
1. INTRODUCTION
Food loss due to pest attacks is a significant problem that affects the global food supply chain. Pests, including insects, rodents, and fungi, can damage crops during growth, harvesting, transportation, and storage. This loss not only impacts the availability and affordability of food, but also has significant economic, social, and environmental consequences [1]. The World Health Organization estimates that about one-third of all food produced globally is lost or wasted, with pests contributing significantly to this figure [2]. Agricultural losses caused by pests in America can vary according to many factors, such as crop type, region, and year. However, it is estimated that losses caused by pests can reach billions of dollars each year in the region. For example, in Brazil, the largest agricultural producer in Latin America, it is estimated that losses caused by pests and diseases in crops can reach around US$17.7 billion per year. In the United States, losses caused by pests and diseases in agricultural crops can reach US$120 billion per year [3,4]. In addition to the economic costs associated with food loss, such as decreased income for farmers and increased prices for consumers, there are also environmental costs, including increased greenhouse gas emissions due to food waste decomposition [5]. Therefore, addressing food loss due to pests is critical to ensuring global food security and sustainability [6].
In this context, Bemisia tabaci, commonly known as the silverleaf whitefly, is a significant worldwide agricultural pest that feeds on a wide range of crops, including fruits, vegetables, and ornamental plants [7]. This tiny insect is considered a serious threat to global food security due to the extensive damage it causes to crops, leading to significant yield losses and reduced crop quality [8]. B. tabaci has a piercing-sucking mouthpart that allows it to extract sap from plants, leading to the development of leaf yellowing, deformation, and even death. In addition, this pest is known to transmit more than 100 plant viruses, causing further damage to crops [9,10]. The economic impact of B. tabaci on agriculture is substantial, with billions of dollars lost every year due to crop damage and the cost of pest management. Therefore, effective control measures are critical to reduce the impact of this pest on food production and ensure global food security [11].
There are several pesticides used to control B. tabaci, including neonicotinoids, organophosphates, pyrethroids, and insect growth regulators. However, these pesticides can have adverse effects on human health and the environment [12]. Neonicotinoids are also known to harm non-target organisms such as bees and other beneficial insects, leading to concerns about their impact on the environment and biodiversity [13]. On the other hand, organophosphates and pyrethroids are also commonly used to control B. tabaci. Nonetheless, these pesticides are highly toxic to humans and can cause a range of health problems, including headaches, nausea, dizziness, and respiratory problems. Long-term exposure to these pesticides has been linked to more serious health issues such as cancer, neurological disorders, and developmental problems [14]. Moreover, insect growth regulators, such as methoprene, are a safer alternative to traditional pesticides as they specifically target insect development and do not harm beneficial insects or humans. However, they can still have negative impacts on the environment if not used properly, such as unintended effects on non-target organisms [15]. Due to these several limitations, the use of alternative pest control methods, such as biological control and biopesticides, is currently targeted, aiming at sustainability and the promotion of consumer health [16].
Plant extracts and essential oils (EOs) have been used for centuries as natural pest control methods due to their broad-spectrum activity and lower toxicity compared to synthetic pesticides. These substances are derived from various parts of plants, including leaves, flowers, roots, and seeds, and contain a variety of compounds that have insecticidal and repellent properties [17]. Particularly, the use of EOs in pest management is gaining increasing attention due to concerns over the safety and environmental impact of synthetic pesticides. Several studies have demonstrated the effectiveness of EOs in controlling various agricultural pests, such as aphids, whiteflies, mosquitoes, and bedbugs. In addition, EOs can be used in different ways, such as spraying, fumigation, and application in baits, which increases their versatility and possibility of application in different cultures [18-20]. In addition to their effectiveness in controlling pests, they have the added advantage of being biodegradable, sustainable, and cost-effective, besides minimizing the emergence of pest resistance, due to the variety and natural complexity of their substances [21,22].
In this context, it is necessary to optimize scientific information on the insecticidal action of natural products found in American countries against B. tabaci, so this review will cover and discuss the studies involved with EOs that have relevant action against this important agronomic pest.
2. METHODOLOGY
The research was carried out using the combination of “essential oil” “activity” and “Bemisia tabaci”. The following research databases were used for the review: Science Direct (116 documents), Web of Science (45 documents), Scopus (1601 documents), and Google Scholar (3670 documents). According to the relevance order, repeated articles or those that only cited studies on the subject in question were excluded, so that 118 articles had their content analyzed and only studies with EOs from plants collected in American countries or purchased by laboratories located in America were taken into account. Then, after careful selection, 11 articles about the proposed central theme were included and discussed in the two next sections of the review, which exclude the articles described in the introduction and discussion sections . Tables 1 and 2 summarize the selected articles and their main information.
