
1. INTRODUCTION
Coccidiosis is a parasitic disease caused by Eimeria species in chickens costing a worldwide annual loss of more than a billion US dollars [1]. Chickens are affected by seven different Eimeria species and amongst them; E. tenella is the most pathogenic [2-4]. The life cycle of Eimeria involves many stages occurring inside and outside the host [5]. This parasite exists in two stages: Sporulated and unsporulated stage. The sporulated stage causes infection in the birds. Dormant Eimeria oocysts undergo a sporulation process to form sporulated oocysts, which are considered pathogenic [6]. The sporulation of the oocysts happens outside the birds and is dependent on temperature, humidity, and access to oxygen [7].
The infection is transmitted to the birds through the ingestion of infective sporulated oocysts that target the gut [1]. After targeting the gut, the intestinal epithelial cells are invaded during the multiplication stage, destroying the intestinal lining [5]. This negatively impacts performance, including increased feed conversion values and reduced body weight gain, leading to mortality [1]. The oocyst cell wall of coccidia is reported to be robust and highly resistant to chemical treatments [8]. Since the infectious nature of the oocyst is decided outside the host, it is possible that by inhibiting the sporulation process, the infectivity of the coccidia could be lessened.
Disinfectants, classified under the biosecurity program, are used to prevent the incidence of disease caused by pathogenic organisms [9]. These products are used to clean and disinfect the farm floor, equipment, cages, and associated accessories. Several studies have reported the efficacy of different classes of disinfectants, reagents, and herbal extracts against Eimeria oocysts [9-11]. In a study reported by Gadelhaq et al., chemicals such as ethanol, formalin, and sodium hypochlorite inhibited the sporulation ranging from 49% to 100% [9]. William et al. showed the sporulation inhibitory effect of phenols, chlorinated phenols, ammonia, and methyl bromide [11]. In another study, chemicals such as acetic acid, cresols, and a combination of benzene and xylene were found to be good inhibitors of E. tenella sporulation [12]. The efficacy of phenolic disinfectant (PD) comprising a mixture of cresylic acid, ortho-benzyl-para-chlorophenol, and ortho-phenyl phenol against sporulation suppression of E. tenella is not known. The objective of the study was to test the efficacy of PD on the inhibition of E. tenella’s sporulation.
2. MATERIALS AND METHODS
2.1. Chemicals and Reagents
Ammonium hydroxide, sodium hydroxide, sodium chloride, and potassium permanganate were purchased from Sigma-Aldrich, India. The PD comprising a mixture of a cresylic acid (30%), ortho-benzyl-para-chlorophenol (3%), and ortho-phenyl phenol (4%) was received from Kemin Industries South Asia Pvt. Ltd., Chennai, India.
2.2. E. tenella Oocysts Collection and Purification
Oocysts of E. tenella were taken from commercial broiler birds affected with clinical coccidiosis. The symptoms of clinical coccidiosis are bloody diarrhea, reduced feed intake, and reduced growth rates, and in severe cases, it leads to mortality [13,14]. Clinical coccidiosis in broiler birds is diagnosed by monitoring the presence of gross lesions in the intestinal tract which includes thickening, hemorrhage, and ulceration [15,16]. Caecal contents were cleaned by flotation method using a saturated sodium chloride salt solution [9]. Briefly, 5 g of caecal contents from the infected birds were collected and mixed with 50 mL of saline [17]. The mixture was vortexed for 1 min and the resulting suspension was filtered using a double-layered muslin cloth and centrifuged at 1000 g for 5 min [17]. After removing the supernatant, the pellet was re-suspended in 5 mL of saturated sodium chloride salt solution [18]. Oocysts float in saturated salt solutions forming an opaque ring on the top, thereby enabling us to collect easily [17,18]. Then, the collected oocysts were pooled and washed 3 times with phosphate-buffered saline using a centrifugation process (1000 g, 15 min, 25°C, Eppendorf, 5810R). The oocysts were counted using a hemocytometer (BS748, Rohem, India) and the morphological characteristics were considered for the confirmation of the oocysts stage [19].
