
1. INTRODUCTION
Plant adaptation to critical environmental conditions depends on the most significant temperature and length of exposure, the sort of plant, and other ecological elements [1]. Botanists interested in temperature stress seek to find the plant reactions to warm resilience and explore how plants manage in high-temperature environments [2].The temperature is constantly increasing in Iraq and the world, where it is expected that the temperature will increase by 5°C in the year 2100 and that the transfer of temperature to and from the plant depends on the difference between the temperatures of the plant and the environment in which it is developing, as the exposure of the plant to a high temperature leads to drought and deterioration of growth the plant [3]. The stress resulting from high temperature causes a reduction in photosynthesis and most of the physiological activities in the plant. And effect on cell membranes and proteins, as some resistant plants produce a group of heat shock proteins (HSPs) that contribute to protecting the planet from the effect of high temperatures, which include small molecular weights [2].
The availability of excellent and vigorous seedlings is one of the most important means of spreading and developing multipurpose plants, including Indian almonds [4]. The Indian almond (Terminalia catappa L.) is a perennial tree grown as an ornamental plant in gardens and parks. It is also used for fixing dunes and planting forests. Mature trees are often used for decoration for the beauty of their appearance, as they have straight trunks topped with horizontal branches and dark green leaves. The flowers are oval, grow at the branches’ end, and are greenish-white [5]. The Indian almond requires adequate care, especially during the summer period. They are deciduous plants that need low temperatures during the winter and moderate temperatures for the spring and summer [6]. Adapting plants to external conditions provide additional factors that improve plant tolerance to unfavorable environmental conditions and enhance biochemical processes [7].
The plant management process requires the development of mechanisms and technologies based on bearing environmental stress and limiting its adverse effects on agriculture and production [1]. Therefore, it is necessary to use some mechanisms that increase the heat tolerance of the Indian almond plant, especially in the Middle East, which is witnessing increases in temperature during spring and summer. Among these mechanisms or technologies is the exogenous application of some osmotic-modifying compounds, such as calcium, or antioxidants, such as ascorbic acid (AsA). AsA plays an essential role in plant advancement, signaling of hormones, the cycle of cells, and the redox system of the cell. The AsA content in plant chloroplasts indicates its essential role in the photosynthetic system [8]. Ascorbate is a significant plant metabolite that protects plants from environmental stresses such as salinity and high temperature [9]. AsA is an indicator compound in many studies related to stress relief and is considered one of the most effective growth regulators in different environmental stresses [10].
Calcium regulates the plant’s response to environmental stresses, such as heat stress [11]. Heat stress increases cellular calcium ion content, reducing the harmful effects of high temperatures and enabling plant cells to survive better [12]. Exogenous calcium treatment has been displayed to further develop heat resistance in many plants, which may be related to increased activities of antioxidant enzymes and reduced lipid peroxidation in cell membranes [13]. Calcium use preserves chlorophyll under heat stress, possibly by decreasing photo-oxidation or keeping up with the trustworthiness of cell membranes [14]. A change in the extreme temperatures in recent years exceeded 50°C, which negatively affected the growth of plants and led to their death, especially in the seedling stage. Note that the importance of the Indian almond plant lies in the diversity of its uses for medicinal purposes and as an ornamental plant, in addition to its use as windbreaks and the possibility of obtaining seeds that are used as a primary method for its propagation, which helps to spread it and obtain a profitable economic return as a result of using the currently imported seeds for breeding in local nurseries [15]. The current research aims to reduce the adverse impact of heat stress by treating the seedlings with different calcium chloride and AsA concentrations.
2. MATERIALS AND METHODS
Seeds of Indian almonds (Terminalia catappa L.) were obtained from Aljouri Agricultural materials/(UAE). The seeds were grown in 20 × 25 cm pots in a local nursery for 4 months. On September 1, 2020, 54 seedlings were transferred to plant at the wooden canopy of the Department of Horticulture and Landscaping of the College of Agriculture at the University of Basrah (30° 33’ 47.3” N 47° 44’ 38.4” E). Then, large plastic pots (30 cm × 32 cm) were prepared, washed well with water and sterilized with formalin, and then filled with sterilized growth medium by autoclaving; at a rate of one plant per pot and in soil consisting of (1:1) Bet moss: soil. The electrical conductivity (EC) of soil was 2 dS/m and to the water of irrigation is EC 1.2 dS/m. The process of servicing the plants was carried out symmetrically from fertilization and irrigation, and the plants were approximately 30–40 cm in height. All the experimental plants were also fertilized with the compound fertilizer of Jordanian origin (20-20-20) NPK on February 1, 2021. The fertilizer was repeated throughout the experiment at 1 g/pot month (Fertigation). The pots were taken from the wooden canopy and put in the field on February 10, 2021. On March 9, 2021, the treatments were conducted monthly and over 4 months as a foliar spray until May 9, 2021. Tween 20 was added at 0.1 ml/L as a diffuser to reduce the surface tension of the leaves. It was sprayed in the early morning using polyethylene bags to prevent contamination from the effect of the treatments. The following nine treatments were applied: (1) Control (spray with distilled Water), (2) AsA 50 mg/L, (3) AsA 100 mg/L, (4) CaCl2 250 mg/L, (5) CaCl2 500 mg/L, (6) AsA 50 mg/L + CaCl2 250 mg/L, (7) AsA 50 mg/L + CaCl2 500 mg/L, (8) AsA 100 mg/L + CaCl2 250 mg/L, and (9) AsA 100 mg/L + CaCl2 500 mg/L. On day 60th of treatment, the leaves were collected for morphological and biochemical assay from every treatment. In July 2021, the minimum temperature was 28°C, the maximum was 46°C, the relative humidity was 27%, and the light intensity was 1750 mmol/m2s.