Table 1: Essential oils obtained in American countries with insecticidal/repellent/deterrent activity against Bemisia tabaci.
| Plant species | Family | EO origin | Main constituents (%) | References |
|---|---|---|---|---|
| Xylopia aromatica | Annonaceae | Leaves | Bicyclogermacrene (44.80), α-pinene (8.23) and β-pinene (7.75) | Peres et al., 2020 |
| Fruits | α-pinene (35.40), β-phellandrene (31.05) and β-pinene (22.51) | |||
| Mansoa alliaceae | Bignoniaceae | ND | Diallyl trisulfide (52.8) and diallyl disulfide (33.9 ) | Santana et al., 2022 |
| Adenocalymma alliaceum | Diallyl trisulfide (66.9) and diallyl sulfide (23.3) | Fanella et al., 2015 | ||
| Pelargonium graveolens | Geraniaceae | Geraniol (42.3) and linalool (16.4) | ||
| Plectranthus neochilus | Lamiaceae | Trans-caryophyllene (30.7), α-thujene (11.7), α-pinene (15.0) and β-pinene (8.3 ) | ||
| Vitex agnus-castus | 1.8-cineole (33.2) and sabinene (22.7) | |||
| Laurus nobilis | Lauraceae | Leaves | ND | Ringuelet et al., 2012 |
| Piper marginatum | Piperaceae | ND | (E)-methyl eugenol (34.7) and (Z)-methyl eugenol (27.5) | Santana et al., 2022 |
| Piper callosum | safrole (29.3), α-pinene (19.2), and β-pinene (14.3) | Fanella et al., 2015 | ||
| Cymbopogon citratus | Poaceae | Leaves | ND | Ringuelet et al., 2012 |
| Zanthoxylum riedelianum | Rutaceae | Fruits | Limonene (29.22), β-myrcene (22.79), bicyclogermacene (18.13) and germacrene D (14.40) | Pereira et al., 2018 |
| γ-elemene (21.2), germacrene D (14.2), sabinene (11.9) and limonene (11.3) | Costa et al., 2017 | |||
| Leaves | γ-elemene (24.81), phytol (18.16), bicyclogermacrene (16.18), cis-nerolidol (8.26), and D-germacrene (6.52) | Christofoli et al., 2022 | ||
| Zanthoxylum rhoifolium | Fruits | β-phellandrene (76.8) | Pereira et al., 2022 | |
| Sabinene (55.9), germacrene D (17.1) and β-myrcene (8.03) | Costa et al., 2017 | |||
| Leaves | Whereas β-elemene (31.26), D-germacrene (18.16), β-caryophyllene (12.09), δ-elemene (7.63), β-cedrene (6.69), bicyclogermacrene (4.57) and E-caryophyllene (3.63) | Christofoli et al., 2015 | ||
| Citrus aurantifolia | Peels | Limonene (37.73), β-pinene (9.89), α-terpineol (5.04) | Ribeiro et al., 2020 | |
| Citrus limon | Limonene (40.70), β-pinene (18.14), α-terpineol (2.78) | |||
| Citrus reticulata | Limonene (70.96), myrcene (4.61) | |||
| Citrus sinensis×reticulata | Limonene (77.79), p-Mentha-2,4 (8)-diene (9.80 ), myrcene (6.50) | |||
| Clonorchis sinensis var. pear | ND | Ribeiro et al., 2010 | ||
| Citrus aurantium |
EO: Essential oil.
Table 2: Biological activity of America-available essential oils on Bemisia tabaci.