2.3. Treatment Groups
The study consisted of three control groups and two treatment groups. The control groups are control (C), a negative control (NC), and a positive control (PC). The C, NC, and PC groups were treated with saline, sodium hydroxide, and ammonium hydroxide, respectively. The C group is treated with saline, which helps in assessing the viability of the oocysts, during the treatment period. In NC, sodium hydroxide is used, which was known to have no effect on the sporulation of oocysts [19]. In PC, ammonium hydroxide is used and is known to inhibit the sporulation process of oocysts [19]. The treatment groups were designated as T1 and T2, treated with PD at 5 mL/L and 10 mL/L, respectively. After the treatment, the oocysts were subjected to sporulation for 7 days.
2.4. Oocyst treatment
The effect of PD on E. tenella oocysts was studied using unsporulated oocysts, as described by Gadelhaq et al., with modifications [9]. Briefly, the oocysts were normalized to the concentration of 105 cells/mL using saline. To 1 mL of cells, chemicals or PD were added at the concentration mentioned in Table 1 in triplicates and incubated at room temperature for 3 h. The cells were then centrifuged at 8000 g for 15 min at 25°C, and the supernatant was discarded. Then, the oocyst pellet was washed 3 times with 1 mL of saline. After washing, the samples were subjected to sporulation using 1 mL of 2.5% KMnO4 and aerated for 7 days using SeBO® air pump AP-500 (AquaSovi, India). Samples were collected (25 μL) from each group on daily basis and immediately analyzed for the sporulation stage using a hemocytometer and Leica DM-1000 microscope (Leica Microsystems, Germany) at 40X magnifying power. At the end of 7 days, the percentage of cells which were sporulated, sporulating, and unsporulated was calculated based on the initial cell count.
Table 1: Description of the treatment groups and their dosage.
| Treatment groups | Products/reagents | Dose* |
|---|---|---|
| Control | Saline | - |
| Negative control | Sodium hydroxide (10 N) | 0.1 mL/mL |
| Positive control | Ammonium hydroxide (33%) | 0.5 mL/mL |
| T1 | PD | 5 mL/L |
| T2 | PD | 10 mL/L |
* Doses were fixed based on the recommended dose of the product and concentrations used in the reported studies. PD: Phenolic disinfectant
2.5. Inhibitory Activity (IA) Measurement
Based on the sporulation stage of cells calculated above, the IA of the product and chemicals was based on the calculation shown in Equation 1, described by Gadelhaq et al., and Samaha et al [9,20].
 |
2.6. Statistical Analysis
The Statistical Analysis System from STATGRAPHICS® Centurion XVI software, Version 16.2.04 (Statgraphics Technologies, Inc., Virginia, USA) was used for performing the statistical analyses. Data were analyzed with one-way ANOVA at a 95% confidence level and P < 0.05 was considered for determining the significant difference.
3. RESULTS AND DISCUSSION
3.1. Identification of Different Stages of the Oocyst
After treatment with the respective products, the cells were identified based on their morphology, and the results are shown in Figure 1 [19]. A clear differentiation between sporulated, sporulating, and unsporulated stages was observed. Unsporulated oocysts appeared as one whole cell inside the oocyst wall (Figure 1a). The sporulating oocysts were identified with three to four divided cells, overlapping each other inside the oocyst (Figure 1b). The sporulated stage showed well-defined, three to four distinct cells inside the oocyst (Figure 1c). In all the groups, we could identify the various stages of oocysts, which had a clear differentiation of cells within the oocyst (Figure 1).
 | Figure 1: (a-c) Differentiation of sporulation of Eimeria oocysts visualized under a microscope at ×40. Images were captured during sporulation analysis. [Click here to view] |
Kasim et al., also performed similar studies for assessing the effect of Rosmarinus officinalis on the sporulation inhibition of E. tenella [21]. In this report, different stages of oocysts were reported with clear differentiation of cells within the oocysts, as observed in this study.
Li et al., investigated the effects of nitric oxide (NO) donors on the sporogony of E. tenella oocysts and the blocking effects of NO scavenger, hemoglobin (Hb), against NO donors [22]. The study found that acidic sodium nitrite, S-nitrosoglutathione, and S-nitroso-N-acetylpenicillamine were able to completely inhibit the sporulation of the oocysts in a dose-dependent and time-dependent manner. However, in this study, sodium nitroprusside did not demonstrate the desired effects, suggesting the compounds plays a major role in the sporulation suppression.
