valorization 2020;11:2321-30. [CrossRef]
110. Harbort CJ, Hashimoto M, Inoue H, Niu Y, Guan R, RombolàAD, et al. Root-secreted coumarins and the microbiota interact to improve iron nutrition in Arabidopsis. Cell Host Microbe 2020;28:825-37.e6. [CrossRef]
111. Philipo M, Ndakidemi PA, Mbega ER. Multilocation dataset on seed Fe and Zn contents of bean (Phaseolus vulgaris L.) genotypes grown in Tanzania. Data Brief 2020;31:105664. [CrossRef]
112. Salhi K, Hajlaoui H, Krouma A. Genotypic differences in response of durum wheat (Triticum durum Desf.) to lime-induced iron chlorosis. Plant Direct 2022;6:e377. [CrossRef]
113. Masuda H, Aung MS, Kobayashi T, Hamada T, Nishizawa NK. Enhancement of iron acquisition in rice by the mugineic acid synthase gene with ferric iron reductase gene and OsIRO2 confers tolerance in submerged and nonsubmerged calcareous soils. Front Plant Sci 2019;10:1179. [CrossRef]
114. Kawakami Y, Gruissem W, Bhullar NK. Novel rice iron biofortification approaches using expression of ZmYS1 and OsTOM1 controlled by tissue-specific promoters. J Exp Bot 2022;73:5440-59. [CrossRef]
115. Kobayashi T, Maeda K, Suzuki Y, Nishizawa NK. Simultaneous enhancement of iron deficiency tolerance and iron accumulation in rice by combining the knockdown of OsHRZ ubiquitin ligases with the introduction of engineered ferric-chelate reductase. Rice (N Y) 2022;15:54. [CrossRef]
116. Tayade AD, Motagi BN, Jadhav MP, Nadaf AS, Koti RV, Gangurde SS, et al. Genetic mapping of tolerance to iron deficiency chlorosis in peanut (Arachis hypogaea L.). Euphytica 2022;218:46. [CrossRef]
117. Wang B, Wei H, Chen Z, Li Y, Zhang WH. Carbonate-induced chemical reductants are responsible for iron acquisition in strategy i wild herbaceous plants native to calcareous grasslands. Plant Cell Physiol 2022;63:770-84. [CrossRef]
118. Ibrahim S, Saleem B, Rehman N, Zafar SA, Naeem MK, Khan MR. CRISPR/Cas9 mediated disruption of Inositol Pentakisphosphate 2-Kinase 1 (TaIPK1) reduces phytic acid and improves iron and zinc accumulation in wheat grains. J Adv Res 2022;37:33-41. [CrossRef]
119. Sutariya BP, Vyas TK, Faldu PR, Patel KG, Vala AK. A Microcosm study on effect of iron nanoparticles on paddy (Oryza sativa) Growth. J Inorg Organomet Polym Mater 2021;31:2425-35. [CrossRef]
120. Chugh G, Siddique KH, Solaiman ZM. Iron fortification of food crops through nanofertilisation. Crop Pasture Sci 2022;73:736-48. [CrossRef]
121. Rai P, Sharma S, Tripathi S, Prakash V, Tiwari K, Suri S, et al. Nanoiron:Uptake, translocation and accumulation in plant systems. Plant Nano Biol 2022;2:100017. [CrossRef]
122. Cieschi MT, Polyakov AY, Lebedev VA, Volkov DS, Pankratov DA, Veligzhanin AA, et al. Eco-friendly iron-humic nanofertilizers synthesis for the prevention of iron chlorosis in soybean (Glycine max) grown in calcareous soil. Front Plant Sci 2019;10:413. [CrossRef]
123. Gülser F, Yavuz H?, Gökkaya TH, Sedef M. Effects of iron sources and doses on plant growth criteria in soybean seedlings. Eurasian J Soil Sci 2019;8:298-303. [CrossRef]
124. Sabet H, Mortazaeinezhad F. Yield, growth and Fe uptake of cumin (Cuminum cyminum L.) affected by Fe-nano, Fe-chelated and Fe-siderophore fertilisation in the calcareous soils. J Trace Elem Med Biol 2018;50:154-60. [CrossRef]
125. Saleem I, Maqsood MA, Aziz T, Bhatti IA, Jabbar A. Potassium ferrite nano coated dap fertiliser improves wheat (Triticum aestivum l.) growth and yield under alkaline calcareous soil conditions. Pakistan J Agric Sci 2021;58:485-92.