| Plant species | EO origin | Biological activity | References |
|---|---|---|---|
| Xylopia aromatica | Leaves | Deterrent activity at 2%: 98% reduction | Peres et al., 2020 |
| Fruits | Deterrent activity at 2%: 96% reduction | ||
| Mansoa alliaceae | ND | Nymph mortality: LC50=10.99 μL/mL and LC90=22.25 μL/mL, after 5 days of exposure), also observed in semifield condition (90.54% efficacy). Also presents ovicidal, repellent and deterrent activities | Santana et al., 2022 |
| Adenocalymma alliaceum | Nymph insecticidal activity: LC50=0.4 and 0.8 μL/L air in 72 and 3 h, respectively. Deterrent activity: 71.1% reduction. Anti-settlement activity on tomato: 50% reduction | Fanella et al., 2015 | |
| Pelargonium graveolens | Adult insecticidal activity: LC50=0.6 μL/L air in 24 h. Deterrent activity: 90.9% reduction. Anti-settlement activity on tomato: 73.6% reduction | ||
| Plectranthus neochilus | Deterrent activity: 61.5% reduction. Anti-settlement activity on tomato: 34.5% reduction | ||
| Vitex agnus-castus | Adult insecticidal activity: LC50=1.2 μL/L air in 24 h. Deterrent activity: 90.9% reduction. Anti-settlement activity on tomato: 73.6% reduction | ||
| Laurus nobilis | Leaves | Repellence (73.1%) and mortality (53%) activities, after 24 h (5% concentration). With C. citratus EO 1:1 proportion: Repellence (84.3%) and mortality (55.5%) | Ringuelet et al., 2012 |
| Piper marginatum | ND | Nymph mortality (LC50=9.39 μL/mL and LC90=20.79 μL/mL, after 5 days of exposure), also observed in semifield condition (91.51% efficacy). Also presents ovicidal, repellent and deterrent activities | Santana et al., 2022 |
| Piper callosum | Deterrent activity: 91.5% reduction. Anti-settlement activity on tomato: 70.8% reduction | Fanella et al., 2015 | |
| Cymbopogon citratus | Leaves | Repellence (77.3%) and mortality (31.2%) activities, after 24 h (5% concentration). With Laurus nobilis EO 1: 1 proportion: Repellence (84.3%) and mortality (55.5%) | Ringuelet et al., 2012 |
| Zanthoxylum riedelianum | Fruits | Insecticidal activity (1.5%): 70% reduction; Larvicidal activity: 91.23% against 2nd instar nymphae | Pereira et al., 2018 |
| Deterrent activity at 1 and 2%: 95 and 98% reduction, respectively | Costa et al., 2017 | ||
| Leaves | Nymph insecticidal activity at 5%; 100%, after 16 days of treatment. Deterrent activity: 97% reduction, after 6 days of treatment | Christofoli et al., 2022 | |
| Zanthoxylum rhoifolium | Fruits | Larvicidal activity (1.5%): 91% against 2nd instar nymphae. Deterrent activity: 71% reduction | Pereira et al., 2022 |
| Deterrent activity at 1 and 2%: 95 and 98% reduction, respectively | Costa et al., 2017 | ||
| Leaves | Nymph insecticidal activity at 5%; 100%, after 16 days of treatment. Deterrent activity: 98% reduction, after 6 days of treatment | Christofoli et al., 2015 | |
| Citrus aurantifolia | Peels | Fumigant activity (LC50=0.70 μL/L air). Deterrent activity: 85.51%, 0.125 μL/L air | Ribeiro et al., 2020 |
| Citrus limon | Fumigant activity (LC50=1.77 μL/L air). Deterrent activity: 86.34%, 0.8 μL/L air | ||
| Citrus reticulata | Fumigant activity (LC50=3.04 μL/L air). Deterrent activity: 82.71%, 1 μL/L air | ||
| Citrus sinensis×reticulata | Fumigant activity (LC50=5.39 μL/L air). Deterrent activity: 94.93%, 3 μL/L air | ||
| Clonorchis sinensis var. pear | Fumigant activity (LC50=3.80 μL/L air). Deterrent activity (IC50=4 μL/L air) | Ribeiro et al., 2010 | |
| Citrus aurantium | Fumigant activity (LC50=5.80 μL/L air). Deterrent activity (IC50=5 μL/L air) |
EO: Essential oil
3. RESULTS
3.1. EOs Found in America with Insecticidal Activity Against Bemicia Tabaci
3.1.1. Annonaceae
Free and nanoencapsulated EOs from Xylopia aromatica leaves and fruits decreased the oviposition of B. tabaci in common bean leaves. At a concentration of 2%, the free EO from leaves and fruits caused up to 98 and 96% deterrent activity, respectively, whereas the encapsulated EOs reached 91 and 88% activity, respectively. These results were considered very high since the positive control (pyriproxifen 1%) caused less than 10% deterrent activity. For the leaves EO, the major compounds were bicyclogermacrene (44.80%), α-pinene (8.23%), and β-pinene (7.75%), while in fruits they were α-pinene (35.40%), β-phellandrene (31.05%), and β-pinene (22.51%) [23].
3.1.2. Bignoniaceae
Mansoa alliaceae EO caused nymph mortality (LC50 = 10.99 μL/mL and LC90 = 22.25 μL/mL, after 5 days of exposure), which was also observed in the semifield condition (90.54% efficacy). Still, the EO demonstrated a reduction in cotton pest colonization as well as ovicidal, repellent, and deterrent activities. The major constituents of the oil were diallyl trisulfide (52.8%) and diallyl disulfide (33.9%) [24]. The EO from Adenocalymma alliaceum was also effective against the B. tabaci type B tomato settlement (50% reduction), besides producing an oviposition deterrent effect (71.1% reduction). Moreover, the EO showed insecticide activity against nymphs and adults, with a LC50 equal to 0.4 and 0.8 μL/L air in 72 and 3 h, respectively. Its main compounds were diallyl trisulfide (66.9%) and diallyl sulfide (23.3%) [25].