In general, all freshly shed oocysts consist of a thickened outer well and a rounded mass with a nucleated zygote, and this structure remains intact until the sporulation process occurs [23]. In our study, such morphology of the oocyst was considered as the unsporulated oocyst. In Eimeria species, once the sporulation process occurs, the distinguishing characteristic of each species becomes more apparent. For instance, in the final stages of sporulation, four sporocysts develop within the circumplasm of the oocyst, each containing two banana-shaped sporozoites, and such cells are identified as sporulated oocysts [24]. The sporozoites within the sporulated oocyst may vary in shape and size depending on the species. Before the formation of the sporocysts, the unsporulated oocyst contains a diploid sporont stage which further develops by meiosis, and these oocysts cells are in the sporulating stage, which then divides into four sporocysts [25].
Cysteine proteases (CPs) are enzymes that cleave peptide bonds using a cysteine residue in their active site. They are expressed by many protozoan parasites including E. tenella and are considered to be major virulence factors due to their roles in parasite invasion, replication, and immune evasion [26]. The CP-dependent activities were present throughout the sporulation process suggesting that CPs are active at various stages of the parasite life cycle [27]. This is consistent with the reported studies that CPs as important virulence factors that are involved in many aspects of the parasite life cycle. As various stages of sporulation were seen in this study, it could be possible that the CP was inhibited by the tested compounds leading to sporulation suppression.
3.2. Effect on Sporulation and IA
After treatment of oocysts, the percentage of sporulated, unsporulated, and sporulating oocysts were calculated and the results are shown in Figure 2. It was observed that each treatment group showed varying percentages of oocyst in each of the three stages. More than 65% of oocysts were in a sporulated stage in C and NC, whereas treatment groups showed less percentage of sporulated oocysts. Among the groups, sporulating oocysts were found to be higher with saline and NC. Jitviriyanon et al. also found that the sporulating percentage of oocysts in the saline-treated group was more than 65%, which corroborates the findings observed in this study [28]. The highest percentage (91%) of unsporulated oocyst was observed in the PC group. Chroustova and Pinka also conducted similar kinds of experiments with ammonium hydroxide as one of the test groups and the unsporulated oocysts were found to be >98% [29]. There was a difference in efficacy between this study and reported by Chroustova and Pinka, which could be due to the difference in the number of cells taken for the study [29]. However, >90% of unsporulated oocysts were observed in this study and reported by Chroustova and Pinka [29]. A similar study was also performed with ammonium hydroxide (125 μg/mL) and the unsporulated oocysts were reported to be <5% and the sporulated oocysts were 65% [28]. Such difference in efficacy was inferred to be concentration linked. The concentration of ammonium hydroxide tested by Samaha et al., and by Chroustova and Pinka was at a high concentration (5%), whereas Jitviriyanon et al. tested ammonium hydroxide at 0.0125% [20,28,29]. Such an effect could be resulting from the binding ability between hydroxyl ions and lipids in the cell membrane, the saponification of the lipid leads to the dysfunction of the cell membrane [30]. In NC, the unsporulated oocysts were found to be around 6% indicating sodium hydroxide that did not exhibit any effect on the sporulation process. The results of the sporulation experiments with PC and NC were found to be very closer to the results reported earlier [29]. This validates the sporulation experiments that were carried out for this study.
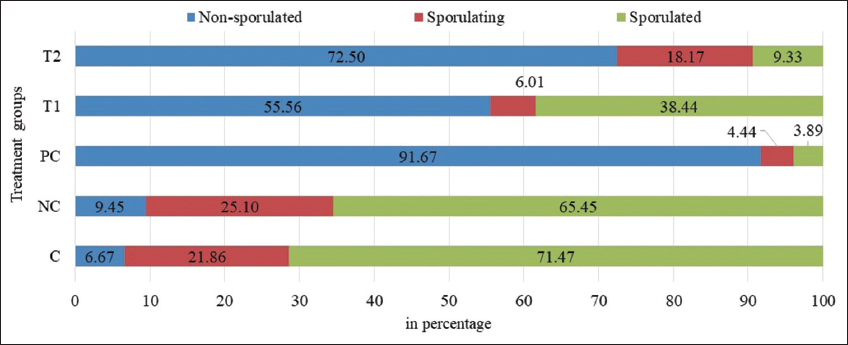 | Figure 2: The percentage of oocysts in the respective sporulation stage at the end of the treatment period. Each data represents the mean of triplicates. C: Saline; NC: Sodium hydroxide; PC: Ammonium hydroxide; PD: Phenolic disinfectant; T1: PD-5 mL/L; T2: PD-10 mL/L. [Click here to view] |
The test groups, T1 and T2, also showed a high percentage of 55% and 71% unsporulated oocysts, respectively. In a study reported by the author, You 30% cresol was taken for the study and the sporulation rate was found to be between 20% and 85% [12]. Higher concentrations showed less sporulation rate indicating the inhibition of the sporulation process and lower concentrations showed a higher sporulation rate suggesting less effect on inhibiting the sporulation process. Since, PD contains a mixture of saponified cresol, ortho–benzyl-para-chlorophenol, and ortho-phenyl phenol, the results observed in this study agree with the results reported by the author, You [12]. Furthermore, a dose-dependent increase in the unsporulated stage was observed between T1 and T2, signifying the impact of PD on the interference of sporulation. In addition, to the high percentage of sporulated oocysts in C and NC, the number of sporulating oocysts was also high and tends to become sporulated oocysts.