2.1. Growth Parameters
Measurement of the height of the plants was from the soil surface to the end of the growing top using a tape measure. The number of lateral branches of each plant was calculated, and the means were recorded. The number of leaves on the main stem and the lateral branches was calculated, and the average was taken and multiplied by the number of branches to get the total number of leaves of the plant. The total leaf area was measured using the ImageJ program according to Easlon and Bloom [16], and after taking ten leaves for each replicate, they were placed in a scanner. The readings representing the leaf area of the plant were taken according to the following equation:
The leaf area of a plant (cm2) = Leaf area (cm2) × the number of leaves
2.2. Assessment of Photosynthetic Pigments
The substance of pigments was taken out from the leaves as per the method depicted by Lichtenthaler and Wellburn [17]. Preliminary 0.2 g of the sample leaf were crushed in 10 mL acetone (80%) and centrifuged at 2000 rpm for 5 min. The absorbance of 645, 663, 534, and 470 nm was used to pick the chlorophyll and carotenoids freely.
2.3. Soluble Carbohydrate Estimation
The carbohydrate was assessed by the phenol-sulfuric acid technique, as suggested by Yemm and Willis [18]. 100 mg of fresh leaf sample was homogenized in 5 mL of 2.5 N HCl and put in a boiling water bath for 3 h. The cooled rough homogenate was balanced and centrifuged at 10,000 rpm for 10 min. To 100 μL of supernatant, 100 μL phenol (5% [v/v]), and 500 μL of sulfuric acid (96% [v/v]) were mixed. The absorbance was estimated at 490 nm. Glucose going from 0 to 500 μg/mL was utilized as a standard to calculate the soluble carbohydrate of the sample.
2.4. Proline Estimation
The proline content was assessed by Bates et al. [19]. 0.5 g of the test was homogenized with 5 mL of 3% sulfosalicylic acid. The sample was separated, and 3 mL of this separate was mixed with ninhydrin reagent and glacial acetic acid, 3 mL each. This combination was warmed in a bubbling water bath for an hour and cooled. A chromophore was shaped by adding 4 mL toluene to this cooling solution. The absorbance was measured at 520 nm utilizing the ultraviolet–visible (UV-VIS) spectrophotometer. Proline going from 0 to 10 μg/mL was utilized as standard, and a diagram was plotted from which the proline content of the sample was assessed.
2.5. Total Phenolic Assay
The phenols extract was assessed per the Folin-Ciocalteu technique [20]. A 25 μL of the concentrate (500 μg/mL) was utilized, to which 25 μL of (1:1) Folin-Ciocalteu reagent and 100 μL of 7.5% sodium bicarbonate solution were the arrangement and hatched at room temperature for 2 h in dim circumstances 765 nm using a UV-VIS spectrophotometer to record the absorbance. Gallic acid going from 0 to 100 μg/mL was utilized as a standard to calculate the phenol content of the sample.
2.6. AsA Content
AsA was measured by Luwe et al. [21] procedure. Tests (0.5 g) of leaf were homogenized with 10 mL of 6% trichloroacetic acid. The concentrate was blended with 2 mL of 2% dinitrophenylhydrazine (pH 5) trailed using one drop of 10% thiourea (in 70% ethanol). The blend was bubbled for 15 min in a water bath, and after cooling at room temperature, 5 mL of 80% (v/v) H2SO4 was mixed into the mixture at 0°C. 265 nm was used to record the absorbance. The AsA content was determined from a standard bend plotted with its known focus.
2.7. Abscisic Acid (ABA) Analysis
One gram of new leaf tissue was homogenized in 70% methanol at 4°C; it was mixed for the time being. Under a vacuum, the concentrate was separated and dissipated with Whatman channel paper (No.1). The pH of the aqueous stage was changed following 8, utilizing a 0.2 mol phosphate buffer. The aqueous stage was divided twice utilizing methanol. A rotary evaporator eliminates the methanol stage. pH edited to 2.5, utilizing 1N HCl through the aqueous stage. The concentrate injection specified ABA into a turned around stage high-performance liquid chromatography, C12 column in an isocratic elution mode using a convenient stage including (CH3)2CO: H2O (26:74) with 30 mmol phosphoric acids according to Tang et al. [22]. 1 N sodium hydroxide using to keep up pH at 4. The transition rate was 0.6 mL/min, and the elution of ABA was seen at 270 nm at 25ºC.
2.8. Electrolyte Leakage
Leaf segments one gram utilized with 12 mL separated water was saved at 27ºC overnight alongside vibration. The first conductivity (C1) was determined after getting a thermometer test of 27ºC along with a conductivity meter. The models get under autoclaved for 15 min and cooled to 26ºC; by then, the second conductivity (C2) was determined. The film unfaltering quality rundown was once assessed to determine the estimation into C1 and C2 and imparted among the ratio [23].
2.9. Minerals Concentration
Fresh leaf samples were dried at 70°C until arriving at a consistent weight, and the Cresser and Parsons [24] method was applied. The arrangement was straight forward to control the calcium by atomic absorption spectrophotometer at 422.7 nm [25].
2.10. Extraction of Proteins and Gel Electrophoresis
Proteins were separated (300 mg of the dried leaf) in 1 mL of extraction buffer (0.2 mol, tris-hydroxymethyl aminomethane [Tris] + 0.001 mol ethylenediamine tetraacetic acid + [Na2 + ethylenediaminetetraacetic acid] + 12% glycerol + 0.01 M dithiothreitol [DTT] + 0.05 mmol phenylmethylsulfonyl fluoride [PMSF]) a mortar and pestle use to homogenized. By then, the examples were centrifuged at 14,000 × g for 20 min; the buffer comprised of 0.125 M Tris HCl (pH = 6.8) + 4% SDS + 20%, glycerol + 10% b-mercaptoethanol + 0.01% bromophenol blue. Protein tests were denaturized by heat in the water bath at 90°C for 3 min. Protein electrophoresis was acted in SDS polyacrylamide gel following the methodology [26]. The protein groups exhibited clear changes, which were researched by ImageJ programming.