3.1.3. Geraniaceae
The EO from Pelargonium graveolens was effective against the B. tabaci type B tomato settlement (73.6% reduction), besides producing an oviposition deterrent effect (90.9% reduction). Moreover, the EO showed insecticide activity against adult insects, with a LC50 equal to 0.6 μL/L air in 24 h. Its main compounds were the alcohol monoterpenes geraniol (42.3%) and linalool (16.4%) [25].
3.1.4. Lamiaceae
The EO from Plectranthus neochilus was effective against the B. tabaci type B tomato settlement (34.5% reduction), besides producing an oviposition deterrent effect (61.5% reduction). Its main compounds were trans-caryophyllene (30.7%), as well as the bicyclic monoterpenes α-thujene (11.7%), α-pinene (15.0%), and β-pinene (8.3%). In the same study, it was described the adult insecticidal activity of Vitex agnus castus, presenting an LC50 equal to 1.2 μL/L air in 24 hours, whereas its major constituents were the monoterpenes 1.8-cineole (33.2%) and sabinene (22.7%) [25].
3.1.5. Lauraceae
The study of Ringuelet et al. (2012) demonstrated the effect of Laurel nobilis EO 5% on B. tabaci, leading to its repellence (73.1%) and mortality (53%) after 24 h, with five daily applications. The effect was improved when the laurel EO was mixed with C. citratus EO at a 1:1 proportion. With this condition, the repellence index enhances to 84.34%, and the mortality rate reaches 55.5% [26].
3.1.6. Piperaceae
Piper marginatum EO caused nymph mortality (LC50 = 9.39 μL/mL and LC90 = 20.79 μL/mL, after 5 days of exposure), which was also observed in the semifield condition (91.51% efficacy). Still, the EO demonstrated a reduction in cotton pest colonization as well as ovicidal, repellent, and deterrent activities. The major constituents of the oil were (E)-methyl eugenol (34.7%) and (Z)-methyl eugenol (27.5%) [24]. The EO from Piper callosum was also effective against the B. tabaci type B tomato settlement (70.8 % reduction), besides promoting an oviposition deterrent effect (91.5 % reduction). Its main compounds were safrole (29.3%), α-pinene (19.2%), and β-pinene (14.3%) [25].
3.1.7. Poaceae
The study of Ringuelet et al. (2012) demonstrated the effect of Cymbopogon citratus EO 5% on B. tabaci, leading to its repellence (77.32%) and mortality (31.2%) after 24 h with five daily applications. The effect was improved when the laurel EO was mixed with L. nobilis EO at a 1:1 proportion. With this condition, the repellence index enhances to 84.34%, and the mortality rate reaches 55.5% [26].
3.1.8. Rutaceae
Pereira et al. (2018) showed the insecticidal activity of the Zanthoxylum riedelanum fruit EO as well as that of its nanosphere-encapsulated form against B. tabaci. In a free-choice test, the encapsulated and free EO, in a concentration equal to 1.5%, reduced the number of whitefly eggs by approximately 70%. However, in the larvicidal assay, both formulations killed 82.87% and 91.23% of 2nd-instar whitefly nymphs, respectively. Although the encapsulated EO displayed lower insecticidal activity, it offers a greater advantage over the free EO due to the photodegradation protection conferred by the nanosphere polymer. Limonene (29.22%), β-myrcene (22.79%), bicyclogermacene (18.13%), and germacrene D (14.40%) were the major constituents of the EO [27]. More recently, the same research group evaluated the insecticidal activity of the Zanthoxylum rhoifolium fruit EO in its free and encapsulated form in nanospheres. The concentrations in a range between 0.25 and 1.25% reduce adult whitefly oviposition by up to 71%. Also, the formulation containing 1.5% EO caused up to 91% mortality of second-instar nymphs. But when the test was conducted under high temperature and light radiation conditions, the insecticidal effect of the treatments with 1.5% encapsulated EO improved the mortality (84.3%) when compared to the free form (64.8%). The sesquiterpene β-phellandrene (76.8%) was the major compound of the oil [28]. In a previous study, both aforementioned Zanthoxylum EOs were evaluated for their deterrent oviposition activities against B. tabaci biotype B. The best results obtained in the bioassays were achieved at concentrations ranging from 1.0 to 2.0% for both EOs, with a reduced efficiency of egg laying of 85 and 98%, respectively. For these assays, the chemical composition of the oils was γ-elemene (21.2%), germacrene D (14.2%), sabinene (11.9%), and limonene (11.3%) for Z. riedelanium, and sabinene (55.9%), germacrene D (17.1%), and β-myrcene (8.03%) for Z. rhoifolium [29]. Also, the insecticidal and deterrent effects of Z. riedelianum and Z. rhoifolium leaf EOs were evaluated. Bioassays with free and nanoencapsulated EOs of Z. rhoifolium significantly reduced the number of nymphs and eggs of B. tabaci, so that the best results were observed with the free EO at a concentration of 5% in a free-choice test, which causes 100% mortality (nymphs) after 16 days of treatment and 98% (oviposition) reduction after 6 days of treatment. Under the same conditions, the EO from Z. ridelianum 5% presented the best result, causing 100% mortality of nymphs and 97% ovipositon reduction. γ-elemene (24.81%), phytol (18.16%), bicyclogermacrene (16.18%), cis-nerolidol (8.26%), and D-germacrene (6.52%) were the major compounds of Z. riedlianum EO, whereas β-elemene (31.26%), D-germacrene (18.16%), β-caryophyllene (12.09%), δ-elemene (7.63%), β-cedrene (6.69%), bicyclogermacrene (4.57%), and E-caryophyllene (3.63%) were the main substances in Z. rhoifolium EO [30,31].