In a study conducted by Del Cacho et al., with artemisinin on the sporulation of E. tenella, reduced sporulation rates were observed [1]. Such reduced sporulation rates were inferred to affect the reproduction of this parasite by altering the E. tenella oocyst wall [1]. Fatemi et al. also showed that sporulation of mixed oocysts of Eimeria species was inhibited on treatment with Artemisia annua [31].
The sporulation inhibition effect of Phyllanthus emblica against E. tenella was tested by Sharma et al [32]. In this study, the oocysts were treated with various concentrations of P. emblica, and an almost 35% reduction of sporulated oocysts was observed. Boyko et al., reported the activity of various plant species having essential oils toward sporulation inhibition of Eimeria [33]. The results were essential oil from Syzygium aromaticum exerted a 54% killing effect on the sporulated oocysts and the sporulated oocysts presence was found be zero [33]. These findings suggest that the essential oil exerted its action through the alteration of the cell wall and sporulation process.
The groups were evaluated for IA and the results are shown in Table 2. IA results give the overall view on the efficiency of the reagent or product that was tested in this study, as it considers the sporulating cells tending to become a sporulated stage. It was observed that the highest IA of 91.30% was observed for PC followed by T2 (70.58%), T1 (52.25%), and NC (2.96%), and was found to be statistically different between each of the groups (P < 0.05).
Table 2: Inhibitory activity of the treatment groups.
| Treatment groups | IA (%), (mean±SD, n=3) |
|---|---|
| NC | 2.96±1.44d |
| PC | 91.30±3.04a |
| T1 | 52.25±2.47c |
| T2 | 70.58±2.42b |
Statistical difference between the treatment groups is represented by different alphabets in superscripts (P<0.05). NC: Sodium hydroxide; PC: Ammonium hydroxide; PD: Phenolic disinfectant; T1: PD-5 mL/L; T2: PD-10 mL/L
In an in vitro study conducted by Abouelenien et al., with 10% crude watery extract of Psidium guajava, the IA was found to be 86.4% [34]. Psidium guajava was also tested in the litter, in the manure of birds under field conditions, and reported to decrease the sporulation percent of coccidial oocyst [34]. Such sporulation inhibition resulted in reduced infection rates in the broiler birds.
In another study conducted by Daugschies et al., the efficacy of the cresol-based disinfectant varied between 17% and 49% for various types of coccidia [35]. Cresol derivatives such as chlorocresol were evaluated for their IA by El sherry et al [36]. At 10% and 20%, the IA was found to be 86.7% and 92.7%, respectively. In this study, the PD at 10mL/L (1%) showed IA of 70.58%. Daugschies et al. (2007) showed oocyst destruction after a contact time of 90 min or more with the cresol-based products [35]. Williams indicated that the efficacy of the disinfectant was superior toward unsporulated oocysts compared to sporulated ones [11].
These preliminary results suggest that PD has the potential to prevent coccidiosis incidence in broiler farms.
4. CONCLUSION
Although drugs are available to treat coccidiosis, it has disadvantages such as acquiring resistance, residue concerns in meat, and incompatibility with other chemicals. In broiler farms, the disinfection process is followed regularly, and by choosing the appropriate disinfectant formulation, the coccidiosis incidence in broiler chickens could be prevented. The results that were reported in the study suggest that PD could inhibit sporulation of the oocysts and thereby, coccidiosis can be circumvented to a larger extent. By adopting this practice, the usage of antibiotics in broiler feed can be reduced.
5. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
6. FUNDING
There is no funding to report.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. DATA AVAILABILITY
All the data generated in this study are included in this article.
10. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Del Cacho E, Gallego M, Francesch M, Quilez J, Sanchez-Acedo C. Effect of artemisinin on oocyst wall formation and sporulation during Eimeria tenella infection. Parasitol Int 2010;59:506-11. [CrossRef]
2. Ellakany HF, Abuakkada SS, Oda SS, El-Sayed YS. Influence of low levels of dietary aflatoxins on Eimeria tenella infections in broilers. Trop Anim Health Prod 2011;43:249-57. [CrossRef]
3. Macdonald SE, Nolan MJ, Harman K, Boulton K, Hume DA, Tomley FM, et al. Effects of Eimeria tenella infection on chicken caecal microbiome diversity, exploring variation associated with severity of pathology. PLoS One 2017;12:0184890. [CrossRef]
4. Madlala T, Okpeku M, Adeleke MA. Understanding the interactions between Eimeria infection and gut microbiota, towards the control of chicken coccidiosis:A review. Parasite 2021;28:48. [CrossRef]
5. Lopez-Osorio S, Chaparro-Gutiérrez JJ, Gómez-Osorio, LM. Overview of poultry Eimeria life cycle and host-parasite interactions. Front Vet Sci 2020;7:384. [CrossRef]
6. Lal K, Bromley E, Oakes R, Prieto JH, Sanderson SJ, Kurian D, et al. Proteomic comparison of four Eimeria tenella life-cycle stages:Unsporulated oocyst, sporulated oocyst, sporozoite and second-generation merozoite. Proteomics 2009;9:4566-76. [CrossRef]
7. Rao PV, Raman M, Gomathinayagam S. Sporulation dynamics of poultry Eimeria oocysts in Chennai. J Parasit Dis 2015;39:689-92. [CrossRef]
8. Mai K, Sharman AP, Walker AR, Katrib M, De Souza D, McConville MJ, et al. Oocyst wall formation and composition in coccidian parasites. Mem Inst Oswaldo Cruz 2009;104:281-9. [CrossRef]
9. Gadelhaq SM, Arafa WM, Abolhadid SM. In vitro activity of natural and chemical products on sporulation of Eimeria species oocysts of chickens. Vet Parasitol 2018;251:12-6. [CrossRef]
10. Daugschies A, Bose R, Marx J, Teich K, Friedhoff K. Development and application of a standardized assay for chemical disinfection of Coccidia oocysts. Vet Parasitol 2002;103:299-308. [CrossRef]
11. Williams RB. Laboratory tests of phenolic disinfectants as oocysticides against the chicken coccidium Eimeria tenella. Vet Rec 1997;141:447-8. [CrossRef]
12. You MJ. Suppression of Eimeria tenella sporulation by disinfectants. Korean J Parasitol 2014;52:435-8. [CrossRef]
13. Abbas A, Iqbal Z, Abbas RZ, Khan MK, Khan JA, Mahmood MS, et al. In vivo anticoccidial effects of Beta vulgaris (sugar beet) in broiler chickens. Microb Pathog 2017;111:139-44. [CrossRef]
14. Blake DP. Eimeria genomics:Where are we now and where are we going?Vet Parasitol 2015;212:68-74. [CrossRef]
15. Fatoba AJ, Adeleke MA. Diagnosis and control of chicken coccidiosis:A recent update. J Parasit Dis 2018;42:483-93. [CrossRef]
16. Mesa-Pineda C, Navarro-Ruíz JL, López-Osorio S, Chaparro-Gutiérrez JJ, Gómez-Osorio LM. Chicken coccidiosis:From the parasite lifecycle to control of the disease. Front Vet Sci 2021;8:7?3. [CrossRef]
17. Felici M, Tugnoli B, Piva A, Grilli E. In vitro assessment of anticoccidials:Methods and molecules. Animals (Basel) 2021;11:1962. [CrossRef]
18. Molan AL, Liu Z, De S. Effect of pine bark (Pinus radiata) extracts on sporulation of coccidian oocysts. Folia Parasitol (Praha) 2009;56:1-5. [CrossRef]
19. Desalegn AY, Ahmed MR. Anticoccidial activity of Aloe debrana and Aloe pulcherrima leaf gel against Eimeria oocysts. J Parasitol Res 2020;2020:8524973. [CrossRef]
20. Samaha HA, Haggag YN, Nossair MA, Habib HM. Assessment efficiency of some chemical disinfectants commonly used against Coccidia in poultry farms. Alex J Vet Sci 2013;39:82-90.