2.11. Response to Heat Stress
Percentage of plants responding to heat stress (%) = Number of plants responding to stress/Total number of plants
2.12. Statistical Analysis
The experiment was conducted as a randomized block complete design of nine treatments. Each one consisted of four replications. The information was exposed to the examination of difference (analysis of variance) utilizing SPSS variation 20.0 (SPSS, Chicago, IL), and the means were isolated utilizing the Duncan test at the 0.05 level.
3. RESULTS
3.1. Growth Parameters
Significant changes in growth parameters in Indian almond seedlings subjected to AsA and CaCl2 alone or combined under high field temperature are shown in Table 1. Applying Ca2+ and AsA individually and in combination improved the height plant, branch number, leaf number, and leaf area compared with the control [Table 1]. The combined application of AsA + Ca2 or individual showed a more enhancing effect on height plant than other treatments. However, all treatments enhancing branches number except control, CaCl2 500, and AsA 100 + CaCl2 500. Whereas treatment of CaCl2 250 increased leaves number and leaf area relative to other treatments.
Table 1: Exogenous AsA and Cacl2 alone or combined on plant height, number of branches, number of leaves, and leaf area in Indian almond seedlings under high field temperature.
| Treatments (mg/L) | Plant height (cm) | Number of branches | Number of leaves (Leaf/plant) | Leaf area (cm2/plant) |
|---|---|---|---|---|
| Control | 63.25±3.30d | 2.50±0.57b | 167.50±35.64cd | 168.50±35.85c |
| AsA 50 | 94.25±4.19a | 5.00±0.05a | 191.25±61.69c | 256.46±82.73c |
| AsA 100 | 92.50±2.38ab | 4.50±1.00a | 299.50±67.95b | 454.34±103.08b |
| Cacl2 250 | 81.75±7.50c | 5.00±0.00a | 381.25±22.50a | 903.18±53.30a |
| Cacl2 500 | 93.75±3.59a | 3.25±0.50b | 121.50±31.89cd | 287.83±75.54c |
| AsA 50+Cacl2 250 | 86.25±4.42bc | 4.50±0.57a | 360.50±26.85ab | 834.91±62.18a |
| AsA 50+Cacl2 500 | 69.25±2.75d | 4.50±1.00a | 388.25±86.96a | 787.75±176.44a |
| AsA 100+Cacl2 250 | 84.50±5.97c | 4.50±0.57a | 350.50±51.57ab | 872.39±128.36a |
| AsA 100+Cacl2 500 | 99.75±5.56a | 3.00±0.50b | 105.75±6.65d | 277.06±17.42c |
Means of 4 replications±SD. Using Duncan’s multiple range test, means with different letters are different at P≤0.05.
3.2. Photosynthetic Pigments
The application of the treatments showed a varying increase in the content of photosynthetic pigments. The combined treatments, especially those of AsA 100 + CaCl2 250 and AsA 100 + CaCl2 500, led to a significant increase in chlorophyll content [Figure 1]. Moreover, applying the combined treatments, the total chlorophyll was increased compared to the individual treatments. The highest total chlorophyll content was applied by treating AsA 100 + CaCl2 500. The combined treatment AsA 100 + CaCl2 500 and the individual treatment CaCl2 500 led to a significant increase in carotenoid pigment compared to the other treatments.
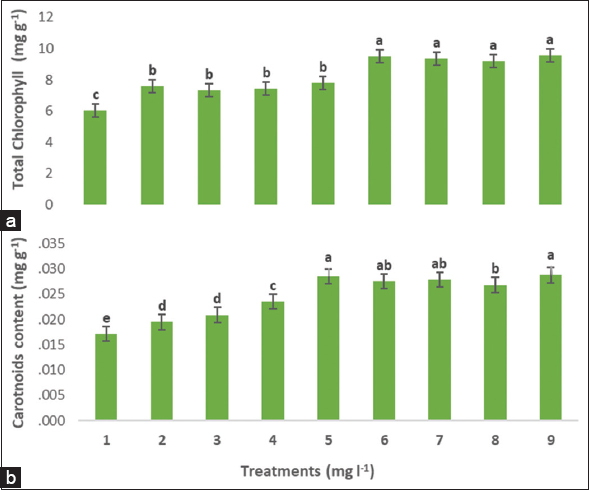 | Figure 1: Exogenous AsA and Cacl2 alone or combined on total chlorophyll (a), and carotenoid (b) content in Indian almond seedlings under high field temperature in July. Treatment numbers are described as (1) Control, (2) AsA 50, (3) AsA 100, (4) Cacl2 250, (5) Cacl2 500, (6) AsA 50 + Cacl2 250, (7) AsA 50 + Cacl2 500, (8) AsA 100 + Cacl2 250, (9) AsA 100 + Cacl2 500. According to the Duncan’s multiple range test, values within the same column show significant differences at P ≤ 0.05 significance level among treatments. Data are means of four replicates ± standard deviation. [Click here to view] |
3.3. Metabolic Compounds
There was a significant increase in proline content under heat stress, whereas total soluble carbohydrate, phenolic, and AsA contents decreased [Figure 2]. The combined AsA + Ca2 or individually modulated proline, total soluble carbohydrate, phenolic, and AsA contents. Moreover, AsA 100 + CaCl2 500 increased total soluble carbohydrate, phenolic, and AsA contents significantly compared with other treatments. Application of AsA 50 decreased proline content significantly compared with other treatments. Application of AsA 100 + CaCl2 250, AsA 100 + CaCl2 500, and AsA 100 increased AsA significantly relative to other treatments.