The insecticidal activity of the genus Citrus on B. tabaci type B was evaluated by Ribeiro et al. (2020). In this study, the fumigant and fecundity bioassays were performed in order to measure the efficiency of peel EOs from four Citrus species after 24 h. Among the evaluated EOs, Citrus aurantiifolia (LC50 = 0.70 μL/L air) and Citrus limon (LC50 = 1.77 μL/L air) presented the best results in the fumigant test. In turn, C. reticulata and C. sinensis × C. reticulata EOs showed LC50 values equal to 3.04 and 5.39 μL/L air, respectively. Regarding the fecundity bioassay, the EO from C. sinensis × C. reticulata inhibits 94.93% of deposited eggs with a concentration of 3 μL/L air. In sequence, the EOs with the best results were: C. limon (86.34%, 0.8 μL/L air), C. aurantiifolia (85.51%, 0.125 μL/L air), and C. reticulata (82.71%, 1 μL/L air). The chemical composition of the EOs demonstrated a higher percentage of limonene in all species (77.79% in C. sinensis × reticulata, 60.96% in C. reticulata, 40.70% in C. limon, and 37.73% in C. aurantiifolia), whereas β-pinene was representative in C. aurantiifolia (9.89%) and C. limon (18.14%), p-Mentha-2,4(8)-diene in C. sinensis × C. reticulata (9.80%), myrcene in C. reticulata (4.61%) and C. sinensis × reticulata (6.50%), and α-terpineol in C. aurantiifolia (5.04%) and C. limon (2.78%) [32]. Ten years before, similar results were obtained with peel EOs from C. sinensis var. pear and C. aurantium. In the fumigant assay, C. sinensis EO was the most active (LC50 = 3.80 μL/L air), followed by C. aurantium EO (LC50 = 5.80 μL/L air). In the oviposition deterrence assay, the C. sinensis EO was also more efficient than C. aurantium, presenting an IC50 = 4 μL/L air against a value equal to 5 μL/L air [33].
4. DISCUSSION
In this review, at least eighteen plant species belonging to eight families were found to have EOs available in countries on the American continent with insecticide activity against B. tabaci [Figures 1 and 2]. The diversity of sources of natural resources is considered of great importance so that the emergence of resistance is minimized, in addition to the fact that the inherent chemical variety can also contribute to the synergistic action of the oils active compounds [Figure 3]. In addition, the diversity of active natural resources prevents only one or a few species from being exhausted to provide products in the fight against a particular pest.
 | Figure 1: Example of plants with high activity EOs against Bemisia tabaci. At the top: Citrus aurantifolia (left), Citrus limon (right). At the bottom: Pelargonium graveolens (left), Piper callosum (right). [Click here to view] |
 | Figure 2: Adults of Bemisia tabaci settled on leaves. [Click here to view] |
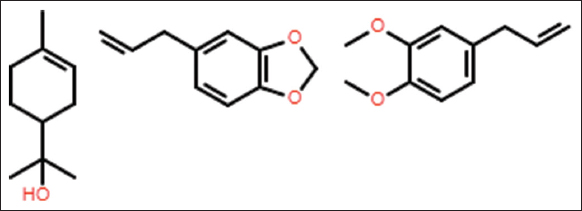 | Figure 3: Important major compounds from EOs with high insecticidal activity against B. tabaci. From left to right: α-terpineol, safrole and methyl eugenol. [Click here to view] |
Among the most studied species are those of the Rutaceae family. In the study by Ribeiro et al. (2020) [32], it was demonstrated that the EOs of Citrus aurantiifolium and C. limon were the most efficient in insecticide and deterrent activities. The same article informs that the two EOs were the richest in α-terpineol, which, in turn, was also tested for insecticidal activity, presenting an LC50 of 1.43 μL/L air, the most active among the majority tested, which suggests the specific participation of this substance in the mortality of B. tabaci, while the deterrent effect seems to have a multiple contribution of active substances. Furthermore, it is already well known that α-terpineol has repellent or insecticidal activity against different species, such as Aedes aegypti, Sitophilus zeamais, and Stomoxys calcitrans, and may interfere with olfactory responses in insect antennae, besides also presenting relevant anticholinesterasic activity [34,35]. As for the species of the genus Zanthoxylum, both were efficient in terms of mortality and oviposition deterrence of the intenses, even though their chemical components varied greatly in the aforementioned studies.