21. Kasem SM, Mira NM, Mahfouz ME, Helal IB. In vitro study to evaluate the efficacy of ultrasonicated ethanolic extract of Rosmarinus officinalis and its chitosan-based nanoparticles against Eimeria tenella oocysts of chickens. AAPS PharmSciTech 2022;23:295. [CrossRef]
22. Li JG, Liu ZP, Tao JP. The effects of nitric oxide donors on the sporulation of Eimeria tenella oocysts. Vet Parasitol 2008;154:336-40. [CrossRef]
23. Muller J, Hemphill A. In vitro culture systems for the study of apicomplexan parasites in farm animals. Int J Parasitol 2013;43:115-24. [CrossRef]
24. Deplazes P, Eckert J, Mathis A, Samson-Himmelstjerna GV, Zahner H. Parasitology in Veterinary Medicine. Netherland:Wageningen Academic Publishers;2016. [CrossRef]
25. Chartier C, Paraud C. Coccidiosis due to Eimeria in sheep and goats, a review. Small Rumin Res 2012;103:84-92. [CrossRef]
26. Mendes TA, Lobo FP, Rodrigues TS, Rodrigues-Luiz GF, daRocha WD, Fujiwara RT, et al. Repeat-enriched proteins are related to host cell invasion and immune evasion in parasitic protozoa. Mol Biol Evol 2013;30:951-63. [CrossRef]
27. Rieux A, Gras S, Lecaille F, Niepceron A, Katrib M, Smith NC, et al. Eimeripain, a cathepsin B-like cysteine protease, expressed throughout sporulation of the apicomplexan parasite Eimeria tenella. PLoS One 2012;7:e31914. [CrossRef]
28. Jitviriyanon S, Phanthong P, Lomarat P, Bunyapraphatsara N, Porntrakulpipat S, Paraksa N. In vitro study of anti-coccidial activity of essential oils from indigenous plants against Eimeria tenella. Vet Parasitol 2016;228:96-102. [CrossRef]
29. Chroustova E, Pinka K. The efficiency of disinfectants on the oocysts of Eimeria tenella. Acta Vet Brno 1987;56:141-9. [CrossRef]
30. Maris P. Modes of action of disinfectants. Rev Sci Tech 1995;14:47-55. [CrossRef]
31. Fatemi A, Razavi SM, Asasi K, Goudarzi MT. Effects of Artemisia annua extracts on sporulation of Eimeria oocysts. Parasitol Res 2015;114:1207-11. [CrossRef]
32. Sharma UN, Fernando DD, Wijesundara KK, Manawadu A, Pathirana I, Rajapakse RP. Anticoccidial effects of Phyllanthus emblica (Indian gooseberry) extracts:Potential for controlling avian coccidiosis. Vet. Parasitol Reg Stud Reports 2021;25:100592. [CrossRef]
33. Boyko O, Shendryk L, Shaban O, Brygadyrenko V. Influence of essential oils on sporulation of Eimeria magna oocysts. Ann Parasitol2021;67:11-7.
34. Abouelenien FA, Elshahawy IS, Hamad ME, Elsaidy NR. Biosecurity assessment in relation to the occurrence of some coccidian parasites in poultry farms, with in vitro evaluation of Psidium gujava as Coccidia sporulation inhibitor. Emir J Food Agric 2021;33:532-43. [CrossRef]
35. Daugschies A, Agneessens J, Goossens L, Mengel H, Veys P. The effect of a metaphylactic treatment with diclazuril (Vecoxan) on the oocyst excretion and growth performance of calves exposed to a natural Eimeria infection. Vet Parasitol 2007;149:199-206. [CrossRef]
36. El-sherry SH, Aly MA, Soliman MA, Darwish M, Amen O. Efficiency of four disinfectants against Eimeria tenella isolates from Egyptian chickens (in vitro assessment). Assiut Vet Med J 2021;67:91-100. [CrossRef]