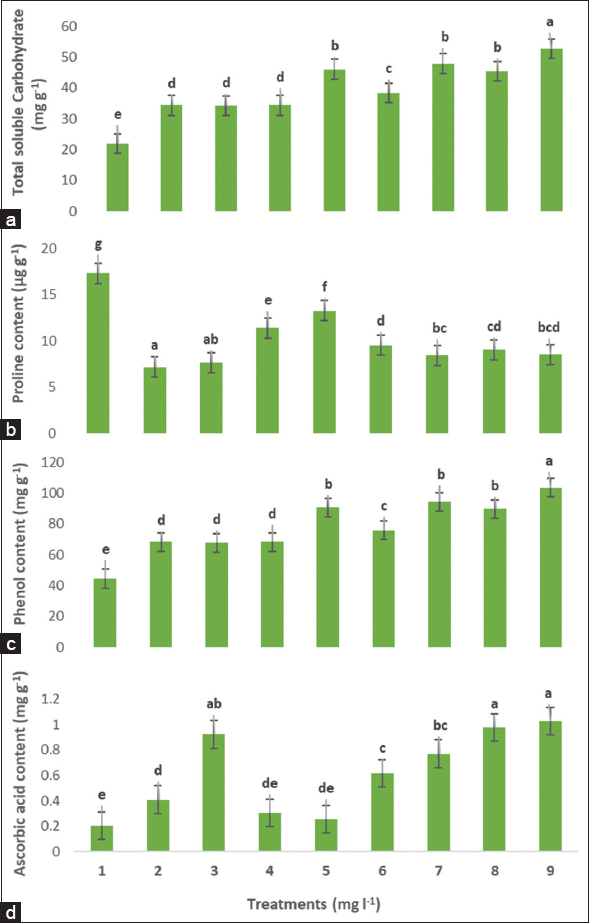 | Figure 2: Exogenous AsA and Cacl2 alone or combined on total soluble carbohydrate (a), proline content (b), phenol content (c), and ascorbic acid (d) content in Indian almond seedlings under high field temperature in July. Treatment numbers are described as (1) Control, (2) AsA 50, (3) AsA 100, (4) Cacl2 250, (5) Cacl2 500, (6) AsA 50 + Cacl2 250, (7) AsA 50 + Cacl2 500, (8) AsA 100 + Cacl2 250, (9) AsA 100 + Cacl2 500. According to the Duncan’s multiple range test, values within the same column show significant differences at P ≤ 0.05 significance level among treatments. Data are means of four replicates ± SD. [Click here to view] |
Heat stress increased ABA content and electrolyte leakage percentage, whereas calcium decreased [Figure 3]. The combined application of AsA + Ca2 or individual decreased ABA content and electrolyte leakage. Moreover, the application of AsA 50 decreased ABA content significantly compared with other treatments. Application of AsA 50, AsA 100, and AsA 100 + CaCl2 500 decreased electrolyte leakage significantly relative to other treatments. Application of CaCl2 250, AsA 50 + CaCl2 500, and AsA 100 + CaCl2 250 increased calcium content significantly compared with other treatments.
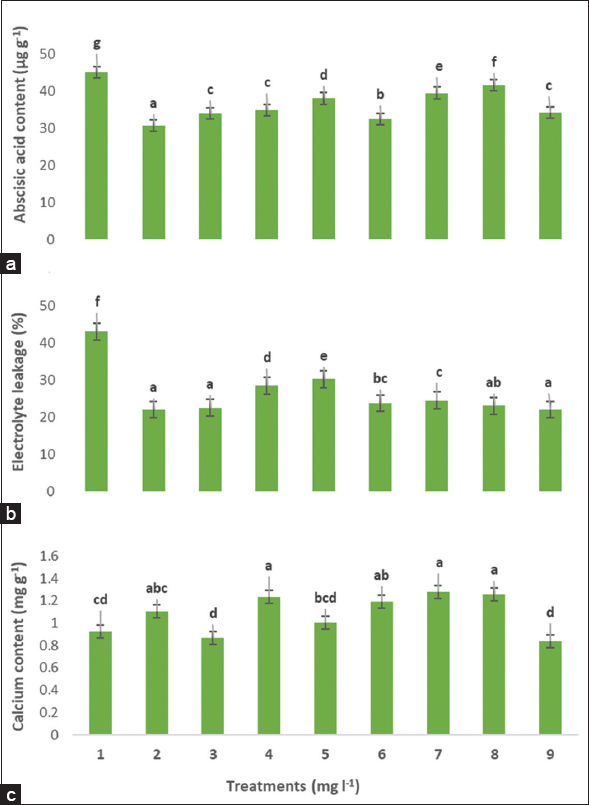 | Figure 3: Exogenous AsA and Cacl2 alone or combined on total abscisic acid content (a), electrolyte leakage (%) (b), and calcium content (c) in Indian almond seedlings under high field temperature in July. Treatment numbers are described as (1) Control, (2) AsA 50, (3) AsA 100, (4) Cacl2 250, (5) Cacl2 500, (6) AsA 50 + Cacl2 250, (7) AsA 50 + Cacl2 500, (8) AsA 100 + Cacl2 250, (9) AsA 100 + Cacl2 500. According to the Duncan’s multiple range test, values within the same column show significant differences at P ≤ 0.05 significance level among treatments. Data are means of four replicates ± SD. [Click here to view] |
3.4. Change in Protein Pattern
The gel electrophoresis of the leaves of Terminalia catappa L. seedlings due to spraying with AsA and calcium [Figure 4]. Differences in the number of proteins separated into bands on the gel and the thickness and density of these bands indicate differences in the molecular weights of proteins. It is clear from the figure that the control treatment gave eight protein bands with molecular weights that included (286.786, 145.178, 111.787, 75.954, 61.375, 45.616, 26.602, and 19.816) kDa. The treatment of AsA 50 mg/L showed ten bands with molecular weights amounted to (200.786, 147,412, 105.394, 75.954, 68.935, 62.595, 54.061, 37.477, 18.792, and 17.496) kDa, which did not differ from the molecular weights of the treatment AsA 100 mg/L that had ten bands, except in two of the bands, had molecular weights (62.595, 27.597) kDa. The treatments of CaCl2 250 and CaCl2 500 showed eight similar bands with molecular weights (122.554, 104.070, 75.954, 66.797, 58.409, 44.828, 28.958, and 19.183) kDa, and only two is different (29.303 and 20.195) kDa in treatment of Cacl2 500. The treatment of AsA 50 + CaCl2 250 and AsA 50 + CaCl2 500 have eleven bands with molecular weights (200.786, 147,412, 105.394, 75.954, 68.585, 62.595, 54.061, 37.477, 27.597, 18.792, and 17.496) kDa except in two of the bands, had molecular weights (37.477 and 19.481) kDa in treatment of AsA 50 + CaCl2 500. The treatment of AsA 100 + CaCl2 250 showed eleven bands with molecular weights amounted to (200.786, 147,412, 105.394, 75.954, 68.935, 62.595, 54.061, 36.825, 27.597, 18.792, and 17.496) kDa. At the same time, the treatment of AsA 100 + CaCl2 500 showed eleven bands gave molecular weights identical to the previous treatment except for two bands whose molecular weight reached (152.045 and 37.477) kDa.