On the other hand, the study by Fanella et al. (2015) [25] demonstrated that the EOs of P. graveolens and P. callosum were more efficient in reducing the settlement of B. tabaci in tomato plants and oviposition. P. graveolens has a high concentration of oxygenated monoterpenes, while P. callosum has safrole as its main substance. Oxygenated monoterpenes (OM) are related to the specificity of the biological activity of EOs, as described by Moghaddam and Mehdizadeh (2017), as they can interact specifically with enzymes [36]. OMs with aromatic functions, such as safrole and methyl eugenol (the main compound in P. marginatum), are also powerful insecticides, which in turn are related to different mechanisms, including octopamine receptors blocking and potentiating ligand-gated chloride channels in the insect nervous system. Furthermore, the interaction with the cholinergic and detoxification systems of insects is also an important consideration [37].
The correlated diallyl trisulfide and diallyl disulfide are characteristic substances of Bigoniaceae species and possess several biological properties, including insecticide activity. However, it is believed that both compounds act antagonistically at the same action sites, connecting to them through bonds with the sulfur atom [38,39]. Then, according to these studies, they are probably the main ones responsible for the activity of the oils extracted from the species M. alliaceae and A. alliaceum.
Furthermore, the study by Ringuelet et al. (2012) [26] demonstrated that the EO from L. nobilis was more efficient than that from Cymbopogon citratus in causing mortality in B. tabaci, while the latter showed slightly more repellence efficacy. In addition, the study also reported that the equal mixture between the two oils increased both activities, contributing synergistically. Still, the EO from X. aromatica also presented interesting deterrent activity, in which the sesquiterpene bicyclogermacrene and the pinene isomers were the major compounds. Bicyclogermacrene has already demonstrated intense insecticidal activity and acts as a hormonal interferent [40]. In turn, pinenes can disrupt the cellular membrane due to their low polarity characteristic, disaggregating the lipid structure that makes up the inner layer of the membrane [41].
In addition, the use of EOs to control B. tabaci can also have a beneficial impact on the economic sector, in order to encourage the cultivation and commercialization of active species, the market for natural products, as well as, possibly in the long term, decrease the cost related to health treatments due to the adverse effects caused by synthetic pesticides.
Finally, the studies discussed here can serve as a basis for the implementation of new products for the control of B. tabaci in countries of the American continent in order to move towards a more sustainable insecticide control, replacing, in part, the conventional pesticides found at the moment [Figure 4].
 | Figure 4: Graphical abstract (scheme). [Click here to view] |
5. CONCLUSION
This review article compiled and discussed the biological activities of American-available EOs against B. tabaci, one of the main agricultural pests worldwide. Among the most studied species were those of the Rutaceae family, including those of the genus Citrus and Zanthoxylum. In addition, species that produce particular groups of insecticidal compounds, as is the case with the Bigoniaceae, also demonstrated relevant activity against the pest. Furthermore, EOs from species that are widespread throughout America, such as Cymbopogum citratus and L. nobilis, and which can be considered abundant resources in the control of B. tabaci, have also been reported. In general, these studies can serve as a basis for the development of specific agricultural products that are more sustainable and safe for consumers.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. FUNDING
There are no fundings for this work.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Kiaya V. Post-harvest Losses and Strategies to Reduce them. Vol. 25. Technical Paper Postharvest Losses;2014. 1-25.
2. World Health Organization. Guidance on Mainstreaming Biodiversity for Nutrition and Health. Geneva:World Health Organization;2020. Available from:https://apps.who.int/iris/bitstream/handle/10665/351047/9789240006690-eng.pdf?sequence=1 [Last accessed on Jun].
3. Oliveira CM, Auad AM, Mendes SM, Frizzas MR. Crop losses and the economic impact of insect pests on Brazilian agriculture. Crop Prot 2014;56:50-4. [CrossRef]
4. Government Accountability Office (GAO). Imported Agriculture:Updated Planning and Communication Could Enhance Agency Coordination of Inspections;2021. Available from:https://www.gao.gov/products/gao-21-471 [Last accessed on Jun].