126. Awad AA, Sweed AA, Rady MM, Majrashi A, Ali EF. Rebalance the nutritional status and the productivity of high CaCO3-stressed sweet potato plants by foliar nourishment with zinc oxide nanoparticles and ascorbic acid. Agronomy 2021;11:1443. [CrossRef]
127. Soliemanzadeh A, Fekri M. Effects of green iron nanoparticles on iron changes and phytoavailability in a calcareous soil. Pedosphere 2021;31:761-70. [CrossRef]
128. Taskin MB, Gunes A. Iron biofortification of wheat grains by foliar application of nano zero-valent iron (nZVI) and other iron sources with urea. J Soil Sci Plant Nutr 2022;22:4642-52. [CrossRef]
129. Cao X, Yue L, Wang C, Luo X, Zhang C, Zhao X, et al. Foliar application with iron oxide nanomaterials stimulate nitrogen fixation, yield, and nutritional quality of soybean. ACS Nano 2022;16:1170-81. [CrossRef]
130. Yang YZ, Li MJ, Li XL, Zhou BB, Wei QP, Zhang JK. Preparation of soluble ferrous complex with high alkaline stability by reducing iron nanoparticle in Maillard reaction. Chem Pap 2021;75:3227-39. [CrossRef]
131. Khallizadeh J, Dordipour E, Baranimotlgh M, Gharanjiki A. Effect of iron impregnated wheat straw and particleboard biochar on the iron uptake and growth of two soybean cultivars in a calcareous soil. J Soil Manag Sustain Prod 2020;10:83-100.
132. Zulfiqar U, Maqsood M, Hussain S, Anwar-Ul-Haq M. Iron nutrition improves productivity, profitability, and biofortification of bread wheat under conventional and conservation tillage systems. J Soil Sci Plant Nutr 2020;20:1298-310. [CrossRef]
133. Zhao Y, Li H, Sun M, Liang Z, Yu F, Li F, et al. Effects of soil aeration and fertilisation practices on alleviating iron deficiency chlorosis in “Huangguan“pears grafted onto quince a in calcareous soils. Horticulturae 2021;7:172. [CrossRef]
134. Lampreave M, Mateos A, Valls J, Nadal M, Sánchez-Ortiz A. carbonated irrigation assessment of grapevine growth, nutrient absorption, and sugar accumulation in a tempranillo (Vitis vinifera L.). Vineyard Agric 2022;12:792. [CrossRef]
135. Poonia T, Bhunia SR, Choudhary R. Effect of iron fertilisation on growth, yield and economics of groundnut (Arachis hypogaea L.). Int J Econ Plants 2022;9:38-44. [CrossRef]
136. Porkodi G, Shanmugasundaram R, Saravanapandian P, Swaminathan C. Impact of different sources and levels of iron on yield and quality of groundnut. Pharma Innov J 2022;11:2798-801.
137. Khosroshahi MZ, Nezami S, Hajiamiri A. Rapid correction of iron deficiency in peach trees by direct injection of ferrous sulfate into the trunk. Arch Agron Soil Sci 2023;69:1405-18. [CrossRef]
138. Ayyat AM, Ramadan MH, Gahory A, Abdel-Mola M. Biochar combined with foliar application of chelated and nano iron enhanced qualitative and quantitative attributes of Polianthes tuberosa L. plants. Egypt J Hortic 2023;50:17-33. [CrossRef]
139. ?pek M, Ar?kan ?, E?itken A, P?rlak L, Dönmez MF, Turan M. Influence of bacterial inoculation on growth and plant nutrition of peach grafted in different rootstocks in calcareous soil. Sains Malays 2021;50:2615-24. [CrossRef]
140. Gong Y, Yu L, Zhao L. Ecological networks in agroecosystems:Approaches and applications. Front Agric Sci Eng 2022;9:523-35. [CrossRef]