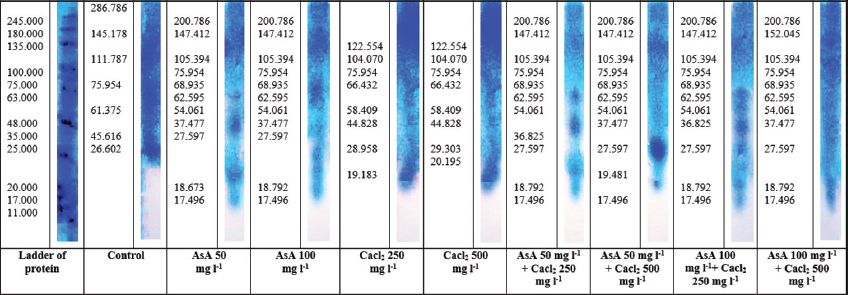 | Figure 4: Analysis of protein patterns by One-D SDS-PAGE extracted from the leaf, showing protein pattern changes in Indian almond seedlings under field high temperature and treatments. [Click here to view] |
3.5. Response to Thermal Stress
The effect of AsA and calcium on the response rate to thermal stress during the 2nd week of July [Figure 5]. The results showed that with the increase in the average temperature during the 2nd week to 40.28°C (49/31°C), the seedlings responded to heat stress and that the control treatment was more responsive to the temperature rise than the other treatments. The response rate to thermal stress in control reached 50%, followed by treatment of AsA 100 + CaCl2 250 with a response rate of 33.33%. In contrast, the treatments of Cacl2 250, AsA 50 + CaCl2 250, AsA 50 + CaCl2 500, and AsA 100 + CaCl2 500 showed a response rate of 16.66%. In contrast, the other treatments did not respond to thermal stress this week. The data is not shown; the high and low temperatures varied, and when the temperatures reached the average of 34°C (42/26°C) in mid-September, all seedlings recovered their growth.
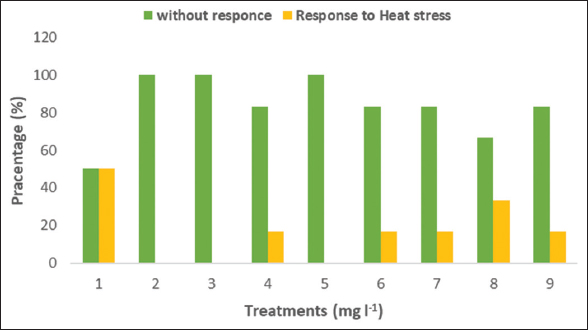 | Figure 5: Exogenous AsA and Cacl2 alone or combined in response to heat stress in Indian almond seedlings under high field temperature in July. Treatment numbers are described as (1) Control, (2) AsA 50, (3) AsA 100, (4) Cacl2 250, (5) Cacl2 500, (6) AsA 50 + Cacl2 250, (7) AsA 50 + Cacl2 500, (8) AsA 100 + Cacl2 250, 9) AsA 100 + Cacl2 500. [Click here to view] |
4. DISCUSSION
In this study, a tropical plant of Indian almond was grown in a subtropical environment. Subtropical regions suffer from high temperatures and little or no rain in the summer. Biochemical reactions within plants are sensitive to high-temperature stress, which vary with temperature, duration of exposure, and plant type [27]. High temperatures cause cell damage or plant death within minutes, and moderate temperatures may cause cell injury or death with prolonged exposure [28]. High temperature impairs plant growth stages and physiological processes and significantly reduces the productivity of many plant species [29]. High-temperature formats the harmful substances in plants due to disruption in the processes of photosynthesis and the ability to respiration in plants, subsequently, a change in the color of leaves to yellow, inhibiting growth processes in plants and destroying chlorophyll [30]. Water evaporates greatly from plants, causing them to dry out [Figure 1a]. Higher temperatures in the summer cause a decrease in growth parameters, photosynthetic pigments, and metabolic compounds, whereas proline and ABA increase [Figures 1-3]. The high temperatures increase the activity of the enzyme chlorophyllase, which destroys chlorophyll and increases the level of ABA, accelerating the decomposition of chlorophyll [30].
Carotenoids perform two significant roles in the photosynthetic plant system. The first role is to act as light-harvesting pigments, expanding the spectrum of light available for use in photosynthesis. It absorbs light at 450–570 nm, which does not absorb chlorophyll molecules, and passes the captured energy onto chlorophyll. Second, carotenoids provide photosynthetic systems with photoprotective methods. Single oxygen (O-2) was detected in the chloroplasts of hydrolyzed wheat [31]. O-2 is a potent oxidizer, strong enough to cause the death of the plant. Carotenoids inhibit the formation of O-2 by quenching the triplet state of chlorophyll molecules when they appear [31].
The effect of heat stress was reversed due to the foliar spray with antioxidants, as AsA is one of the non-enzymatic antioxidants that remove reactive oxygen radicals (ROS), which reduces the breakdown of chlorophyll pigment under stress conditions [32,33]. The role of AsA in increasing plant growth is attributed to increasing the size of cells and the speed of their division, as well as increasing the process of photosynthesis and carbohydrate metabolism [34], and improving the water content of the plant, and reducing transpiration [35]. The effect of AsA on growth parameters agrees with many studies that showed that AsA has a role in encouraging growth and reducing the harmful effect of environmental stresses, as found in Faisal et al. [36] on broad beans; Ullah et al. [37] on barley plants.