5. Porter SD, Reay DS, Higgins P, Bomberg E. A half-century of production-phase greenhouse gas emissions from food loss and waste in the global food supply chain. Sci Total Environ 2016;571:721-9. [CrossRef]
6. Savary S, Willocquet L, Pethybridge SJ, Esker P, McRoberts N, Nelson A. The global burden of pathogens and pests on major food crops. Nat Ecol Evol 2019;3:430-9. [CrossRef]
7. Rathee M, Dalal P. Emerging insect pests in Indian agriculture. Indian J Entomol 2018;80:267-81. [CrossRef]
8. Sani I, Ismail SI, Abdullah S, Jalinas J, Jamian S, Saad N. A review of the biology and control of whitefly, Bemisia tabaci (Hemiptera:Aleyrodidae), with special reference to biological control using entomopathogenic fungi. Insects 2020;11:619. [CrossRef]
9. Chandel RS, Banyal DK, Singh BP, Malik K, Lakra BS. Integrated management of whitefly, Bemisia tabaci (Gennadius) and potato apical leaf curl virus in India. Potato Res 2010;53:129-39. [CrossRef]
10. Ellango R, Singh ST, Rana VS, Priya NG, Raina H, Chaubey R, et al. Distribution of Bemisia tabaci genetic groups in India. Environ Entomol 2015;44:1258-64. [CrossRef]
11. Horowitz AR, Ghanim M, Roditakis E, Nauen R, Ishaaya I. Insecticide resistance and its management in Bemisia tabaci species. J Pest Sci 2020;93:893-910. [CrossRef]
12. Vassiliou V, Emmanouilidou M, Perrakis A, Morou E, Vontas J, Tsagkarakou A, et al. Insecticide resistance in Bemisia tabaci from Cyprus. Insect Sci 2011;18:30-9. [CrossRef]
13. Bass C, Field LM. Neonicotinoids. Curr Biol 2018;28:R772-3. [CrossRef]
14. Ongono JS, Beranger R, Baghdadli A, Mortamais M. Pesticides used in Europe and autism spectrum disorder risk:Can novel exposure hypotheses be formulated beyond organophosphates, organochlorines, pyrethroids and carbamates?-a systematic review. Environ Res 2020;187:109646. [CrossRef]
15. Tunaz H, Uygun N. Insect growth regulators for insect pest control. Turk J Agric For 2004;28:377-87.
16. Smith, CJ, Perfetti TA. A comparison of the persistence, toxicity, and exposure to high-volume natural plant-derived and synthetic pesticides. Toxicol Res Appl 2020;4:1-15. [CrossRef]
17. Dubey NK, Shukla R, Kumar A, Singh P, Prakash B. Global scenario on the application of natural products in integrated pest management programmes. In:Natural Products in Plant Pest Management. Wallingford:CABI;2011. 1-20. [CrossRef]
18. Bakkali F, Averbeck S, Averbeck D, Idaomar M. Biological effects of essential oils--a review. Food Chem Toxicol 2008;46:446-75. [CrossRef]
19. Isman MB. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu Rev Entomol 2006;51:45-66. [CrossRef]
20. Pavela R. Essential oils for the development of eco-friendly mosquito larvicides:A review. Ind Crops Prod 2015;111:896-902. [CrossRef]
21. Iqbal T, Ahmed N, Shahjeer K, Ahmed S, Al-Mutairi KA, Khater HF, et al. Botanical insecticides and their potential as anti-insect/pests:Are they successful against insects and pests?In:Global Decline of Insects. London:IntechOpen;2021. 123. [CrossRef]
22. Ogungbe IV, Setzer WN. In-silico Leishmania target selectivity of antiparasitic terpenoids. Molecules 2013;18:7761-847. [CrossRef]
23. Peres MC, de Souza Costa GC, dos Reis LE, da Silva LD, Peixoto MF, Alves CC, et al. In natura and nanoencapsulated essential oils from Xylopia aromatica reduce oviposition of Bemisia tabaci in Phaseolus vulgaris. J Pest Sci 2020;93:807-21. [CrossRef]
24. Santana AS, Baldin EL, Lima AP, dos Santos TL, Santos MC, Vieira TM, et al. New challenges demand new solutions:Selected essential oils as an alternative to control Bemisia tabaci MED in Brazil. Crop Prot 2022;155:105909. [CrossRef]
25. Fanela TL, Baldin EL, Pannuti LE, Cruz PL, Crotti AM, Takeara R, et al. Lethal and inhibitory activities of plant-derived essential oils against Bemisia tabaci Gennadius (Hemiptera:Aleyrodidae) biotype B in tomato. Neotrop Entomol 2016;45:201-10. [CrossRef]
26. Ringuelet JA, Urrutia MI, Yordaz RM, Henning CP. Insecticidal and repellent activity of essential oils of lemongrass and laurel on Bemisia tabaci. Bol Sanid Veg Plagas 2012;38:353-60.