Bhattacharya et al. [38] indicated that the increase in antioxidants and the decrease in hydrogen peroxide (H2O2) delay leaf aging. The antioxidants protect the chloroplast, preventing the breakdown of chlorophyll by scavenging ROS. When treated with calcium, the increase in leaf area is one of the components of cellular membranes and its increased stability when exposed to external stresses–cells in carrier cells in phloem under stress conditions [39]. The protective role of calcium is attributed to maintaining the structural integrity of the plasma membrane and thus controlling ion uptake. However, the increased growth is due to cell expansion and photosynthesis activation by increasing membrane stability [Figure 3b].
The decrease in total soluble carbohydrates in the control treatment due to the effects of high temperature and exposure of seedlings to thermal quiescence programming is represented by a decrease in the leaf area and the total chlorophyll content [Figure 1b]. The effect on the activity of the enzymes responsible for carbon dioxide reduction, especially the RuBSCO enzyme and RuBP Carboxylase [40]. The increase in the total soluble carbohydrates in the leaves when sprayed with the antioxidant’s AsA and calcium is due to the role of these two compounds in increasing the efficiency of the photosynthesis process and the leaf area exposed to light, which led to an increase in carbohydrates of leaves.
The proline accumulation under heat stress conditions gives the plant the energy needed for growth and endurance to stress [41]. The increased proline in plants exposed to stress may adapt to protect the plant in such conditions. Proline acts in many physiological roles and functions under environmental stress conditions, where it performs the process of osmoregulation inside the plant cell, in particular between the gap and the cytoplasm, as the accumulation of proline inside the cytoplasm works to regulate the state of homeostasis and facilitate the process of water absorption by the roots. In addition, proline protects the organelles from harmful effects such as oxidizing factors and contributes to the destruction of free radicals [42]. The treatment with the antioxidants AsA and calcium led to a modification of the proline content in the leaves. It can be attributed to their role in relieving heat stress on plants.
Phenolic compounds are plants’ most important secondary metabolites that act as natural antioxidants, free radical scavengers, inhibitors of free radical production, and catalysts for antioxidant synthesis [43]. The increase in total phenols by treatment with AsA and calcium is due to the critical role of this antioxidant in reducing respiration. Increased phenols indicate plant adaptation to extreme environmental conditions [43].
The decrease in plant growth at 49/31°C was associated with the emergence of stress damages reflected in decreased membrane stability, indicating membrane damage. It is probably due to a loss of ability to rapidly and completely reorganize cell membranes. The previous studies have indicated membrane permeability as a valuable indicator of heat stress damage [44,45]. Our findings show that heat stress decreases membrane stability when electrolyte leakage is used as the index of permeability of membranes. In plants treated with AsA and calcium, a significant increase in membrane stability, especially for those grown at 49/31°C. A higher temperature tolerance was observed in plants grown than those not treated with AsA and calcium. The higher membrane stability index resulting from AsA and calcium is related to the antioxidant responses that protect the plant from oxidative damage and the higher ionic content and induced activities of antioxidant enzymes.
The increase in calcium in the leaves [Figure 3c] when treated with calcium or AsA due to the action of these substances is similar to the action of antioxidants, thus reducing the oxidative process, increasing the activity of the vital processes of the plant [46]. In addition to being a stimulant for the representation of proteins and enzymes that accompany carbohydrate representation, photosynthesis, and ionic balance, as well as being an osmotic regulator that participates in the processes of opening and closing stomata, etc. [47].
Heat stress is the cause of an increase in ABA, and growth inhibitors, such as ABA, known as one of the components of the transduction signal, lead to gene induction and the formation of proteins necessary to protect plants under stress conditions [48]. Examination of protein patterns in control and treatments showed that the relative density in Indian almonds was more significantly related to the AsA 100 mg/L + Cacl2 500 mg/L, AsA 50 mg/L, and AsA 100 mg/L treatments than the other treatments [Figure 4]. Plant development and variation to ecological circumstances are affected by protein metabolism [49]. Different examinations have demonstrated that the development of new proteins connects with changes in natural plant circumstances, for example, heat stress, prompting an expansion or diminishing in polypeptides. The results demonstrate that heat stress prompted the development of new proteins and the disappearance of others, accompanied by a distinctive appearance of stress proteins. The formation of new proteins affected by high temperature demonstrates that these proteins were shaped ahead of schedule by programming cell passing as we noticed shrinkage and stiffness in plants [Figure 6a]. All ecological burdens invigorate the creation of a gathering of proteins called HSPs, or stress-incited proteins. The acceptance of the record of these proteins is a typical peculiarity in every living being. These proteins are assembled in plants into five classes as per their approximate molecular weight: (1) Hsp100, (2) Hsp90, (3) Hsp70, (4) Hsp60, and (5) small HSPs. This broadening of these proteins reflects variation in enduring heat stress. The heat shock protein gene guideline is controlled by regulated proteins called heat stress transcription factors. These proteins likewise coordinate in all periods of cyclic reactions to high temperature (operation, maintenance, and recovery [Figure 6b] [50].
 | Figure 6: (a) Exposure of Indian almond seedlings to thermal stress, (b) the recovery of Indian almond seedlings from thermal stress. [Click here to view] |
The plant’s response to the high temperature in the summer, which included all the treatments, indicates its behavior in the semi-tropical regions with high temperatures in the summer, as it is a deciduous plant in the winter. When the plants are exposed to temperatures higher than 45°C, the plants are exposed to thermal stress. The hardening and shedding of leaves under the influence of high temperature may be attributed to the high concentration of growth inhibitors such as ABA, which leads to the production of ethylene and thus stiffness and defoliation [50]. Applying treatments led to a delay in the plant’s response to heat stress, due to the role of those compounds such as AsA and calcium in the plant’s tolerance of environmental stresses, including heat stress, by reducing oxidative stress, a state of metabolic compounds. One of the treatments that delay effect thermal stress is the treatment of AsA, which may be attributed to the physiological roles of AsA in protecting the plant from the harmful effects of high and low temperatures, reducing the damage of environmental stresses, reducing the harmful effect of ozone, the severity of freezing, drought stress, and its role in plant tolerance to drought from the method of closing the stomata during the day, which is essential to protect plants from high temperatures, and thus leads to a delay in plant aging [51].
5. CONCLUSIONS
Growing plants in an environment different from their original environment must improve their tolerance to new environmental conditions. The adaptation of the Indian almond plants to high temperatures during the summer in semi-tropical regions is possible by using antioxidants to reduce heat stress damage on the plant. Using AsA or calcium chloride alone or combined improved the plants’ tolerance to heat stress.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. FUNDING
There is no funding to report.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY
All data generated and analyzed are included in this research article.
11. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Henry RJ. Innovations in plant genetics adapting agriculture to climate change. Curr Opin Plant Biol 2020;56:168-73. [CrossRef]
2. Raza A, Razzaq A, Mehmood SS, Zou X, Zhang X, Lv Y, et al. Impact of climate change on crops adaptation and strategies to tackle its outcome:A review. Plants (Basel) 2019;8:34. [CrossRef]
3. Guillot E, Hinsinger P, Dufour L, Roy J, Bertrand I. With or without trees:Resistance and resilience of soil microbial communities to drought and heat stress in a Mediterranean agroforestry system. Soil Biol Biochem 2019;129:122-35. [CrossRef]
4. Phulwaria M, Ram K, Harish, Gupta AK, Shekhawat NS. Micropropagation of mature Terminalia catappa (Indian Almond), a medicinally important forest tree. J For Res 2012;17:202-7. [CrossRef]
5. Rojas-Sandoval J, Meléndez-Ackerman E. Population dynamics of a threatened cactus species:General assessment and effects of matrix dimensionality. Popul Ecol 2013;55:479-91. [CrossRef]
6. Akubude VC, Maduako JN, Egwuonwu CC, Olaniyan AM, Ajala EO, Ozumba CI, et al. Effect of processing parameters on the expression efficiency of almond oil in a mechanical expression rig. Agric Eng Int CIGR J 2018;20:109-17.
7. Shareef HJ, Alhamd AS, Naqvi SA, Eissa MA. Adapting date palm offshoots to long-term irrigation using groundwater in sandy soil. Folia Oecol 2020;48:55-62. [CrossRef]
8. Chen K, Zhang M, Zhu H, Huang M, Zhu Q, Tang D, et al. Ascorbic acid alleviates damage from heat stress in the photosystem II of tall fescue in both the photochemical and thermal phases. Front Plant Sci 2017;8:1373. [CrossRef]
9. Shareef HJ. Role of antioxidants in salt stress tolerant of date palm offshoots (Phoenix Dactylifera L) female and male cultivars. Int J Curr Agric Res 2015;9:8.
10. Subedi RP, Vartak RR, Kale PG. Management of heat stress in Drosophila melanogasterwith Abhrak bhasma and ascorbic acid as antioxidant supplements. J Appl Biol Biotechnol 2018;6:20-6.
11. Jasim AM, Abbas MF, Shareef HJ. Calcium application mitigates salt stress in Date Palm (Phoenix dactylifera L.) offshoots cultivars of berhi and sayer. Acta Agric Slov 2016;107:103. [CrossRef]
12. Naeem M, Khan MN, Khan MM, Moinuddin AS. Adverse Effects of Abiotic Stresses on Medicinal and Aromatic Plants and their Alleviation by Calcium. In:Tuteja N, Gill SS, editors. Plant Acclimation to Environmental Stress. New York:Springer Science Business Media;2013. 1-493. [CrossRef]
13. Bahamonde HA, Gil L, Fernández V. Surface properties and permeability to calcium chloride of Fagus sylvatica and Quercus petraea leaves of different canopy heights. Front Plant Sci 2018;9:494. [CrossRef]
14. Myrene RD, Devaraj VR. Role of calcium in increasing tolerance of Hyacinth bean to salinity. J Appl Biol Biotechnol 2013;1:11-20.
15. Kaneria MJ, Rakholiya KD, Marsonia LR, Dave RA, Golakiya BA. Nontargeted metabolomics approach to determine metabolites profile and antioxidant study of tropical almond (Terminalia catappa L.) fruit peels using GC-QTOF-MS and LC-QTOF-MS. J Pharm Biomed Anal 2018;160:415-27. [CrossRef]
16. Easlon HM, Bloom AJ. Easy leaf area:Automated digital image analysis for rapid and accurate measurement of leaf area. Appl Plant Sci 2014;2:1400033. [CrossRef]
17. Lichtenthaler HK, Wellburn AR. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 1983;11:591-2. [CrossRef]
18. Yemm EW, Willis AJ. The estimation of carbohydrates in plant extracts by anthrone. Biochem J 1954;57:508-14. [CrossRef]
19. Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water stress studies. Plant Soil 1973;39:205-7. [CrossRef]
20. Waterman PG, Mole S. Analysis of Phenolic Plant Metabolites. Hoboken:Blackwell Scientific Publications;1994. 235.
21. Luwe MW, Takahama UH, Heber U. Role of ascorbate in detoxifying ozone in the apoplast of spinach (Spinacia oleracea L.) leaves. Plant Physiol 1993;101:969-76. [CrossRef]
22. Tang Y, Wang L, Ma C, Liu J, Liu B, Li H. The use of HPLC in determination of endogenous hormones in anthers of bitter melon. J Life Sci 2011;5:139-42.