27. Pereira KC, Quintela ED, da Silva DJ, do Nascimento VA, da Rocha DV, Silva JF, et al. Characterization of nanospheres containing Zanthoxylum riedelianum fruit essential oil and their insecticidal and deterrent activities against Bemisia tabaci (Hemiptera:Aleyrodidae). Molecules 2018;23:2052. [CrossRef]
28. Pereira KC, Quintela ED, do Nascimento VA, da Silva DJ, Rocha DV, Silva JF, et al. Characterization of Zanthoxylum rhoifolium (Sapindales:Rutaceae) essential oil nanospheres and insecticidal effects to Bemisia tabaci (Sternorrhyncha:Aleyrodidae). Plants (Basel) 2022;11:1135. [CrossRef]
29. Costa EC, Christofoli M, de Souza Costa GC, Peixoto MF, Fernandes JB, Forim MR, et al. Essential oil repellent action of plants of the genus Zanthoxylum against Bemisia tabaci biotype B (Homoptera:Aleyrodidae). Sci Hortic 2017;226:327-32. [CrossRef]
30. Christofoli M, Costa EC, Peixoto MF, Alves CC, Costa AC, Fernandes JB, et al. Nanoparticles loaded with essential oil from Zanthoxylum riedelianum Engl. Leaves:Characterization and effects on Bemisia tabaci Middle-east Asia minor 1. Neotrop Entomol 2022;51:761-76. [CrossRef]
31. Christofoli M, Costa EC, Bicalho KU, de Cássia Domingues V, Peixoto MF, Alves CC, et al. Insecticidal effect of nanoencapsulated essential oils from Zanthoxylum rhoifolium (Rutaceae) in Bemisia tabaci populations. Ind Crops Prod 2015;70:301-8. [CrossRef]
32. Ribeiro NC, Da Camara CA, De Melo JP, De Moraes MM. Insecticidal potential of Citrus and Mango essential oils and selected constituents on Silverleaf whitefly. Rev Caatinga 2020;33:90-9. [CrossRef]
33. Ribeiro NC, da Camara CA, de Souza Born F, de Siqueira HÁ. Insecticidal activity against Bemisia tabaci biotype B of peel essential oil of Citrus sinensis var. pear and Citrus aurantium cultivated in Northeast Brazil. Nat Prod Commun 2010;5:1819-22. [CrossRef]
34. Khaleel C, Tabanca N, Buchbauer G. α-Terpineol, a natural monoterpene:A review of its biological properties. Open Chem 2018;16:349-61. [CrossRef]
35. Hung NH, Quan PM, Satyal P, Dai DN, Hoa VV, Huy NG, et al. Acetylcholinesterase inhibitory activities of essential oils from Vietnamese traditional medicinal plants. Molecules 2022;27:7092. [CrossRef]
36. Moghaddam M, Mehdizadeh L. Chemistry of essential oils and factors influencing their constituents. In:Soft Chemistry and Food Fermentation. Cambridge:Academic Press;2017. 379-419. [CrossRef]
37. Tabari MA, Rostami A, Khodashenas A, Maggi F, Petrelli R, Giordani C, et al. Acaricidal activity, mode of action, and persistent efficacy of selected essential oils on the poultry red mite (Dermanyssus gallinae). Food Chem Toxicol 2020;138:111207. [CrossRef]
38. Sarma R, Adhikari K, Mahanta S, Khanikor B. Combinations of plant essential oil based terpene compounds as larvicidal and adulticidal agent against Aedes aegypti (Diptera:Culicidae). Sci Rep 2019;9:9471. [CrossRef]
39. Rao PS, Midde MN, Miller DD, Chauhan S, Kumar A, Kumar S. Diallyl sulfide:Potential use in novel therapeutic interventions in alcohol, drugs, and disease mediated cellular toxicity by targeting cytochrome P450 2E1. Curr Drug Metab 2015;16:486-503. [CrossRef]
40. Govindarajan M, Benelli G. Eco-friendly larvicides from Indian plants:Effectiveness of lavandulyl acetate and bicyclogermacrene on malaria, dengue and Japanese encephalitis mosquito vectors. Ecotoxicol Environ Saf 2016;133:395-402. [CrossRef]
41. Salakhutdinov NF, Volcho KP, Yarovaya OI. Monoterpenes as a renewable source of biologically active compounds. Pure Appl Chem 2017;89:1105-17. [CrossRef]