23. Shanahan JF, Edwards IB, Quick JS, Fenwick JR. Membrane thermostability and heat tolerance of spring wheat. Crop Sci 1990;30:247. [CrossRef]
24. Cresser MS, Parsons JW. Sulphuric-perchloric acid digestion of plant material for the determination of nitrogen, phosphorus, potassium, calcium, and magnesium. Anal Chim Acta 1979;109:431-6. [CrossRef]
25. Waling I, Houba WV, Lee JJ. Soil and plant analysis, a series of syllabi. In:Plant Analysis Procedures. Vol. 7. Netherlands:Wageningen University;1989. 712-7.
26. Laemmli U. Cleavage of structural proteins during the assembly of the head bacteriophage T4. Nature 1970;227:680-5. [CrossRef]
27. Fragkostefanakis S, Röth S, Schleiff E, Scharf KD. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ 2015;38:1881-95. [CrossRef]
28. Nievola CC, Carvalho CP, Carvalho V, Rodrigues E. Rapid responses of plants to temperature changes. Temperature (Austin) 2017;4:371-405. [CrossRef]
29. Hatfield JL, Prueger JH. Temperature extremes:Effect on plant growth and development. Weather Clim Extrem 2015;10:4-10. [CrossRef]
30. Jamloki A, Bhattacharyya M, Nautiyal MC, Patni B. Elucidating the relevance of high temperature and elevated CO2 in plant secondary metabolites (PSMs) production. Heliyon 2021;7:e07709. [CrossRef]
31. Ogbonna JC, Nweze, NO, Ogbonna CN. Effects of light on cell growth, chlorophyll, and carotenoid contents of Chlorella sorokiniana and Ankistrodesmus falcatus in poultry dropping medium. J Appl Biol Biotechnol 2021;9:157-63.
32. Akram NA, Shafiq F, Ashraf M. Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front Plant Sci 2017;8:613. [CrossRef]
33. Ismael BF, Abd AK, Jabbar FJ. Study the effect of antioxidants on the traits of the fruits of two cultivars of jujube (Ziziphus mauritiana Lamk.) Al-Tufahi and Alarmouti cultivars. Basrah J Agric Sci 2022;35:1-20. [CrossRef]
34. Alabdulla SA, Alfreeh L, Al-shumary A. The Impact of Foliar Spray with Ascorbic acid on some Growth Parameters and Grain Yield for Two Genotypes of Maize Zea mays L. In:2nd Al-Noor International Conference for Science and Technology 2NICST2020. Baku, Azerbaijan;2020. 198-202.
35. Al-Mayahi AM. Influence of salicylic acid (SA) and ascorbic acid (ASA) on in vitro propagation and salt tolerance of date palm (Phoenix dactylifera L.) cv. “Nersy.“Aust J Crop Sci 2016;10:969-76. [CrossRef]
36. Faisal HA, Jerry AN, Abbas MF. Effect of salicylic and ascorbic acids and method of application on flowering and green yield of broad bean (Vicia faba L.) plants. Basrah J Agric Sci 2014;27:34-43. [CrossRef]
37. Ullah HA, Javed F, Wahid A, Sadia B. Alleviating effect of exogenous application of ascorbic acid on growth and mineral nutrients in cadmium stressed barley (Hordeum vulgare) seedlings. Int J Agric Biol 2016;18:73-9. [CrossRef]
38. Bhattacharya A, Palan B, Char B. An insight into wheat haploid production using wheat x maize wide hybridization. J Appl Biol Biotechnol 2015;3:44-7.
39. White PJ, Broadley MR. Calcium in plants. Ann Bot 2003;92:487-511. [CrossRef]
40. Kahrizi S, Sedghi M, Sofalian O. Effect of salt stress on proline and activity of antioxidant enzymes in ten durum wheat cultivars. Ann Biol Res 2012;3:3870-4.
41. Liang X, Zhang L, Natarajan SK, Becker DF. Proline mechanisms of stress survival. Antioxid Redox Signal 2013;19:998-1011. [CrossRef]
42. Kishor PB, Kumari P, Sunita MS, Sreenivasulu N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front Plant Sci 2015;6:544. [CrossRef]
43. Cosme P, Rodríguez AB, Espino J, Garrido M. Plant phenolics:Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants (Basel) 2020;9:1263. [CrossRef]
44. El Basyoni I, Saadalla M, Baenziger S, Bockelman H, Morsy S. Cell membrane stability and association mapping for drought and heat tolerance in a worldwide wheat collection. Sustain 2017;9:1606. [CrossRef]
45. Jia K, Da Costa M, Ebdon JS. Comparative effects of hydro-, hormonal-, osmotic-, and redox-priming on seed germination of creeping bentgrass under optimal and suboptimal temperatures. HortScience 2020;55:1453-62. [CrossRef]
46. Alhamrani KS, Abdullah AA, Taain DA. Effect of planting dates, soil mulching and foliar spraying calcium on the quantitative yield characteristics of okra plant grown in plastic greenhouse. Basrah J Agric Sci 2019;32:171-82. [CrossRef]
47. Da Silva DL, de Mello Prado R, Tenesaca LF, da Silva JL, Mattiuz BH. Silicon attenuates calcium deficiency by increasing ascorbic acid content, growth and quality of cabbage leaves. Sci Rep 2021;11:1770. [CrossRef]
48. Vishwakarma K, Upadhyay N, Kumar N, Yadav G, Singh J, Mishra RK, et al. Abscisic acid signaling and abiotic stress tolerance in plants:A review on current knowledge and future prospects. Front Plant Sci 2017;8:161. [CrossRef]
49. Schlüter U, Colmsee C, Scholz U, Bräutigam A, Weber APM, Zellerhoff N, et al. Adaptation of maize source leaf metabolism to stress related disturbances in carbon, nitrogen and phosphorus balance. BMC Genomics 2013;14:23-31. [CrossRef]
50. Islam MR, Feng B, Chen T, Tao L, Fu G. Role of abscisic acid in thermal acclimation of plants. J Plant Biol 2018;61:255-64. [CrossRef]
51. Khan TA, Mazid M, Mohammad F. A review of ascorbic acid potentialities against oxidative stress induced in plants. J Agrobiol 2012;28:97-111. [CrossRef]